
荔枝铝激活苹果酸转运蛋白基因家族的鉴定及表达分析
2023-11-13 07:16钱义容全振炫郑雪文李伟才
广东农业科学 2023年9期
钱义容,王 弋,全振炫,郑雪文,李伟才,董 晨
(1.中国热带农业科学院南亚热带作物研究所/农业农村部热带果树生物学重点实验室,广东 湛江 524091;2.云南农业大学热带作物学院,云南 普洱 665099)
【研究意义】铝激活苹果酸转运蛋白(Alactivated malate transporter,ALMT)是植物中重要的一类膜蛋白,在植物有机酸的跨膜转运中起着重要作用,如参与苹果酸积累和转运、重金属胁迫、矿质营养、渗透势和pH值调节、果实品质形成、气孔运动、种子发育等过程[1-4]。【前人研究进展】在植物的生长过程中,酸性土壤中的铝离子(Al3+)会对植物根部造成毒害,ALMT 作为阴离子通道能够释放苹果酸盐有机阴离子并以此为螯合剂,与Al3+形成稳定无毒的复合物,从而降低其毒性[5]。最早Sasaki 等[6]在耐铝小麦(ET8)根尖中分离得到铝诱导表达的一个ALMT1基因(TaALMT1),是植物中克隆获得的第一个铝激活苹果酸转运基因。ALMT 蛋白大多定位于细胞质膜、液泡膜等细胞器上,N 端和C 端均位于膜的外侧,其N 端通常包含5~7 个跨膜结构域,参与Al3+结合的苹果酸运输,C 端有一个跨膜输水区来维持运输功能所必需[7-8]。基于ALMT 的重要性,拟南芥AtALMT1和小麦TaALMT1基因的鉴定引起人们对该家族的研究兴趣,目前,已知在拟南芥基因组中有14 个ALMTs 蛋白,其相关功能基因的表达也有一些报道[9]。AtALMT1参与AL3+的耐受,主要在拟南芥根中表达,其表达水平受过氧化氢、pH 值、植物激素的影响[10];AtALMT4主要在叶肉和保卫细胞中表达[11];AtALMT6主要在保卫细胞中表达,其表达水平受苹果酸、液泡pH 值和胞质钙离子的影响[11-12]。据报道,多种植物ALMT家族基因的生物学功能也相应被鉴定,如橡胶树(Heveabrasiliensis)中鉴定出17 个ALMTs基因[13];普通烟草(Nicotianatabacum)中鉴定出30 个ALMTs基因,该基因受AL3+的诱导影响,大多数在茎中表达较高而在根部表达相对较低[14];苹果(Malusdomestica)基因组中鉴定出25 个ALMTs 成员,其中MaALMT1影响果实酸度、主要在果实中表达[15];大豆中鉴定出34 个ALMTs基因,GmALMT5在低磷环境下显著表达[8];中国白梨中鉴定出27 个ALMTs基因,其中Pbr020270.1 作为候选基因可能在梨果实苹果酸的积累中发挥重要作用[16]。