
基于优化MaxEnt 模型的南方三棘果在中国的潜在适生区预测
2023-02-22 02:48曾权朱雪珍周利娟
华南农业大学学报 2023年2期
关键词:模型
曾权 朱雪珍 周利娟


摘要: 【目的】分析预测南方三棘果Emex australis 在我国的潜在适生区以及影响其分布的主要环境变量,为防止南方三棘果入侵我国和保护我国农业生产、生态安全提供理论参考。【方法】运用刀切法(Jackknife) 计算各个环境变量對物种分布的影响。使用ENMeval 软件包对MaxEnt 生态位模型进行优化处理,将南方三棘果的分布数据和不同气候情景下的气候数据输入优化后的MaxEnt 模型,对我国潜在分布区进行预测。【结果】最冷季度平均气温(Bio11) 对南方三棘果分布影响最大,贡献率为27.7%。环境因子响应曲线表明,最冷季度平均气温为9.35~12.76 ℃ 时,南方三棘果的存在概率大于0.5;MaxEnt 结果表明,南方三棘果在我国的适生区主要集中于云南、广东、广西和福建。【结论】对我国南方三棘果适生区应当建立常态化监测方案,在适生区最冷季度平均气温适宜其生存的年份要加大监测力度,防止其在我国定殖、扩散。
关键词: 南方三棘果;MaxEnt 模型;潜在适生区;杂草入侵
中图分类号: S451;S412文献标志码: A 文章编号: 1001-411X(2023)02-0254-09
随着经济全球化的加深,我国进口货物和进口国家数量不断增长,同时,许多有害杂草伴随着国际流通进入我国[1]。研究表明,进口额每增长1 个百分点,植物疫情在海关截获量约增长2 个百分点[2]。近年来有多起南方三棘果Emex australis 在海关截获的报告。例如,2012—2015 年广东黄埔口岸在主要进境粮食中截获的有害生物中,南方三棘果截获高达90 批次排第7 位。此外在广东的肇庆和汕头口岸以及江苏口岸也都有截获报道[3-4]。
南方三棘果是蓼科亦模属的冬型一年生杂草,喜生长在湿润和半湿润热带、亚热带和温带气候区,被我国列入进口植物检疫对象。目前在我国内地尚未发现,仅在台湾有分布记录。南方三棘果单株植株可以产生1 000 粒以上的果实,种子在土壤中休眠多年,仍有繁殖能力,且具有繁殖速度快、传播迅速、危害性大和防治难度高的特点,一旦大面积入侵我国,难以根治,将会给农业生产造成巨大破坏[5-7]。
随着信息化技术的发展,目前用于预测物种潜在分布区的模型有CLIMEX、GARP、DIVA-GIS、B I O C L IM 和M a x E n t 等。相比于其他模型,MaxEnt 生态位模型在分布数据较少的情况下,预测精确度和准确度高[8-9]。其通过将研究区域的环境因子转换为栅格数据,形成多个栅格单元,把物种已知分布区域的栅格单位作为样点,然后以分布点区域的环境变量数据对所有栅格单元进行数理统计,得到当前和未来一段时间内研究区域的每一个栅格单元分布的概率以及环境变量数值与分布概率之间的关系[10-12]。
由于各物种的研究和关注程度不同,物种在世界范围内详实的分布记录存在差异,因此物种的分布数据获得量存在差异[12]。南方三棘果属于数据获得量较少的物种,因此选择MaxEnt 作为预测南方三棘果在我国潜在适生区的模型。全球气温呈现升高趋势,会影响植物的潜在分布区域变化[13-14]。气候系统模式能够很好地反映气候变化的趋势,对未来一段时间的气候变化进行准确预测[15-17]。因此本文采用国家气候中心4 种气候情景下的未来气候数据对南方三棘果在我国的适生区进行分析预测。
本研究将MaxEnt 模型与ArcGIS 软件相结合,得到南方三棘果潜在分布区的可视化地图,以期能够为有关部门在潜在分布区的相关地区开展入侵风险评估、检验检疫和监测预警等提供理论参考,对防控南方三棘果传入、定殖和扩散,保障农业生产和生物多样性健康以及生态安全和经济贸易安全等具有重要意义[18]。
1 材料与方法
