
温和灸足三里穴对自然衰老大鼠肾组织p16 DNA 甲基化的影响
2024-03-02 07:59刘文博赵舒羽殷之光杨秋红崔云华黄艳焦丹丽吴焕淦赵琛
上海针灸杂志 2024年2期
刘文博,赵舒羽,殷之光,杨秋红,崔云华,,黄艳,,焦丹丽,吴焕淦,,赵琛
(1.上海中医药大学附属岳阳中西医结合医院,上海 200437;2.上海市针灸经络研究所,上海 200030;3.上海中医药大学,上海 201203)
衰老是人类生活中一个自然而渐进的过程,衰老紧张过程具有伤害性物质累积、生理功能完整性丧失,死亡易感性增加的特点[1]。目前针对衰老的研究集中在长寿蛋白修饰、表突变、甲基化、代谢副产物等[2-4]。衰老是年龄增长的动态变化过程,随着老龄化过程的推进,多种慢性疾病伴随发生,对老年人的身心健康、认知能力等都构成了威胁,严重影响老年人生活质量,因此,对于延缓老龄化,改善衰老进程的研究亟需关注[5-6]。随着人口老龄化的增长,增龄性疾病的发病率逐渐增加,肾脏是受衰老影响最明显的器官之一, 随着年龄增长,肾脏代偿和细胞修复的能力下降,与其他年龄相关的危险因素结合,使得老年性肾病风险增大,故肾脏已渐渐成为了目前研究衰老的主要器官之一,但目前尚无明确针对肾组织衰老及相关疾病的治疗方法。课题组前期研究[7-9]发现,衰老大鼠存在肾组织炎症性改变,温和灸干预可降低衰老大鼠血清BUN 水平,改善衰老大鼠的肾小球结构,减轻肾组织炎症;同时温和灸足三里穴可抑制肾组织FN 表达、TGF-β1 基因转录,延缓肾脏衰老。DNA 甲基化是近年来研究最多、最为广泛的表观遗传现象之一,在人体生长发育和衰老过程中都起着极为重要的作用,其动态变化被认为是衰老主要标志之一[10-11]。DNA 甲基化不仅与多种衰老相关性疾病有密切联系,而且可以预测生物预期寿命和死亡风险[12]。目前针对DNA 甲基化参与肾脏衰老发生发展的机制尚不清楚。p16 作为抑癌基因,是细胞衰老的关键性效应物,在衰老发生发展中起着非常重要的作用。研究[13-15]发现,艾灸可以通过调控p16 的表达延缓衰老,但其调控p16 延缓衰老的具体机制的研究鲜有报道。基于此,本研究选取自然衰老大鼠模型,衰老典型器官(肾组织)进行靶向研究,以经典的大鼠衰老相关β-半乳糖苷酶作为肾脏衰老的生物学指标,通过观察温和灸足三里穴对自然衰老大鼠肾组织衰老生物学指标的影响,并进一步观察大鼠肾组织衰老相关甲基转移酶及甲基化程度,研究温和灸足三里穴对肾组织衰老的作用机制。
1 材料与方法
1.1 实验动物
选用3 月龄,雄性清洁级Sprague-Dawley (SD)大鼠30 只,体质量(350±20)g(上海中医药大学动物实验中心提供,合格证号2008001646891);饲养条件为温度(22~27)℃,相对湿度为50%~70%,昼夜节律12 h/12 h。所有大鼠自由饮水进食,进行适应性饲养,1 周后如无不良反应,饮食饮水正常,则可纳入正式试验。所有实验开展前均获得上海中医药大学动物伦理委员会批准(批准号PZSHUTCM18120705)。