综上,ALMT基因家族成员在不同植物中均起着重要作用。荔枝(Litchichinensis)是无患子科荔枝属常绿乔木,产于亚热带地区,喜高温高湿气候,在历史上有“百果之王”等美称。但荔枝产业中仍存在一些问题,如大小年、缺乏特早熟优质荔枝、早熟荔枝偏酸等。【本研究切入点】ALMT 蛋白与植物生长发育和逆境响应有密切关系,荔枝果实酸度的研究目前集中在生理水平,分子水平的研究较少,ALMT 参与果实酸度调节,但在荔枝中ALMT基因家族的全基因组鉴定尚未见报道。【拟解决的关键问题】本研究基于全基因组水平对荔枝ALMT基因家族成员进行鉴定与分析,为ALMT基因家族在荔枝果实有机酸生物学功能研究方面提供初步依据。
1 材料与方法
1.1 试验材料
供试材料为‘妃子笑’荔枝,种植于中国热带农业科学院南亚热带作物研究所荔枝种植园(21°10′02″N、110°16′34″E,海拔21.32 m),试验基地地势平坦、水源充足、灌溉条件优良,土壤为红壤、肥力中等,土肥水管理一致。采集‘妃子笑’不同组织样品,取秋梢新抽发的嫩梢叶片,老熟梢的成熟老叶,盛花期的雄花、雌花和根(新生长的须根),果实成熟期的果皮、果柄,用液氮速冻后于-80 ℃保存,用于后续RNA 提取。
1.2 荔枝LcALMT 家族成员鉴定及其蛋白理化性质、亚细胞定位分析
荔枝品种‘妃子笑’(LitchichinensisSonn.cv.Feizixiao)的基因组来源于荔枝基因组数据库(http://www.sapindaceae.com/),META SEARCH工具搜索Al-activated malate transporter(铝激活苹果酸转运蛋白),搜索注释为Al-activated malate transporter 的基因,进一步通过NCBI Conserved Domains Search(https://www.ncbi.nlm.nih.gov/cdd)验证搜索到的蛋白序列是否含有保守ALMT 结构域,得到含有完整保守结构域的蛋白,并将其命名为LcALMT,最终从荔枝全基因组中确定有16个LcALMTs。运 用ExPASy(https:web.expasy.org/protparam/)预测家族成员对应蛋白质的长度(aa)、分子量(kD)、等电点(pI)、不稳定系数和亲水指数,运用在线网站(https:wolfpsort.hgc.jp/)进行亚细胞定位预测。
1.3 荔枝LcALMTs 系统进化关系与蛋白结构分析
