
影响小麦面团强度的贮藏蛋白基因表达研究
2023-07-17 09:45晁岳恩王沙沙汪庆昌曹廷杰
麦类作物学报 2023年7期
晁岳恩,王沙沙,杨 剑,杨 攀,黄 超,汪庆昌,曹廷杰
(河南省农业科学院小麦研究所,河南郑州 450002)
小麦是我国主要农作物之一,是多种面食的原料。小麦面粉的独特性在于其蛋白与水相互作用能交联形成疏松多孔的面筋。小麦的加工品质与其蛋白含量密切相关,小麦粉含有8%~20%的蛋白质,其中面筋蛋白约占小麦总蛋白质含量的80%~85%,面筋蛋白的类型和含量影响面团的强度、粘弹性、延展性等指标,决定着面粉的加工适用性[1-5]。在实际面制品加工中发现,部分小麦品种蛋白含量较低,但面筋强度较高,说明面粉质量不仅受籽粒蛋白含量影响,蛋白成份也是重要的影响因素[6]。面筋蛋白的主要成份是麦谷蛋白和醇溶蛋白,麦谷蛋白按分子量大小可分为高分子量麦谷蛋白(HMW-GS)和低分子量麦谷蛋白(LMW-GS);HMW-GS通过分子间或分子内二硫键作用形成面筋骨架,决定着面团的强度和弹性[5-10]。HMW-GS仅占总面筋蛋白的10%,其亚基的组合方式被认为决定着70%的面粉质量特性[11-12];除遗传因素外,栽培措施对面粉质量也有影响[13]。
在育种过程中却发现,即使HMW-GS组合完全一致、蛋白含量相同的小麦品种的面粉质量存在较大差异,面团强度参数存在显著差异,暗示除HMW-GSs外,可能还存在其它影响面团强度性状的蛋白类型。本研究以具有一定亲缘关系且HMW-GS组合完全一致、蛋白含量基本相同、面粉质量有较大差异的两个品种为材料,在籽粒面筋蛋白积累速度较为稳定的灌浆中期[14-16],通过转录组测序分析两个品种面筋蛋白相关基因的表达差异,探讨其面粉质量差异的可能原因,为小麦品质育种提供参考。
1 材料与方法
1.1 试验材料
供试品种为郑麦369和郑麦158,前期研究发现,二者间HMW-GS组含完全相同,蛋白含量相同,面团强度差异显著。于2019—2020年度种植在河南省农业科学院试验基地,小区长6 m,宽2 m,行距0.2 m,相邻种植,无重复,常规管理。郑麦369亲本组合为郑麦366×良星99,郑麦158亲本组合为(Bigeaz-250/96)×周麦16)×郑麦366。转录组测序分析样品分别取自花后14、21、28 d的穗中部两侧籽粒,液氮速冻后带回试验室保存于-80 ℃冰箱。收取成熟籽粒,磨粉并测定面粉质量参数。
1.2 面粉品质参数检测
制粉:参照AACC 26-20方法,用BUHLER实验磨磨粉。粗蛋白含量:用丹麦FOSS公司凯氏定氮仪Kjeltec测定。面团流变学参数:用德国Brabender公司的810104型粉质仪(Farinograph),按GB/T14614-93测定吸水率、面团形成时间、稳定时间、弱化度。面筋参数:用瑞典Perten公司的2200型面筋仪(Glutomatic),按GB/T14608-93测定小麦粉湿面筋含量及湿面筋指数。