1.1 技术路线
技术路线包括查找和处理数据、模型优化与构建以及制作可视化地图3 部分,按照“物种分布点查找、处理→查找影响分布的环境因子以及SPSS分析→MaxEnt 模型参数优化→构建MaxEnt 模型→ArcGIS 制作可视化图”的研究方案进行,数据分析贯穿于整个研究过程。
1.2 相关软件和程序
MaxEnt 软件3.4.1 版、ArcGIS 软件10.4.1 版、SPSS Statistics 软件20.0 版、R 语言3.6.3 版、ENMeval 程序包以及Microsoft Excel 2016。
1.3 环境数据及处理
从世界气候数据网站(https://www.worldclim.org/) 下载2.1 版、分辨率为10 min(340 km2) 的1 9 7 0 — 2 0 0 0 年和国家气候中心气候系统模式(BCC-CSM2-MR)4 个气候情景模式(SSP1-2.6、SSP2-4.5、SSP 3-7.0 和SSP5-8.5)2021—2040 年世界范围内19 个生物气候变量数据(Bio1~Bio19) 以及高程海拔数据(Elev)。采用ArcGIS 软件从高程海拔数据中提取出坡度(Slo) 和坡向(Asp)[19-20]。一共得到温度、降水和海拔特征的环境变量数据22 个(表1)。
参考文献[21-22] 方法,用ArcGIS 将所有环境变量坐标系及基准面统一定义,并转出为ASCⅡ格式,以待MaxEnt 模型预测分布;使用R 语言将TIFF 格式的环境变量因子统一转换为ASC 格式,以待使用R 语言ENMeval 程序包对MaxEnt 模型进行优化。为了避免变量间的高度共线性导致的过拟合和减少运算量,对环境数据进行相关性分析。将物种分布点与环境变量输入MaxEnt 模型运算10 次,获得各个环境变量的贡献率,将环境数据和物种分布点数据在ArcGIS 处理为“*.txt”格式文件,将文件利用Excel 转化为“*.csv”格式文件,最后将分析数据导入SPSS 进行双变量相关性分析,作双尾检验。对相关系数(R)>0.8 的环境变量,选择贡献率较高的作为主要变量进行下一步分析,舍去贡献率较低项Bio5、Bio6、Bio7、Bio10、Bio12、Bio16 和Bio17。舍去贡献率为0 的环境变量Bio8。由于南方三棘果为冬型一年生草本植物,在最湿月和最暖季度是种子形态[ 2 3 - 2 4 ],因此舍去Bio13 和Bio18。最终得到12 个环境变量因子(表1),分别为Bio1、Bio2、Bio3、Bio4、Bio9、Bio11、Bio14、Bio15、Bio19、Elev、Slo 和Asp。
1.4 南方三棘果分布数据及处理
在全球生物多样性信息交换平台(https://www.gbif.org/) 等平台数据库检索物种分布相关记录,通过发表的文献、专著、文件、报告等查询相关分布信息及记录[25-27]。一共得到1 473 个分布数据点,去除重复、无效、缺失、错误分布数据后,得到1 205 个分布点数据。利用下载好的气候数据用ENMeval程序包对南方三棘果分布点进行过拟合分析,最终得到南方三棘果自然分布点647 个。
1.5 地图数据
从标准地图服务系统(http://bzdt.ch.mnr.gov.cn/) 下载GS(2022)4312 号标准地图。
1.6 模型优化