为进一步了解LcALMTs 蛋白,利用公开发表模式植物拟南芥(14 个)和水稻(8 个)中的ALMTs 蛋白序列,拟南芥AtALMTs 蛋白序列从拟南芥官网TAIR(https://www.arabidopsis.org)下载获得,水稻OsALMTs 蛋白序列从网站Rice Genome Annotation Project(http://eice.plantbiology.msu.edu/)获得。16 个荔枝ALMTs 蛋白序列,使用MEGA 11 软件采用邻接法(Neighbor-Joining,NJ),保持其他参数不变,设置Bootstrap 值为1 000,构建系统进化树。
利用GORIV(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_gor4.html)预测蛋白质二级结构,然后利用在线工具SWISSMODEL(https://swissmodel.expasy.org/)预测蛋白质三级结构。
1.4 荔枝LcALMTs 的基因结构与保守基序分析
从荔枝基因数据库中获取ALMT家族成员的基因序列和对应的CDS序列,运用GSDS(http:gsds.cbi.pku.edu.cn/)在线软件进行荔枝ALMTs成员编码区、内含子和非编码区的结构分析。
使用MEME(http:meme-suite.org/tools/meme)在线软件设置基数为12,其余为默认参数下预测分析荔枝ALMT 家族成员编码蛋白的保守基序。
1.5 荔枝LcALMTs 启动子上顺式作用元件分析
利用在线网站Plantcare(http:bioinformatics.psb.ugent.be/webtools/plantcare/html/)对LcALMTs5'端上游2 000 bp 的序列进行顺式作用元件分析。
1.6 荔枝LcALMTs 的染色体定位
根据荔枝全基因组注释信息得到染色体基因位置信息,借助TBtools 软件实现可视化。
1.7 荔枝LcALMTs 的组织表达
从荔枝基因组数据库获取LcALMTs基因在不同组织中的表达量(http://www.sapindaceae.com/),通过TBtools 绘制热图。
1.8 部分LcALMTs 的实时荧光定量PCR
使用植物RNA 提取试剂盒(华越洋生物科技有限公司),提取妃子笑不同组织(根、嫩叶、老叶、雄花、雌花、果皮、果柄)RNA。取1 μL 检测RNA 质量和浓度,样品符合要求后利用M-MLV 逆转录酶(TaKaRa)反转录合成cDNA。利用在线软件Primer 3.0 设计荧光定量引物,以LcActin为内参(表1),通过Blast 分析引物的特异性,并交由广州艾基生物技术有限公司合成引物序列。

表1 RT-PCR 引物序列Table 1 Primer sequences for RT-PCR used in this study