1.3 面粉蛋白提取
醇溶蛋白和麦谷蛋白提取按照文献优化的方法[17],其中,SDS-PAGE分离胶浓度12%,浓缩胶浓度4%,考马斯亮蓝(R-250)染色。
1.4 转录组测序及生物信息学分析
RNA提取、转录组测序及生物学信息分析由杭州联川生物技术有限公司(中国,杭州)完成,所有技术操作、基因组比对、转录本组装、FPKM定量及GO和KEGG富集分析工作均由该公司完成。每个样品三次生物学重复。使用StringTie软件(https://ccb.jhu.edu/software/hisat2)对基因或转录本进行组装并用FPKM[每百万测序片段中来自某一基因每千碱基长度的数目:FPKM = total_exon_fragments / mapped_reads(millions) 的exon_length(kB)]定量,使用R包edgeR(https://bioconductor.org/packages/release/bioc/html/edgeR.html)对样本之间的差异基因进行分析,差异倍数>2倍或<0.5倍,且P<0.05定义为差异表达基因。
1.5 面粉硫及巯基含量测定
硫含量检测:浓硝酸消化后用液相离子色谱测定硫元素含量(IC-2001; TOSOH, Japan),阴离子标准液使用Wako Pure Chemicals(Japan),详细操作采用Maruyama优化的方法[18]。总巯基和自由巯基含量测定及二硫键含量计算参考Wang等[19]的方法。
1.6 蛋白质量评价
从NCBI数据库下载不同类型的HMW-GSs氨基酸序列信息,利用巯基和二硫键含量预测在线程序(SCRATCH Protein Predictor:http://scratch.proteomics.ics.uci.edu),对部分差异表达基因编码蛋白和HMW-GSs的自由巯基、二硫键含量进行预测,并利用蛋白质量评价模型进行评分比较[20]。
1.7 数据处理
采用 Excel 2007进行数据分析和显著性检验。
2 结果与分析
2.1 面粉质量参数分析
从表1可知,两个品种的粗蛋白含量无显著差异,郑麦158的湿面筋含量和吸水率显著低于郑麦369,但面筋指数、形成时间和稳定时间显著高于后者,郑麦158稳定时间约为郑麦369的两倍,推测郑麦158含有较多对面团强度贡献较大的面筋蛋白。
2.2 面筋蛋白亚基分析