使用R 语言ENMeval 程序包将南方三棘果分布点分为2 部分,75% 随机样本数据用于训练集,2 5% 用于测试集,对M a x E n t 软件中调控倍频(Regularization multiplier,RM) 和特征组合(Featurecombination,FC) 参数进行优化,利用数据包设定RM 从0.1 到4,每次增加0.1,共40 个调控倍频,同时设定29 个特征组合,即H、L 、LH、LP、LPH、LPT、LPTH、LQ、LQH、LQP、LQPH、LQPT、LQPTH、LQT、LQTH、LT、P、PH、PT、PTH、Q、QH、QP、QPH、QPT、QT、QTH、T、TH,其中L 为线性(Linear),Q 为二次型(Quadratic),H 为片段化( H i n g e ) , P 为乘积型( P r o d u c t ) , T 为阈值型(Threshold),ENMeval 语言包将上述 1 160 种参数组合进行检验, 最终采用最小信息准则( T h eminimum information criterion AICc value) 模型评估受试者工作特征曲线(Receiver operating curve,ROC) 下的面积(Area under curve,AUC) 差异检验模型的拟合度与复杂度,当AICc 值最低(AICc=0) 时模型参数组合最佳[ 2 8 ],用于MaxEnt 模型建模。
1.7 构建 MaxEnt 预测模型
参考前人所述方法[29-33],将处理后的当前环境变量和物种分布点数据输入MaxEnt 模型,模型参数设置随机选择25% 的样本作为测试集,75% 的样本数据为训练集,同时勾选制作响应曲线功能以获得分布概率与气候因子的关系,勾选刀切法检验环境变量的重要程度,RM 与FC 取优化后的参数,其他参数为默认设置。以AUC 评价模型的准确性。AUC 越大,表明模型的可靠性越高,0.9 以上表示模型结果优秀可靠,为降低模型的随机性和模拟结果的不确定性,模型重复运行10 次,以运行的均值作为模型运算结果。
1.8 绘制物种分布图
将MaxEnt 模型生成的ASC 格式物种分布数据导入ArcGIS 软件,设置坐标系为地理坐标系GCS_WGS_1984,通过掩膜提取获得南方三棘果在我国的分布图,对预测结果利用自然间断点分级法进行重分类,得到南方三棘果的非适生区、低适生区、中适生区和高适生区。
2 结果与分析
2.1 MaxEnt 生态位模型优化结果
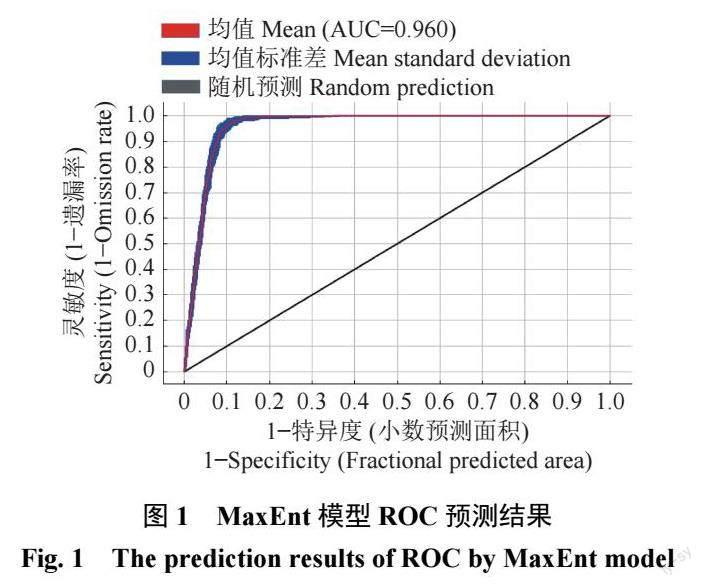
基于1 2 个环境变量因子和南方三棘果的647 个分布点数据,MaxEnt 软件初始默认RM=1,FC 为LQPH,AICc=142.939 251,经ENMeval 语言包优化后RM=1,FC 为LQPT,AICc=0,表明其为最优参数组合。在优化参数下进行模型预测,采用ROC 分析法进行模型准确度检验。经10 次重复训练得到的AUC > 0.9(图1),模拟结果优秀可靠。
2.2 影响分布环境变量因子贡献率及适宜范围
使用12 个环境因子建立模型,模型参数为ENMeval 语言包优化结果,RM=1,FC 为LQPT,采用刀切法评估各环境因子对南方三棘果地理分布的重要性。贡献率超过15% 的包括最冷季度平均气温(Bio11,27.7%)、等温性(Bio3,24.9%)、温度季节变化标准差(Bio4,18.2%),这3 个因子的贡献率合计达70.8%。贡献率低于15% 的包括年平均温度(Bio1,14.0%)、最干月降水量(Bio14,6.5%)、降水量变异系数(Bio15, 6.5%)、最冷季度降水量(Bio19,2.7%)、高程海拔(Elev,1.4%)、坡度(Slo,0.6%)、最干季度平均温度(Bio9,0.3%)、昼夜温差月均值(Bio2,0.2%)、坡向(Asp,0.2%)。
刀切法检验表明,仅使用单一环境变量时,正则化训练增益和测试增益都表明最重要的环境因子为最冷季度平均温度(Bio11)(图2)。
一般认为存在概率≥0.5 对应的环境因子范围是物种适宜分布的阈值。根据响应曲线确定影响南方三棘果分布环境因子的阈值:最冷季度平均温度9.35~12.76 ℃、等温性45.2%~53.72% 以及溫度季节变化标准差337.06~616.40 ℃(图3)。
2.3 南方三棘果在当前气候下的潜在适生分布区