使用SYBR Premix Ex TaqTM(TaKaRa)试剂盒制备反应体系,其中cDNA 1 μL(相当于25 ng 总RNA),2 μL 基因特异性引物,10 μL 2×SYBR Premix ExTaq,用去RNA 酶的ddH2O 补足20 μL。使用Roche 480 Ⅱ定量PCR 系统(瑞士)进行RT-PCR,反应条件为:94℃预变性2 min,94 ℃ 15 s,58 ℃ 30 s,72 ℃ 30 s,共40 个循环。每个样品3 次重复。采用2-ΔΔct法计算目标基因相对表达量,以根为对照。
2 结果与分析
2.1 荔枝LcALMT 家族成员基本信息及蛋白理化性质、亚细胞定位分析
本研究共鉴定出16个荔枝LcALMTs(LcALMT1~LcALMT16),其基本信息如表2。理化性质分析发现,LcALMTs 蛋白编码的氨基酸数目在118~803 aa,分子量大小在12.60~89.54 kD,等电点在5.16~9.07 之间,其中碱性蛋白(pI>7)10 个、酸性蛋白(pI<7)6 个,不稳定系数介于25.57~45.17,且LcALMT1、LcALMT13、LcALMT14、LcALMT16属于不稳定蛋白(不稳定系数>40)。亚细胞定位预测显示,该家族成员蛋白均定位于质膜上。
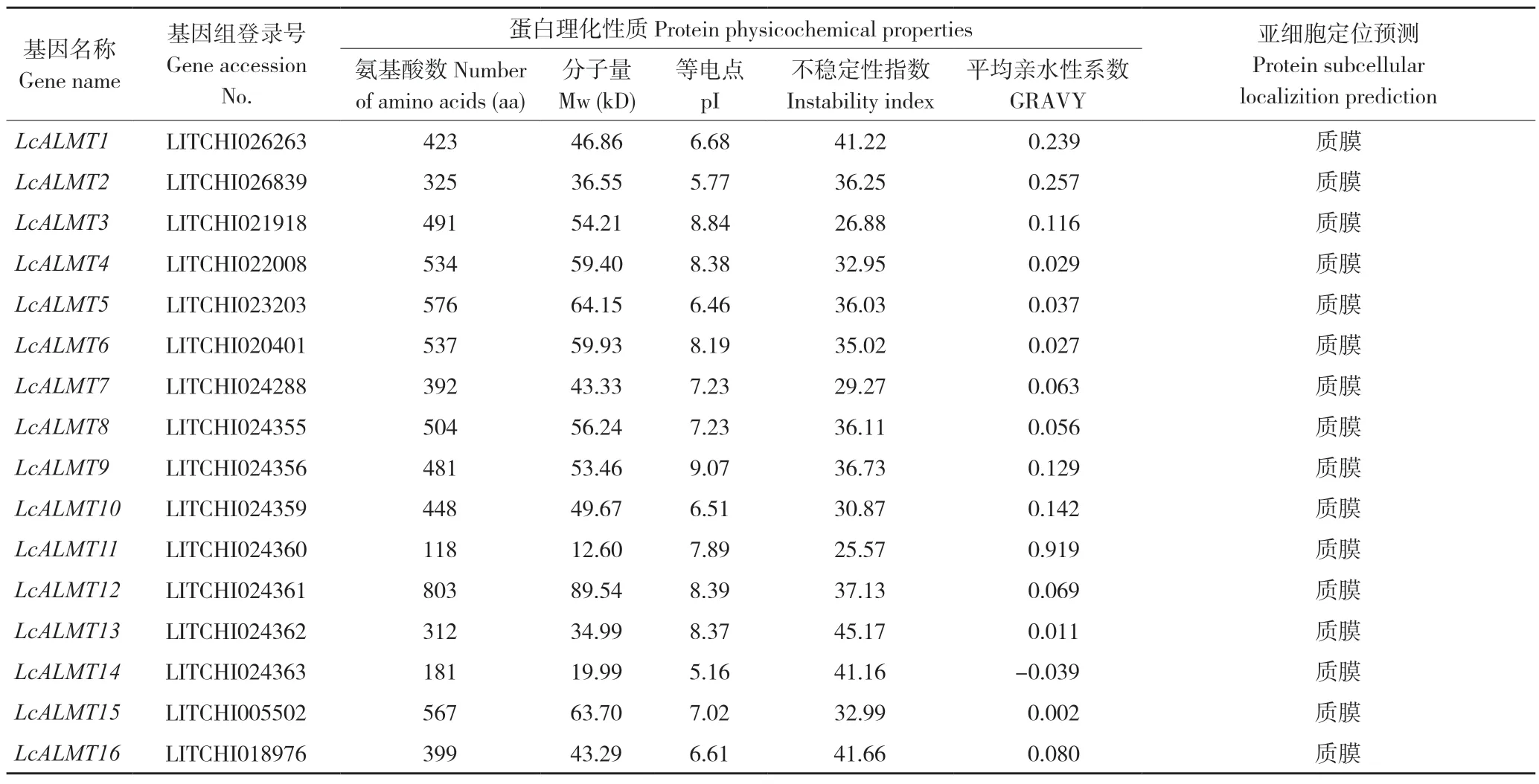
表2 荔枝LcALMTs 基本信息及其蛋白理化性质、亚细胞定位分析Table 2 Basic information of LcALMTs in litchi and analysis of its protein physicochemical properties and subcellular localization
2.2 荔枝LcALMTs 系统进化树及蛋白结构分析
使用MEGA 11 生成荔枝LcALMTs 系统进化树图,由图1 可知,该家族成员可分为5 个亚族。第Ⅰ、Ⅱ、Ⅴ亚族均包含2 个LcALMT 成员,第Ⅳ亚族包含3 个LcALMT 成员,第Ⅲ亚族包含7个LcALMT 成员。荔枝LcALMTs 成员在5 个亚族中分布不均匀,主要在第Ⅲ亚族。从进化树来看,荔枝ALMT 家族成员与拟南芥的ALMT 家族成员亲缘关系更近。
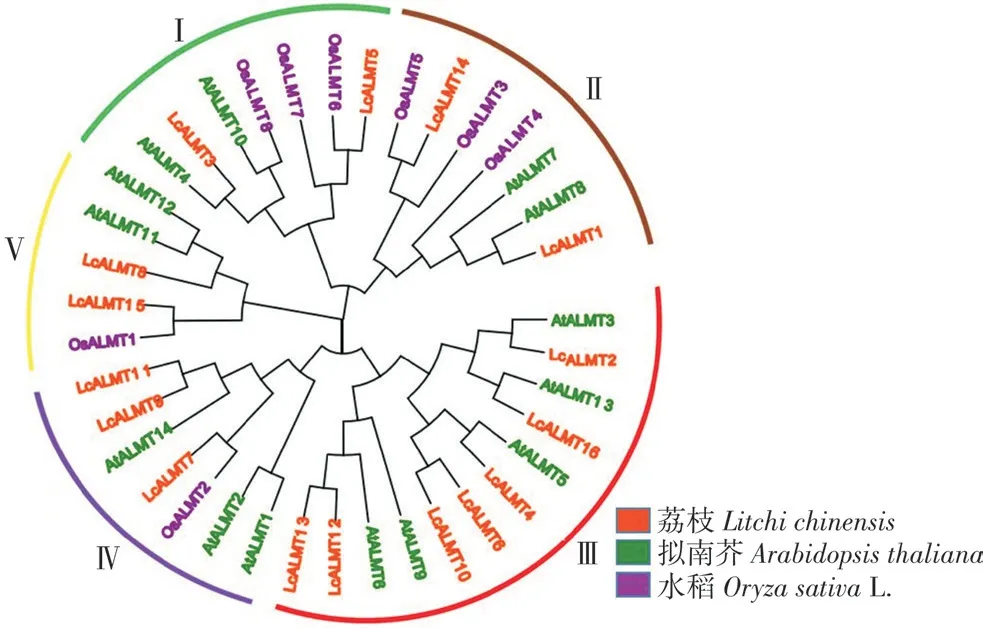
图1 荔枝、拟南芥和水稻ALMTs 系统进化树Fig.1 Phylogenetic tree of ALMT proteins in Litchi chinesis,Arabidopsis thaliana and Oryza sativa L.