从图1可以看出,两个品种的HMW-GS组合完全一致(1、5+10、7+8),说明HMW-GS组合可能不是两个品种之间面粉质量差异的主要原因;LMW-GSs和醇溶蛋白的电泳图谱稍有差异,推测这些蛋白的组成和含量差异可能是导致两个品种之间品质差异的原因。
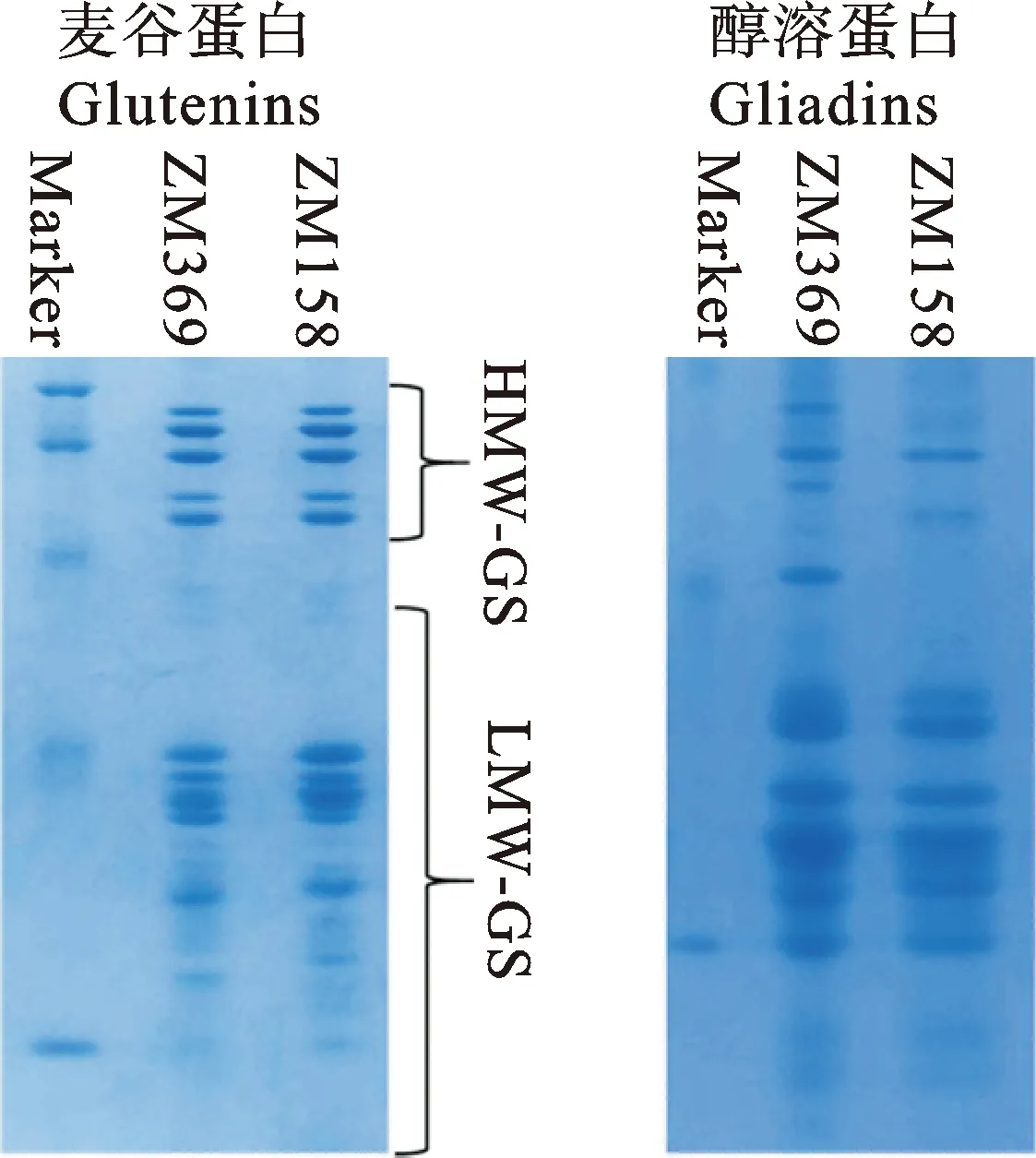
图1 两个品种的麦谷蛋白和醇溶蛋白电泳图
2.3 面筋蛋白的基因表达差异
根据测序结果,发现在小麦花后14、21、28 d时,两个品种间籽粒中HMW-GS基因表达均无显著性差异,这与其蛋白检测结果相同,说明HMW-GSs不是两个品种间面团强度性状差异的主要原因。由表2可知,与郑麦369比较,郑麦158在3个时间点共有24个(46次)基因表达有显著差异,其中显著上调表达基因12个(23次),显著下调表达基因12个(23次)。上调表达基因包括9个燕麦类似蛋白基因(18次)、2个γ-醇溶蛋白基因(4次)和1个α-醇溶蛋白基因;下调表达基因包括11个α-或α/β-醇溶蛋白基因(21次)和1个燕麦类似蛋白基因(2次)。其中,燕麦类似蛋白基因占郑麦158显著上调表达基因总量的75%,暗示燕麦类似蛋白可能对面团强度性状有重要作用。

表1 两品种的面粉品质特性
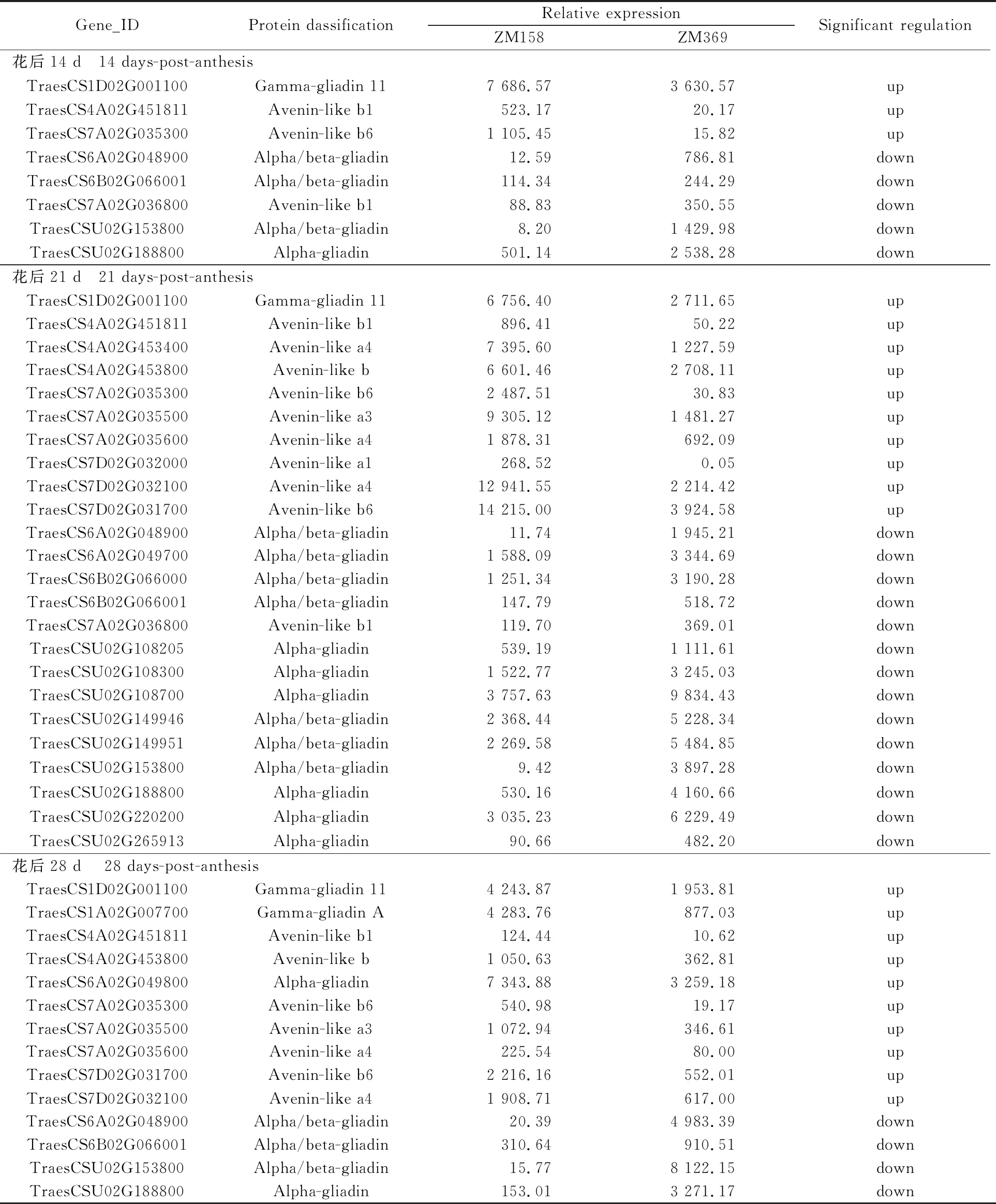
表2 不同发育时期籽粒中表达显著差异的贮藏蛋白基因
郑麦158显著上调表达基因的染色体组定位分析显示,上调表达的基因全部位于A、D染色体组,其中A染色体组基因出现15次,占总数的65%。另外,从燕麦类似蛋白的染色体定位看,这些基因分别定位在7A、7D和4A染色体,7B染色体未发现差异表达的燕麦类似蛋白基因。所以,从本研究两个材料来说,A组染色体编码的面筋蛋白与面粉强度性状的相关性最高,D组次之。
2.4 面粉硫及巯基集团含量分析
为分析两个品种的巯基集团含量与面粉品质的相关性,对面粉中的硫和巯基(自由巯基、总巯基)及分子内二硫键含量进行了分析,结果(表3)发现,二者间总巯基含量无显著差异,郑麦158的总硫含量明显较低,其自由巯基含量显著高于郑麦369。面粉中面筋蛋白占总蛋白含量的80%以上,可以用面粉中的硫及巯基含量衡量其在面筋蛋白中的含量。暗示在含硫氨基酸的组成上,郑麦158面筋蛋白中的半胱氨酸残基含量比例较高,且分子内二硫键比例较低,而郑麦369面筋蛋白中的甲硫氨酸残基含量比例较高。

表3 两个品种面粉中的巯基含量
2.5 差异表达蛋白的质量评价