将南方三棘果分布点和环境变量数据导入MaxEnt 模型进行模拟,得到南方三棘果在当前(2022 年) 气候下的潜在分布区(图4)。总适生面积约69.13 万km2,高适生区、中适生区、低适生区分别为11.64 万、13.37 万、44.02 万km2。南方三棘果的适生范围主要集中在我国云南、广西、广东、福建以及西藏局部,其中,高适生区和中适生区主要位于云南、海南以及西藏局部,低适生区主要位于广西、广东、台湾、福建和四川。
2.4 南方三棘果在未来气候模型下的潜在适生分布区
如表4 所示,SSP1-2.6 气候情景下,2040 年南方三棘果总适生区面积约63.54 万km2,较2022 年分布面积减少约5.48 万km2,其中,低适生区面积减少5.93 万km2,中适生区面积增加 6.57 万km2,高适生区面积减少6.12 万km2;SSP2-4.5 气候情景下,2040 年南方三棘果总适生区面积约65.10 万km2,较2022 年分布面积减少约3.92 万km2,其中,低适生区面积减少2.31 万km2,中适生区面积增加2.31 万km2,高适生区面积减少3.92 万km2;SSP3-7.0 气候情景下, 2040 年南方三棘果总适生区面积约6 3 . 3 4 万k m 2 ,较2 0 2 2 年分布面积减少约5.68 万km2,其中,低适生区面积减少9.74 万km2,中适生区面积增加6.51 万km2,高适生区面积减少2.45 万km2;SSP5-8.5 情景下,2040 年南方三棘果总适生区面积约62.90 万km2,较2022 年分布面积减少约6 . 1 2 万k m 2,其中,低适生区面积减少5.42 万km2,中适生区面积增加3.39 万km2,高适生区面积减少4.09 万km2。
如图5 所示,对不同气候情景下南方三棘果适生区的空间格局进行对比分析, 结果表明,2040 年,未来4 种情景模式下的适生区都呈现轻微缩小趋势,广东和广西的部分低适生区变为非适生区,云南的高适生区变为中适生区,海南的高、中、低适生区都呈减少趋势。从单个情景模式下看也有一些地区变得更加适应南方三棘果分布,SSP1-
2.6 氣候情景模式下西藏出现局部高适生区,四川的局部低适生区转变为中适生区;SSP2-4.5 气候情景模式下福建的一些非适生区变为低适生区;SSP3-7.0 气候情景模式下四川部分低适生区转变为中适生区,局部地区变为高适生区,西藏出现局部高适生区;SSP5-8.5 气候情景模式下,四川和西藏的一些非适生区变为低适生区,其中,西藏出现局部高适生区。这些变化区域为南方三棘果分布的敏感区。
3 结论与讨论
本模型的AUC 测试值 > 0.9,模型结果优秀可靠。最冷季度平均气温(Bio11)、等温性(Bio3) 和温度季节变化标准差(Bio4) 是影响南方三棘果地理分布的主要环境因子。正则化训练增益和测试增益以及贡献率都表明最冷季度平均气温(Bio11) 对南方三棘果分布影响最大,贡献率为27.7%。环境因子响应曲线表明,当最冷季度平均气温为9.35~12.76 ℃ 时,南方三棘果发生概率≥0.5。南方三棘果的适宜分布区总面积约为69.02 万km2,适生区域主要集中于云南、广东、广西和福建地区,其中高、中适生区主要集中于云南。
通过对2022—2040 年国家气候中心气候系统模式(BCC-CSM2-MR) 的4 种未来气候情景下的南方三棘果潜在分布区研究发现,总体上适生区呈现轻微减少趋势。四川、西藏、广东和广西属于南方三棘果分布的敏感区,易随气候变化而导致适生区等级以及适生面积发生变化。广东、广西、海南和云南的一些适生区在未来一段时间会出现转变为非适生区或降低适生等级的情况。福建部分地区在SSP2-4.5 情景模式下会更适宜南方三棘果生存,西藏和四川部分地区在SSP1-2.6、SSP3-7.0 和SSP5-8.5 情景下会更适宜南方三棘果生存。