对荔枝LcALMTs 蛋白二级结构进行分析发现,该家族蛋白二级结构包含α-螺旋、延伸链、无规则卷曲和β 转角(表3)。其中α-螺旋占比最高、均大于50%,延伸链占比为9.69%~19.50%,无规则卷曲占比为18.64%~57.32%,β 转角占比为1.94%~7.73%。蛋白质三级结构(图2)预测显示,位于同一亚族的LcALMTs 蛋白结构相似,如亚族I 中的LcALMT3 和LcALMT5,亚族Ⅲ中的LcALMT12、LcALMT13 和LcALMT16 均具有相似结构。
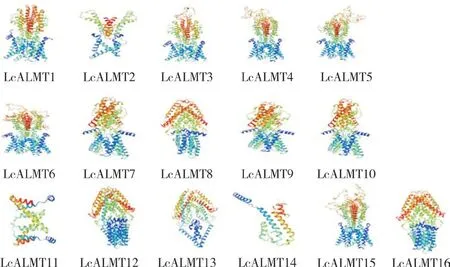
图2 LcALMTs 蛋白三级结构Fig.2 Tertiary structure of LcALMT proteins

表3 LcALMTs 蛋白二级结构Table 3 Secondary structure of LcALMT proteins
2.3 荔枝LcALMTs 基因结构及保守基序分析
运用GSDS 软件对荔枝ALMT家族成员编码区、内含子和非编码区进行结构分析,由图3 可知,荔枝LcALMTs外显子数目在3~10 个,其中LcALMT12外显子数目最多(10 个);LcALMT1、LcALMT3、LcALMT4、LcALMT5、LcALMT6、LcALMT8和LcALMT15均含有6 个外显子,LcALMT7、LcALMT10、LcALMT11、LcALMT14基因均不含有非编码区。
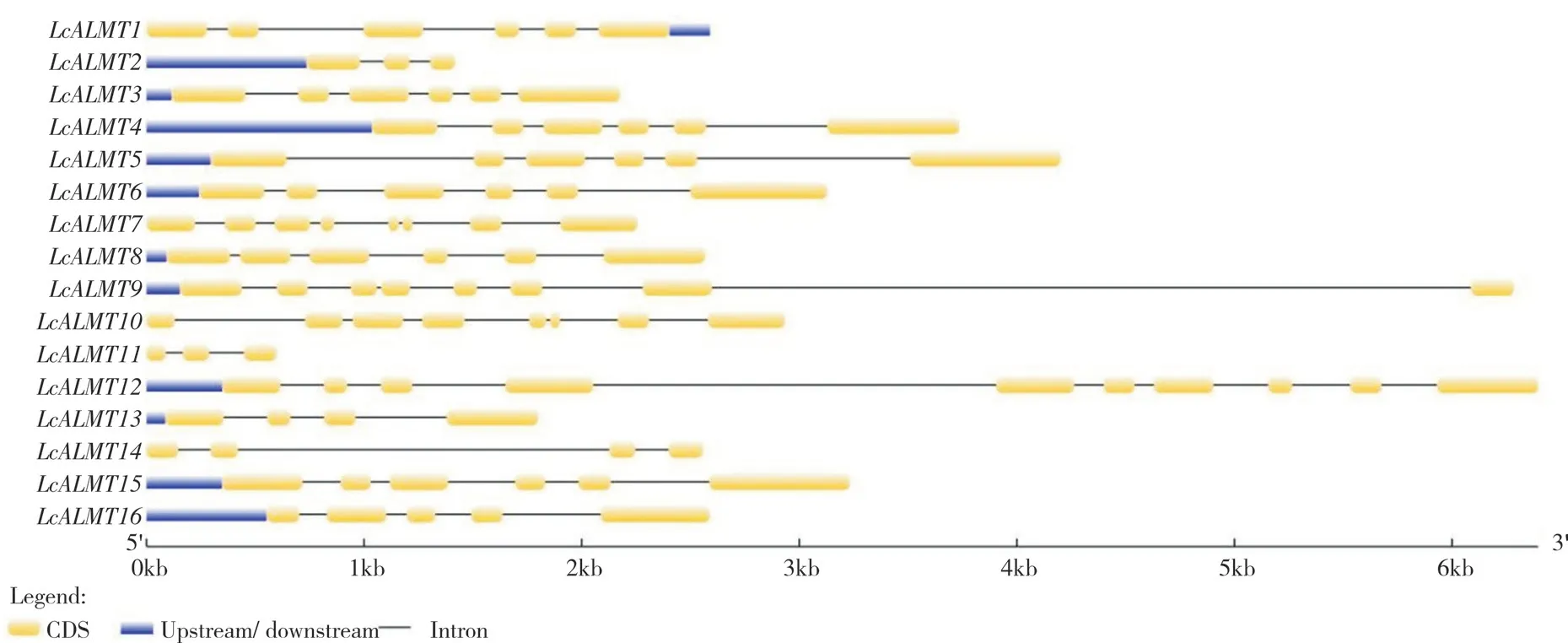
图3 荔枝 LcALMTs 基因结构Fig.3 Gene structure of LcALMTs in litchi
利用MEME 在线网站分析保守基序,基序数量上限设置为12,其他参数不变,结果(图4A)显示,有13 个LcALMTs 蛋白均含有motif 1;除LcALMT11 外,其他蛋白均含有motif 4;LcALMT11 和LcALMT14 仅含有2 个保守基序,分别为motif 11、motif 5 和motif 6、motif 4。由NCBI 网站分析得知荔枝ALMT 家族含有ALMT和ALMT superfamily 两个结构域,由图4B 可知,15 个LcALMTs 蛋白均只含有1 个结构域,而LcALMT12 包含上述2 个结构域,其中ALMT superfamily 结构域位于N 端、ALMT 结构域位于C 端。由SMART(http://smart.embl.de/)网站分析可知,motif 1~motif 7 均属于ALMT 结构域,motif 8~motif 12 未知。
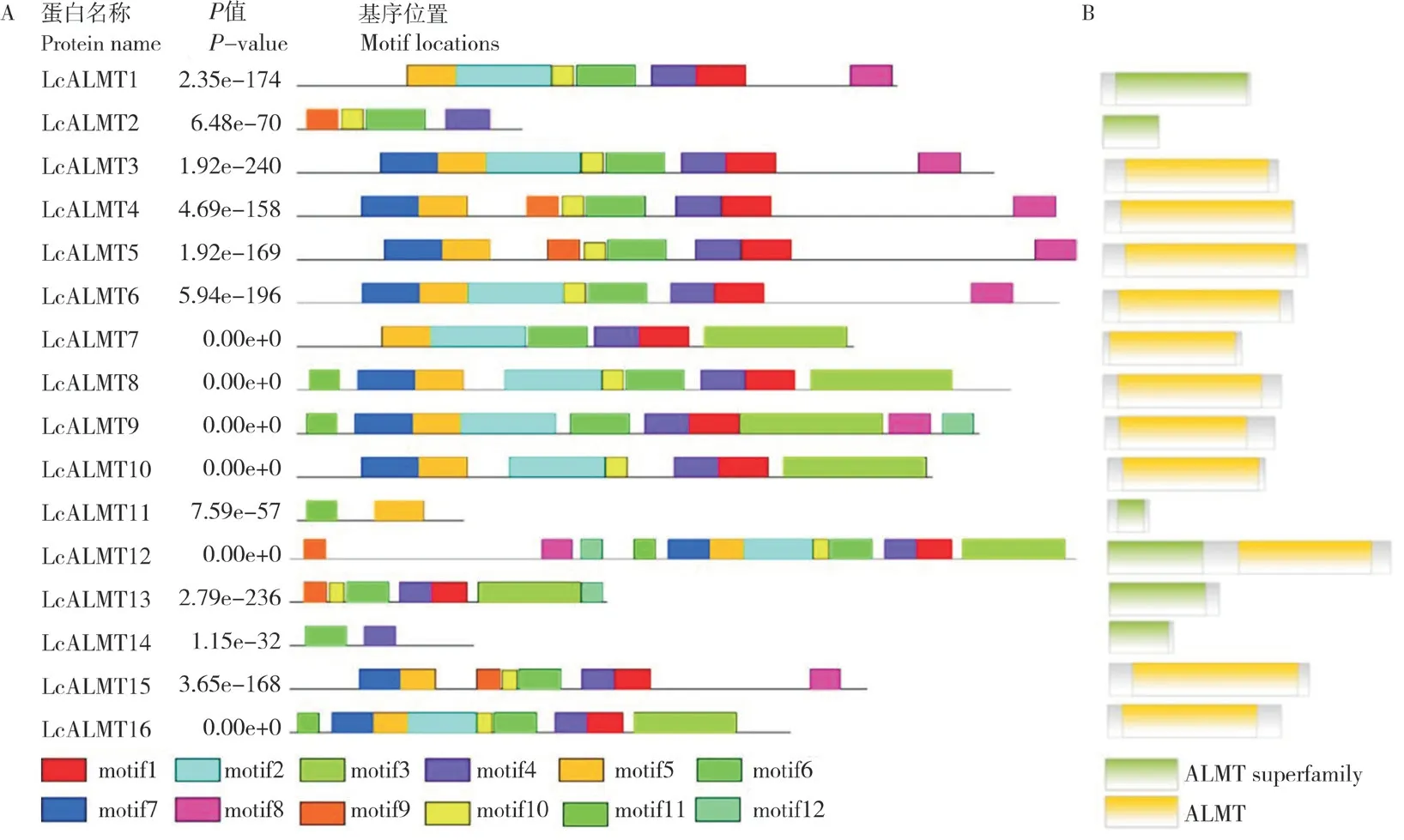
图4 荔枝LcALMTs 蛋白保守结构域Fig.4 Conservative domains of LcALMT proteins in Litchi