1.2 主要仪器与试剂
特制细艾条(南阳汉医艾绒有限责任公司);戊巴比妥钠(Sigma 公司,美国,货号P3761);无水乙醇(国药集团化学试剂有限公司,货号10009218);多聚甲醛(国药集团化学试剂有限公司,货号80096618);二甲苯(国药集团化学试剂有限公司,货号10023418);中性树胶(国药集团化学试剂有限公司,货号10004160);十二水合磷酸氢二钠(国药集团化学试剂有限公司,货号200040618);磷酸二氢钠(国药集团化学试剂有限公司,货号200407008);氯化钠(国药集团化学试剂有限公司,货号10019308);盐酸(国药集团化学试剂有限公司,货号80070560);β-半乳糖苷酶一抗(Santa Cruz 公司);TBA 试剂盒(南京建成生物研究所);SYBR Green PCR 试剂盒(Thermo 公司);DNA 提取试剂盒(Tiangen Biotech 公司提供);酶联免疫吸附法试剂盒(上海源叶生物有限公司);干式恒温器(杭州蓝焰科技有限公司);冷藏冰箱(伊莱克斯电器有限公司);Real-Time 检测仪(ABI 公司);离心机(上海卢湘仪离心机仪器有限公司);移液枪(吉尔森P 型移液器公司);旋涡振荡器(青浦泸西仪器厂);电动匀浆机(FLUKO 公司);冷冻离心机[BBI 公司(Canada)];电子数显温度计(上海医用仪器厂);p16 抗体(abcam 公司,货号ab54210);Dnmt3a 抗体(abcam 公司,货号ab188470);Dnmt3b 抗体(abcam 公司,货号ab2851);二抗试剂盒(武汉博士德公司)。
1.3 分组与干预方法
雄性SD 大鼠分为24 月龄组和艾灸组,每组10 只。24 月龄组饲养至24 月龄,不进行治疗。艾灸组从大鼠12 月龄开始温和灸足三里穴(穴位定位方法参照《实验针灸学》[16]相关标准),采用直径5 mm 小艾条点燃后在距离穴位2 cm 处施灸,电子数显温度计在大鼠穴位局部监测体表温度,维持在(40±1)℃(通过手动方法上下移动,控制局部穴区温度),左右两侧足三里穴交替施灸,每次20 min,每日1 次。连续温和灸12 个月,直至大鼠24 月龄。另设12 月龄组(12 月龄雄性SD 大鼠10 只)作为对照。
1.4 标本采集
实验结束后,大鼠禁食24 h、不禁水。用2%戊巴比妥钠50 mg/(kg·bw)行腹腔注射,待大鼠麻醉后,冰浴状态取肾组织,一部分(约1 cm3大小)置于多聚甲醛中固定,另一部分置于标记好的冻存管中,投入液氮中保存待测。
1.5 观察指标
1.5.1 免疫荧光检测大鼠肾组织β-半乳糖苷酶表达
治疗结束后,3 组随机选择大鼠,取各大鼠肾组织,制备组织切片,然后进行免疫荧光染色,经过冰冻切片、脱蜡水化、滴加荧光标记的一抗(β-半乳糖苷酶)、二抗、DAPI 覆盖组织切片染色步骤,最后通过荧光显微镜进行拍片。