其中,西藏的适生区尤为特殊,主要集中在藏南地区的错那县和墨脱县以及朗县最南部。这一部分地区由于地形原因导致气候复杂,高原气候和亚热带、热带季风气候同时存在,加上地处边疆,适生区西边与不丹接壤,南边与印度接壤,导致藏南外来物种入侵严重,值得相关部门进一步关注。中国科学院植物研究所的植物物种信息系统(http://www.iplant.cn/) 显示三叶鬼针草Bidens pilosa、胜红蓟Ageratum conyzoides 等原产于热带地区的外来杂草在墨脱县都有分布记录[34]。国家气候中心气候系统模式数据显示未来这一地区的温度季节变化标准差( B i o 4 ) 为3 9 0 ~ 5 5 0 ℃ , 等温性( B i o 3 ) 为39%~45%,最冷季度平均温度(Bio11) 为0.9~19 ℃,这种气候适宜南方三棘果分布,因此,对这里需要加大监测力度,阻止南方三棘果进入藏南地区,并防止由藏南地区入侵我国西南和华南地区。
南方三棘果是具有严重危害的检疫杂草,对农业生产会造成巨大破坏[35-36]。云南是我国葡萄十大产地之一,南方三棘果在我国的高、中适生区主要集中于云南,南方三棘果一旦定殖于葡萄园,可能会有部分种子在机械化收割时与葡萄一起收割,影响葡萄的品相和食品安全性[37]。藏南是西藏的主要农区,藏南地区生产的小麦是西藏的主要粮食作物之一,研究表明南方三棘果茎的水提取物和植株残留物会抑制小麦种子萌发和早期幼苗的生长,降低小麦产量[38-39]。油菜是广西、广东和福建的主要油料作物,在农田里南方三棘果与油料作物存在种间竞争关系[40]。
外来入侵杂草传入与人员和货物的流动密切相关,作为国与国之间管控人员和货物流通的重要节点,海关和动植物检验检疫局实施的植物检疫是防止外来杂草传入的最重要方式。在适生区域内以及邻近适生区域的海关和动植物检验检疫局应当加大对南方三棘果的检疫力度,防止南方三棘果的传入[41]。尤其值得注意的是,广东、广西和福建的海关每年都有大量的货物进口,加上这些地区存在大面积的南方三棘果适生区,海关和动植物检验检疫局的植物疫情数据也表明在广东和福建的海关有多起南方三棘果的截获记录,因此,这些地区是南方三棘果传入的高风险地区。
有害杂草入侵的定殖和扩散阶段在适生区范围内进行,其中,外部气候环境的变化是影响其扩散速度的重要因素,试验表明最冷季度平均温度是影响南方三棘果分布的最重要环境因子,在适生区气候适宜其生存的重要年份要加大南方三棘果的监测,有效防止其定殖、扩散,以免影响农作物的质量和产量[42]。生物入侵杂草的分布与气候环境、地形因素、土壤类型以及植被类型等多种因素相关,在使用MaxEnt 生态位模型预测入侵杂草潜在分布区时,目前尚未有统一的影响因子选择。朱煜建等[43] 在预测疣果匙荠Bunias orientalis 适生区时选择了气候环境、地形因素以及土壤类型的环境变量因子。郭燕青等[ 4 4 ] 在预测假臭草P r a x e l i sclematidea 时仅选择了气候因素有关因子。本研究选用与环境和地形有关的变量因子,缺少土壤类型、物种类型等与南方三棘果分布有关的其他变量,这可能对适生区的预测带来一定的偏差。伴随着我国与世界各国日益频繁的贸易往来,生物入侵难以避免。近些年假高粱Sorghum halepense和豚草Ambrosia artemisiifolia 等入侵生物对我国农业生产和生态安全造成了巨大破坏。采用MaxEnt生态位模型将信息学技术与生物的生态学特性结合,对各种入侵杂草的潜在分布区进行预测,对我国的口岸检疫和入侵杂草的防控具有指导意义。
参考文献:
陈凤新, 蒙彦良, 杜杰, 等. 中国外来生物入侵与社会经济因素多元回归分析[J]. 植物检疫, 2021, 35(4): 8-14.
李瑞法, 柳之光, 楚伟, 等. 我国进境植物疫情截获量的时序特征及预测[J]. 植物检疫, 2016, 30(4): 1-5.