2.4 荔枝LcALMTs 的染色体定位分析
染色体定位分析结果(图5)表明,LcALMT基因家族只定位在荔枝15 条染色体中的6 条染色体上,其中LcALMT1和LcALMT2定位在Chr3 上,LcALMT3、LcALMT4和LcALMT5定位在Chr10 上,LcALMT6、LcALMT15和LcALMT16分别定位在Chr12、Chr14 和Chr15 上,Chr13 上LcALMT 蛋白分布最多且紧密。

图5 荔枝LcALMTs 基因染色体定位Fig.5 Chromosomal localization of LcALMT genes in Litchi
2.5 荔枝LcALMTs 启动子上顺式作用元件分析
利用Plantcare 在线网站对LcALMTs基因5′端上游2 000 bp 的序列预测顺式作用元件,除去一般性调控转录元件(如TATA-box、CAATbox)等未知功能元件外,共发现536 个顺式作用元件,其中光响应元件最多、共有219 个(占40.86%),荔枝LcALMTs基因均至少含有5 个光响应元件,如TCT-motiff、G-box、Box4 和AT1-motif 等(图6)。其次是激素响应元件、共有216 个(占40.30%),主要包括脱落酸响应元件(ABRE)、水杨酸响应元件(TCA-element)、茉莉酸甲酯响应元件、赤霉素响应元件和生长素响应元件。其中LcALMT7含有的茉莉酸甲酯响应元件最多、有94 个。整个LcALMT家族成员中,LcALMT4不含脱落酸响应元件。生长发育元件有30 个(5.60%),主要包括胚乳表达元件、玉米醇溶蛋白代谢调节相关元件、栅栏叶肉细胞分化元件、种子特异性调控元件和参与细胞周期调控元件。非生物胁迫元件有44 个(8.21%),主要包括防御应激响应元件TC-rich repeats、无氧诱导相关元件ARE、低温响应元件LTR 和诱导子激活元件。其他元件有27 个(5.04%),包括光响应MYB 结合位点、干旱胁迫诱导MYB 结合位点、蛋白结合位点、ATBP-1 结合位点和MYBHv1 结合位点。以上结果表明,荔枝LcALMTs成员可能具有差异化的生物学功能,参与荔枝的光响应、激素响应、生长发育与胁迫响应等相关过程。

图6 荔枝LcALMTs 启动子上顺式作用元件分析Fig.6 Analysis of cis-regulatory elements on LcALMTs promoter in Litchi
2.6 荔枝LcALMTs 的组织表达