1.5.2 qRT-PCR 法检测3 组大鼠肾组织端粒长度、p16 mRNA 、 衰老相关甲基转移酶(DNMT3a 和DNMT3b)mRNA 表达
取新鲜大鼠肾组织,加入蛋白酶溶解,与肾组织充分混合,后续依次加入缓冲液GB、无水乙醇、CB3 并分别离心,同时重复操作2~3 次,提取组织中DNA;后续根据设置好的扩增体系依次加入试剂,在特定反应程序下操作,取得结果后通过CT 值进行分析,端粒(T)重复拷贝数与单拷贝基因(S)的比率,即T/S 比率可以得出端粒的相对长度,而T/S 比率与端粒相对长度成正比关系。T/S 计算公式如下,T/S =[2CT(telomeres)/2CT(single copy gene)]=2-ΔCT。肾组织端粒长度 mRNA 检测引物序列设计为 Primer F:5’GGTTTTTGAGGGTGAGGGTGAGGGTGAGGGTGAGGGT 3’,rimer R:5'TCCCGACTATCCCTATCCCTATCCCTATCCCTATCCCTA-3';肾组织 p16 mRNA 检测引物序列设计为 Primer F:5'CGTGCGGTATTTGCGGTATC 3',Primer R: 5' TGCCAGAAGTGAAGCCAAGG 3';肾组织Dnmt3a mRNA 检测引物序 列 设 计 为 Primer F 5' ATTACCACCAGGTCAAACTC,Primer R 5' CCAAACACCCTTTCCATTTC 3;肾组织Dnmt3b mRNA 检测引物序列设计为Primer F 5' TCCTGCGGTAAGAAGAACCC 3',Primer R 5' ACTGATAGCCGTCCTCATCG 3';GAPDH 引物序列设计为Primer F:5' GGAGTCTACTGGCGTCTTCAC 3',Primer R:5'ATGAGCCCTTCCACGATGC 3'。扩增体系,实时荧光定量PCR 预混液12.5 µL、Forwardprimer 0.5 µL。反应程序为95 ℃条件下,10 min(95 ℃条件下15 s;60 ℃条件下45 s;72 ℃条件下1 min)×40;95 ℃条件下15 s;60 ℃条件下1 min;95 ℃条件下15 s;60 ℃条件下15 s。
1.5.3 酶联免疫吸附法检测3组大鼠肾组织p16蛋白表达
取3 组大鼠肾组织匀浆上清进行酶联免疫吸附法(Rat CDKN2A/p16INK4a ELISA Kit, Catalog No. LS-F15224)检测。采用“三明治”法进行检测,将标准品添加至微孔板中,目标抗原与抗体结合,洗去未结合的样品。接着加入生物素结合的检测抗体,与相应抗原结合,洗去未结合的抗体。接着加入Avidin-HRP 标记的结合物、TMB 底物显色等。最后在(450±2)nm 波长下测量每孔的光学密度(OD)。通过生成标准曲线确定抗原浓度。