王昕, 戴良英. 黄埔口岸进境粮食截获疫情分析[J]. 植物检疫, 2016, 30(6): 80-82.
伏建国, 高振兴, 李敏. 江苏口岸首次从进境加拿大油菜籽中截获检疫性杂草南方三棘果[J]. 植物检疫,2010, 24(4): 68.
MENDONCA L P, OLIVERIA E E, ANDERAZZA F, etal. Host potential and adaptive responses of Drosophilasuzukii (Diptera: Drosophilidae) to barbados cherries[J].Journal of Economic Entomology, 2019, 112(6): 3002-3006.
王瑞, 万方浩. 入侵植物银毛龙葵在中国的适生区预测与早期监测预警[J]. 生态学杂志, 2016, 35(7): 1697-1703.
叶建峰, 唐丽霞, 吴先健, 等. 東莞口岸从进境燕麦粒中截获检疫性杂草南方三棘果[J]. 植物检疫, 2010, 24(2):42.
HERNANDEZ P A, GRAHAM C H, MASTER L L, etal. The effect of sample size and species characteristicson performance of different species distribution modelingmethods[J]. Ecography, 2010, 29(5): 773-785.
KHANGHAH S S, MOAMERI M, GHORBANI A, et al.Modeling potential habitats and predicting habitat connectivityfor Leucanthemum vulgare Lam. in northwesternrangelands of Iran[J]. Environmental Monitoring andAssessment, 2022, 194(2): 1-16.
王运生. 生态位模型在外来入侵物种风险评估中的应用研究[D]. 长沙: 湖南农业大学, 2007.
LUO C, XU W H, ZHOU Z X, et al. Habitat predictionfor forest musk deer (Moschus berezovskii) in Qinlingmountain range based on niche model[J]. Acta EcologicaSinica, 2011, 31(5): 1221-1229.
房锋, 张朝贤, 黄红娟, 等. 基于MaxEnt 的麦田恶性杂草节节麦的潜在分布区预测[J]. 草业学报, 2013, 22(2):62-70.
柳晓燕, 李俊生, 赵彩云, 等. 基于MAXENT 模型和ArcGIS 预测豚草在中国的潜在适生区[J]. 植物保护学报, 2016, 43(6): 1041-1048.
唐兴港, 袁颖丹, 张金池. 气候变化对杉木适生区和生态位的影响[J]. 植物研究, 2022, 42(1): 151-160.
李安, 李良涛, 高萌萌, 等. 基于MaxEnt 模型和气候变化情景入侵种黄顶菊在中国的分布区预测[J]. 农学学报, 2020, 10(1): 60-67.
姜大膀, 富元海. 2 ℃ 全球变暖背景下中国未来气候变化预估[J]. 大气科学, 2012, 36(2): 234-246.
JAMAL Z A, ABOU-SHAARA H F, QAMER S, et al.Future expansion of small hive beetles, Aethina tumida,towards North Africa and South Europe based on temperaturefactors using maximum entropy algorithm[J].Journal of King Saud University-Science, 2021, 33(1): 1-17.
KONG F, TANG L, HE H, et al. Assessing the impact ofclimate change on the distribution of Osmanthus fragransusing MaxEnt[J]. Environmental Science and PollutionResearch, 2021, 28(26): 34655-34663.
辛晓歌, 吴统文, 张洁, 等. BCC 模式及其开展的CMIP6 试验介绍[J]. 气候变化研究进展, 2019, 15(5):533-539.
王翀, 林慧龙, 何兰, 等. 紫茎泽兰潜在分布对气候变化响应的研究[J]. 草业学报, 2014, 23(4): 20-30.
SONG R, MA Y, HU Z, et al. MaxEnt modeling of Dermacentormarginatus (Acari: Ixodidae) distribution inXinjiang, China[J]. Journal of Medical Entomology,2020, 57(5): 1659-1667.
ZHANG M G, SLIK J, MA K P. Using species distributionmodeling to delineate the botanical richness patternsand phytogeographical regions of China[J]. Scientific Reports,2016, 6(1): 22400. doi: 10.1038/srep22400.
GRAHAM M H. Confronting multicollinearity in ecologicalmultiple regression[J]. Ecology, 2003, 84(11):2809-2815.
张路. 基于MAXENT 模型预测齿裂大戟在中国的潜在分布区[J]. 生物安全学报, 2015, 24(3): 194-200.