根据在线软件(SCRATCH Protein Predictor)的分析原理,蛋白分子内二硫键的有无和自由巯基数量分属两个独立的预测,即:当一个蛋白被预测为无分子内二硫键时,也会预测潜在的二硫键含量。因此,在对蛋白的面粉质量贡献进行评价时,首先考虑二硫键的有,然后再考虑二硫键的数量。当一个蛋白预测为无分子内二硫键时,其半胱氨酸数量全部视为自由巯基数量;有分子内二硫键时,总半胱氨酸数量减去形成二硫键的半胱氨酸数量后剩余的半胱氨酸数量即为自由巯基数量。具体评分方法按照分值=0.9x+0.3y计算,其中x为某个蛋白质的自由巯基数量,y为这个蛋白质的分子内二硫键数量[20]。
从数据库下载的部分HMW-GSs和差异表达基因编码蛋白的评估分值见表4和表5,按照HMW-GS对面团强度贡献评分不低于3.6作为优质蛋白评价标准,11个上调的差异表达基因编码蛋白中,有9个达到优质蛋白标准,其中γ-醇溶蛋白(TraesCS1D02G001100)在3个时间点都显著上调表达,且分值超过了全部HMW-GS的评分;仅有有两个基因(TraesCS4A02G451811:Avenin-like b1;TraesCS1A02G007700:Gamma-gliadin A)没有达到优质亚基标准,但其评分(3.0分)也与中等类型的HMW-GS相同。在分析的3个下调基因中(在3个时间点出现8次),也有1个编码蛋白达到了优质HMW-GS的分值,表明可能也有对面团强度有贡献的蛋白在郑麦158中呈下调表达趋势。

表4 基于巯基预测结果的HMW-GS质量评价

表5 基于巯基预测结果的差异表达基因编码蛋白的质量评价
3 讨论
面团强度参数是衡量面粉品质的重要指标,与面包烘焙体积呈正相关[5]。HMW-GS及其组合方式是影响面团强度的主要遗传因素[5-12]。本研究结果却表明,燕麦类似蛋白和部分醇溶蛋白对面团强度性状也有较大影响。
燕麦类似蛋白为近年来发现的一类富含半胱氨酸的小麦面筋蛋白,被视为非典型面筋蛋白质成分(atypical gluten components),包括a、b两个大类,又可分为若干小类[21-22]。目前,燕麦类似蛋白与面粉品质的相关研究还不多,已有的研究与本研究结果相似,如Wang等[23]曾将中国春中的一个b类燕麦类似蛋白基因在郑麦9023中表达,转基因材料的面团强度明显提高,但在这个蛋白中额外引入1个半胱氨酸突变后,面团强度和弹性下降;二硫键预测结果表明,新增加的巯基与分子内的其他自由巯基形成了分子内二硫键,表明与总巯基含量相比,面筋蛋白的高自由巯基含量才是其影响面团强度的根本原因。Ma等[24-25]研究也发现,类燕麦b贮藏蛋白具有改善面团强度的潜力。关于a类燕麦类似蛋白与面粉质量的关系,目前尚未检索到相关研究文献,但从本研究结果看,高面团强度品种郑麦158的9个显著上调的燕麦类似蛋白基因中,包括5个a类燕麦类似蛋白基因,且评分也全部达到了优质蛋白水平,由此推测a类燕麦类似蛋白在面团强度性状中发挥着重要效应。