从荔枝基因组数据库中LcALMT家族成员在不同组织中的表达量(图7)来看,LcALMT5和LcALMT15在不同组织中都有表达且表达量均较高,其中LcALMT5在雄花中的表达量最高,LcALMT15在假种皮中的表达量最高、是果皮中的16.9 倍。LcALMT10和LcALMT14在荔枝的各组织中均未检测出表达。LcALMT3只在雄蕊中有表达,且表达量极低。LcALMT4在种子中的表达量最高、是叶中表达量的13.6 倍,在不育雄蕊中未检测出表达。挑取LcALMT家族成员中表达量较高的3 个基因LcALMT4、LcALMT5和LcALMT15进行荧光定量验证,分析妃子笑不同组织(根、嫩叶、老叶、雄花、雌花、果皮、果柄)中LcALMTs的表达情况,以根的表达量为1、各基因在各部位下的表达量与其比较进行定量,结果(图8)显示,LcALMT4、LcALMT5和LcALMT15在不同组织中均有表达,但存在表达量的差异。LcALMT4、LcALMT5和LcALMT15均在雄花中表达最高、其次为雌花。LcALMT5的表达水平与转录组结果较为一致。
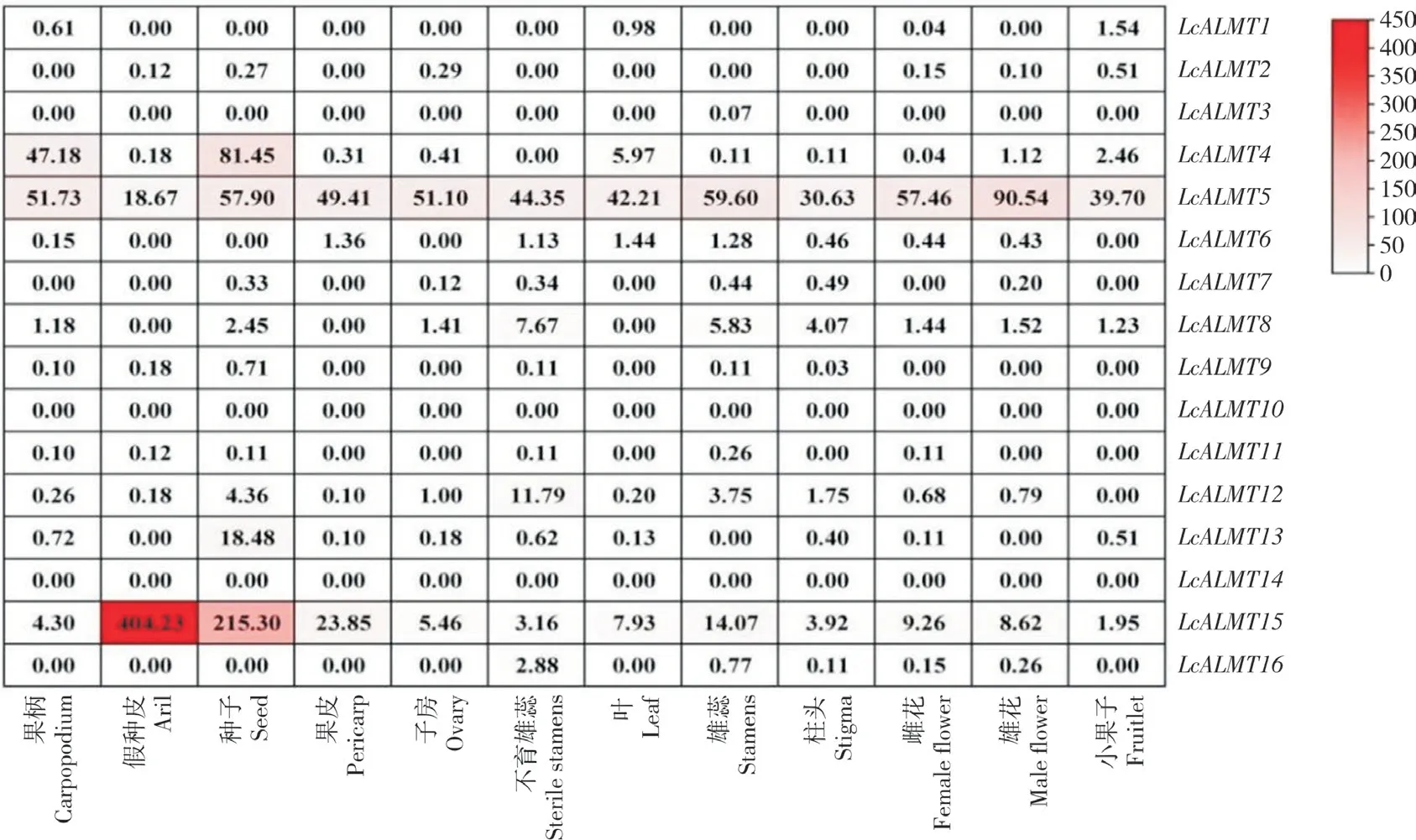
图7 荔枝LcALMTs 基因表达模式分析Fig.7 Expression patterns analysis of LcALMT genes in Litchi

图8 荔枝LcALMT4/5/15 相对表达水平Fig.8 Relative expression levels of LcALMT4/5/15 in Litchi
3 讨论
本研究对荔枝ALMT基因的全基因组进行鉴定与表达分析,对其部分功能进行预测,可为研究荔枝生物学功能奠定一定基础。ALMT 广泛存在于植物中,参与植物的生理过程,其功能也相应被探索,如植物耐铝性[17-19]、矿质营养[20-21]、果实酸度[22-23]、气孔运动[24-27]、参与种子发育[28-29]等。第1 个植物基因TaALMT1[6]被发现后,越来越多的ALMT 家族成员在不同植物中也相应被研究,但在荔枝中未见到ALMT基因家族的报道。本研究从荔枝基因库得到ALMT 数据,对其ALMT 家族成员进行鉴定,并系统分析该家族成员的蛋白理化性质、系统发育、基因结构、染色体定位、保守基序、顺式作用元件、蛋白结构和表达模式等。
从荔枝基因组中鉴定出16个LcALMTs基因,数量与草莓相同、多于拟南芥(14 个)[9]、少于苹果ALMT 家族成员[14]。荔枝ALMT 家族成员均定位于质膜上,与苹果中MdALMT13 在烟草亚细胞定位结果[15]一致,均定位于质膜上。荔枝ALMT家族成员外显子数目在3~10 个,苹果中ALMT家族成员外显子数量为4~12 个[15],则两者之间外显子数量差距较大,这可能是荔枝在进化过程中发生了外显子的缺失。