任子春. 基于MaxEnt 模型在全球变暖条件下菟丝子属全寄生植物及其寄主的潜在分布预测[D]. 临汾: 山西师范大学, 2020.
郭水良, 高平磊, 娄玉霞. 应用MaxEnt 模型预测检疫性杂草毒莴苣在我国的潜分布范围[J]. 上海交通大学学报(农业科学版), 2011, 29(5): 15-19.
WILLIAMS P, YA T, JIAN Y, et al. The bumblebees ofSichuan (Hymenoptera: Apidae, Bombini)[J]. Systematicsand Biodiversity, 2009, 7(2): 101-187.
叶兴状, 张明珠, 赖文峰, 等. 基于MaxEnt 优化模型的闽楠潜在适宜分布预测[J]. 生态学报, 2021, 41(20):8135-8144.
ELITH J, GRAHAM C H, ANDERSON R P, et al. Novelmethods improve prediction of species distributionsfrom occurrence data[J]. Ecography, 2010, 29(2): 129-151.
张海娟, 陈勇, 黄烈健, 等. 基于生态位模型的薇甘菊在中国适生区的预测[J]. 农业工程学报, 2011, 27(S1):413-418.
朱耿平, 范靖宇, 王梦琳, 等. ROC 曲线形状在生态位模型评价中的重要性——以美国白蛾为例[J]. 生物安全学报, 2017, 26(3): 184-190.
PEARCE J L, BOYCE M S. Modelling distribution andabundance with presence-only data[J]. Journal of AppliedEcology, 2006, 43(3): 405-412.
KOZAK K H, GRAHAM C H, WIENS J J. IntegratingGIS-based environmental data into evolutionarybiology[J]. Trends in Ecology & Evolution, 2008, 23(3):141-148.
陈星, 赵韦, 包海龙. 西藏山南地区采集纪行——错那县[J]. 生命世界, 2018(3): 94-95.
张玉环, 郝建华, 吴海荣, 等. 外来入侵植物胜红蓟的胚胎学观察及繁殖系统研究[J]. 植物科学学报, 2020,38(2): 162-172.
王桔红, 史生晶, 陈文, 等. 鬼针草与含羞草化感作用及其入侵性的研究[J]. 草業学报, 2020, 29(4): 81-91.
POHLNER D. Emex australis and dried vine fruit productionin Sunraysia[J]. Plant Protection Quarterly, 1996,11(4): 150-153.
JAVAID M M, TANVEER A, ALI H H, et al. Wheatyield loss in a two species competition with Emex australisand Emex spinosa[J]. Planta Daninha, 2016, 34: 35-46.
ABBAS R N, TANVEER A, ALI A, et al. Effects ofEmex australis Steinh on germination and early seedlinggrowth of wheat (Triticum aestivum L. )[J]. AllelopathyJournal, 2010, 25(2): 513-520.
SCOTT J K, BOWRAN D G. Workshop to identify researchpriorities for Emex species[J]. Plant ProtectionQuarterly, 1996, 11(4): 175-176.
PUCHA?KA R, DYDERSKI M K, V?TKOV? M, et al.Black locust (Robinia pseudoacacia L. ) range contractionand expansion in Europe under changing climate[J].Global Change Biology, 2021, 27(8): 1587-1600.
梁忆冰. 植物检疫对外来有害生物入侵的防御作用[J].植物保护, 2002(2): 45-47.
朱煜建, 周伟光, 张昊, 等. 刺蒺藜草在我国的适生区预测[J]. 植物检疫, 2022, 36(1): 71-76.
郭燕青, 史梦竹, 李建宇, 等. 基于MaxEnt 模型的假臭草潜在分布区预测[J]. 热带亚热带植物学报, 2019,27(3): 250-260.
【责任编辑 霍 欢】
猜你喜欢
网络安全与数据管理(2022年1期)2022-08-29
导航定位学报(2022年4期)2022-08-15
中学生数理化·中考版(2022年8期)2022-06-14
成都医学院学报(2021年2期)2021-07-19
新世纪智能(数学备考)(2020年9期)2021-01-04
中学生数理化·七年级数学人教版(2020年10期)2020-11-26
数学物理学报(2020年2期)2020-06-02
北京航空航天大学学报(2017年7期)2017-11-24
北京航空航天大学学报(2017年12期)2017-04-23
光学精密工程(2016年6期)2016-11-07