差异表达基因的染色体组定位分析显示,燕麦类似蛋白基因全部位于A、D染色体组,在B组染色体上没有发现燕麦类似蛋白基因。据推测,普通六倍体小麦的四倍体祖先中曾发生过4AL/7BS易位或近着丝粒倒位情况,可能导致了原来应该位于7BS的燕麦蛋白编码位点转移到了4AL上[26]。
醇溶蛋白是面筋蛋白的主要成分之一,通常认为,醇溶蛋白是以非共价键形式结合到面筋中,主要影响面团的粘性和延展性,对面团强度具有负向效应。本研究结果表明,高面团强度郑麦158的显著下调表达的基因中,醇溶蛋白基因数量占总数的78.6%,这与醇溶蛋白对于面团强度具有负向效应的观点一致[27-29]。但是,在郑麦158的显著上调表达基因中,两个γ-醇溶蛋白(TraesCS1D02G001100,TraesCS1A02G007700)基因也表现显著性上调,其中位于1D染色体的醇溶蛋白(TraesCS1D02G001100)基因在3个时期均表现显著性上调,生物信息学分析表明其含有8个自由巯基,面团强度的贡献评分为7.2分,超过了公认的优质HMW-GS(1Dy10和1Dx5),暗示其面团强度有较大贡献。有研究表明,某些醇溶蛋白也可以通过分子间二硫键结合到面筋中(尤其是奇数半胱氨酸残基含量的蛋白质至少存在一个自由巯基),进而影响到面团的强度[20,30-34]。
研究表明,面粉蛋白中的自由巯基和二硫键对面团结构及面团稳定性有重要影响,在揉面过程中,不同面粉蛋白质分子的自由巯基相互结合成二硫键,形成的面筋骨架决定着面团的结构和特性[35-39]。因此,小麦蛋白中的巯基含量是决定面团流变学特性及烘焙质量的关键因素[40-41]。本研究表明,高面团强度的郑麦158中,自由巯基含量、总巯基含量和总二硫键含量均高于面团强度较低的郑麦369。因此,进一步探索小麦籽粒蛋白中的高半胱氨酸和高自由巯基含量形成原因,对于阐释小麦面粉质量形成机制、完善优质小麦育种技术具有参考意义。
4 结论
燕麦类似蛋白在面团强度性状上有较大贡献,可能是决定面团强度性状的另一遗传因素,个别类型的γ-醇溶蛋白可能对面团强度性状也有贡献;探索小麦籽粒蛋白中高半胱氨酸和高自由巯基含量的形成原因,对于完善现有优质小麦育种技术具有一定的参考意义。
猜你喜欢
食品科学技术学报(2022年5期)2022-10-11
麦类作物学报(2022年1期)2022-03-01
质谱学报(2021年6期)2021-12-06
中国生物制品学杂志(2021年10期)2021-10-20
幼儿画刊(2020年9期)2020-11-04
粮食加工(2020年4期)2020-02-28
农民致富之友(2019年33期)2019-12-20
东方少年·阅读与作文(2018年3期)2018-05-14
美食(2017年4期)2017-05-04