染色体只定位在荔枝15 条染色体中的6 条染色体上,Chr13 上LcALMTs分布最多且紧密,与萝卜ALMT家族成员定位在9 条染色体上有差异[18],具体原因还需进一步探究。荔枝ALMT基因家族成员含有多种顺式作用元件,均包含光响应元件,激素响应元件生长发育元件,且光响应元件数量最多,暗示LcALMTs基因的表达可能受光照、激素调节和生长发育条件的影响且受光照影响最大,这与前人在其他物种中的研究结果一致。LcALMT7基因含茉莉酸响应元件最多、达94 个,表明该基因的表达量可能主要受茉莉酸的调控,故不同基因成员之间表达量不同主要与光照、激素调控,与生长发育条件有着巨大关系。该家族成员蛋白结构中α-螺旋占比最高,位于同一亚族的ALMT蛋白一般结构相似。系统进化分析发现,荔枝ALMTs 蛋白可以分为5 个亚族,这与萝卜ALMT家族分类[30]一致。荔枝ALMT家族成员在不同组织中的表达量不同,LcALMT4主要在种子中表达,与MdALMT13主要在根中高度表达结果[31]一致。采用qRT-PCR 对LcALMT4、LcALMT5、LcALMT15在荔枝不同组织中的表达水平进行检测,结果表明LcALMT4、LcALMT5、LcALMT15在各组织中均有表达,且在雄花中高度表达,推测LcALMT4、LcALMT5、LcALMT15可能参与雄花的调控,在雄花发育中起作用。综上,不同物种之间的基因在进化过程中多数存在相同或相似功能,表达量之间的差异不同,与生长环境和激素调节相关联。
4 结论
利用生物信息学方法从荔枝基因组中鉴定出16 个ALMTs成员,这些成员蛋白均定位于质膜上,染色体定位在荔枝15 条染色体中的6 条染色体上,且在Chr13 上LcALMTs分布最多且紧密。荔枝ALMT基因家族成员启动子含有多种顺式作用元件,光响应元件含量最多;蛋白结构中,α-螺旋占比最高;五个亚族中,位于同一亚族的ALMTs 蛋白一般结构相似。其相对表达量来看LcALMT4、LcALMT5和LcALMT15在不同组织中均有表达,且均在雄花中表达最高,其次为雌花。
猜你喜欢
科学大众(2022年23期)2023-01-30
江苏农业科学(2022年17期)2022-09-24
四川大学学报(自然科学版)(2021年4期)2021-07-15
中国农业科学(2020年5期)2020-03-30
中国(俄文)(2019年8期)2019-08-24
意林·全彩Color(2019年4期)2019-05-11
广州大学学报(自然科学版)(2019年1期)2019-05-07
岭南音乐(2017年2期)2017-05-17
上海师范大学学报·自然科学版(2016年5期)2016-12-14
天津科技大学学报(2016年1期)2016-02-28