1.5.4 免疫组化法检测3 组大鼠肾组织衰老相关甲基转移酶(DNMT3a 和DNMT3b)蛋白表达
取3 组大鼠肾组织,经福尔马林固定后放入石蜡包埋并切片,将石蜡切片经脱蜡、水化、抗原修复、山羊血清封闭、滴加一抗(p16 abcam54210 1:800,Dnmt3a abcam188470,1:2 000,Dnmt3b abcam2851,1:300)、二抗、 DAB 显色、 中性树胶封片等步骤, 选用LEICADM6000B 光学显微镜镜下观察。切片背景为天蓝色,阳性表达呈黄褐色或者棕色。图像分析通过Imagepro-Plus6.0 图像分析系统进行图像定量测定。摄片时在每张切片中随机选取3 个视野,计算阳性积分光吸度值(IOD),通过公式AOD=IOD/area 计算平均光密度,最终结果取3 个切片的平均值。
1.5.5 重亚硫酸盐测序技术检测3 组大鼠肾组织p16 DNA CPG 岛甲基化程度
亚硫酸氢钠修饰基因组DNA,进一步通过PCR 扩增技术,连接产物转化、蓝白斑筛选、质粒提取、质粒测序等步骤,比对所测序列。
1.6 统计学分析
采用SPSS21.0 统计软件进行数据分析。计量资料数据服从正态分布,采用均数±标准差表示;不服从正态分布,则以中位数(四分位间距)表示。数据方差齐时,采用单因素方差分析,两两比较采用最小显著差法(LSD);方差不齐时采用Games-Howell 比较组间差异。数据不符合正态分布,组间比较采用Kruskal-WallisH检验,两两比较则采用Mann-WhitneyU检验。统计结果以P<0.05 表示差异有统计学意义。
2 结果
2.1 最终纳入实验3 组大鼠数量
12 月龄组大鼠死亡1 只,24 月龄组大鼠死亡2 只,艾灸组大鼠死亡1 只。故最终纳入实验的为12 月龄组9 只、24 月龄组8 只和艾灸组9 只。
2.2 3 组大鼠肾组织衰老标志物检测
2.2.1 3 组大鼠肾组织β-半乳糖苷酶的表达
镜下观察,大鼠肾组织内可看到β-半乳糖苷酶阳性表达。12 月龄组肾组织中阳性表达染色较淡,呈现弱阳性反应;24 月龄组肾组织中阳性表达面积较大,染色较深;艾灸组肾组织中阳性表达较衰老组弱,阳性表达面积小,染色偏淡。详见图1。
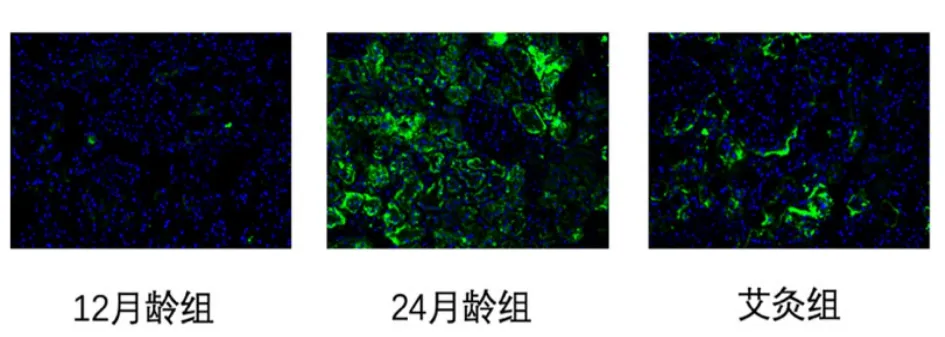
图1 3 组大鼠肾组织β-半乳糖苷酶表达(免疫荧光,×200)
2.2.2 3 组大鼠肾组织端粒长度变化
3 组大鼠肾组织端粒相对长度检测结果显示,与12 月龄组大鼠比较,24 月龄组大鼠肾组织端粒相对长度表达降低,差异具有统计学意义(P<0.05);与24 月龄组比较,艾灸组肾组织端粒相对长度表达升高,差异具有统计学意义(P<0.05)。详见图2。

图2 3 组大鼠肾组织端粒长度表达(12 月龄组n=9,24 月龄组n=8,艾灸组n=9)
2.3 3 组大鼠肾组织p16 表达
2.3.1 3 组大鼠肾组织p16 蛋白表达
3 组大鼠肾组织p16 蛋白表达免疫组化结果显示,p16 阳性表达染色呈棕黄色,在肾组织肾小球细胞、肾间质细胞及肾小管细胞胞核均可看到p16 蛋白表达。12 月龄组肾组织中p16 蛋白阳性表达染色较淡,呈弱阳性反应;24 月龄组肾组织中p16 蛋白表达较12 月龄组增多,呈阳性反应;艾灸组肾组织中p16 蛋白表达较24 月龄组降低,呈弱阳性反应。详见图3。

图3 3 组大鼠肾组织p16 蛋白表达(免疫组化,×200)
3 组大鼠肾组织p16 蛋白经酶联免疫吸附法检测结果显示,与12 月龄组大鼠比较,24 月龄组大鼠肾组织p16 蛋白表达增加(P<0.05);与24 月龄组比较,艾灸组p16 蛋白表达下降(P<0.05)。详见图4。
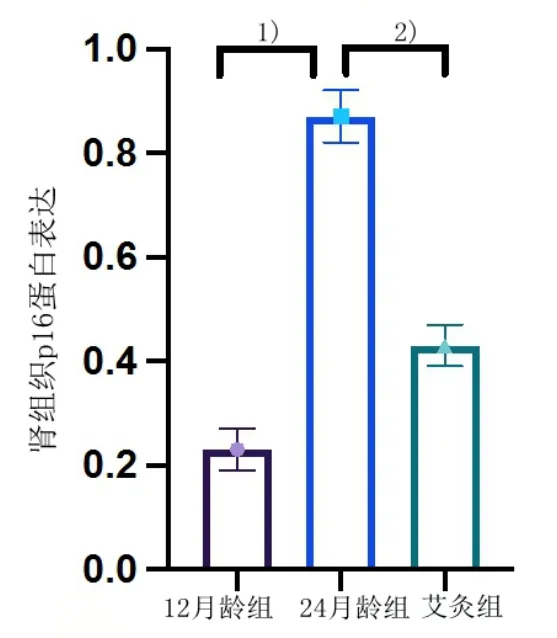
图4 3 组大鼠肾组织p16 蛋白表达(12 月龄组n=9,24 月龄组n=8,艾灸组n=9)
2.3.2 3 组大鼠肾组织p16mRNA 表达
3 组大鼠肾组织p16 mRNA 经qRT-PCR 检测结果显示,与12 月龄组大鼠比较,24 月龄组大鼠肾组织p16 mRNA 表达增加,差异具有统计学意义(P<0.05);与24 月龄组比较,艾灸足三里组p16 mRNA 表达下降,差异具有统计学意义(P<0.05)。详见图5。
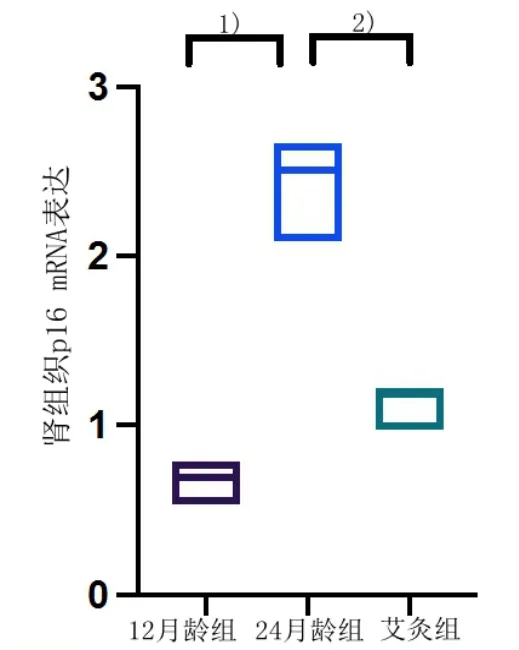
图5 3 组大鼠肾组织p16 mRNA 表达(12 月龄组n=9,24 月龄组n=8,艾灸组n=9)
2.4 3 组大鼠肾组织p16 DNA 甲基化程度
3 组大鼠肾组织p16 DNA CpG 岛甲基化率经BSP检测显示,3 组肾组织p16 DNA CpG 岛甲基化率比较,差异有统计学意义(P<0.05)。与12 月龄组和24 月龄组比较,艾灸组大鼠p16 DNA CpG 岛甲基化率升高,差异具有统计学意义(P<0.05)。详见图6。

图6 3 组大鼠肾组织p16 DNA CpG 岛甲基化(12 月龄组n=9,24 月龄组n=8,艾灸组n=9)
2.5 3 组大鼠肾组织衰老相关甲基转移酶表达
2.5.1 3 组大鼠肾组织衰老相关甲基转移酶蛋白表达
3 组大鼠肾组织Dnmt3a 和Dnmt3b 蛋白表达免疫组化结果显示,Dnmt3a 和Dnmt3b 阳性表达染色呈棕黄色,在肾组织肾小球细胞、肾小管细胞及肾间质细胞胞核均可看到Dnmt3a 和Dnmt3b 蛋白表达。12 月龄组肾组织中Dnmt3a和Dnmt3b阳性表达较多,染色相对较深,呈强阳性反应;24 月龄组肾组织中Dnmt3a 和Dnmt3b表达较低,呈弱阳性反应;艾灸组肾组织中Dnmt3a 和Dnmt3b 蛋白阳性表达较24 月龄组增加,呈阳性反应。详见图7。
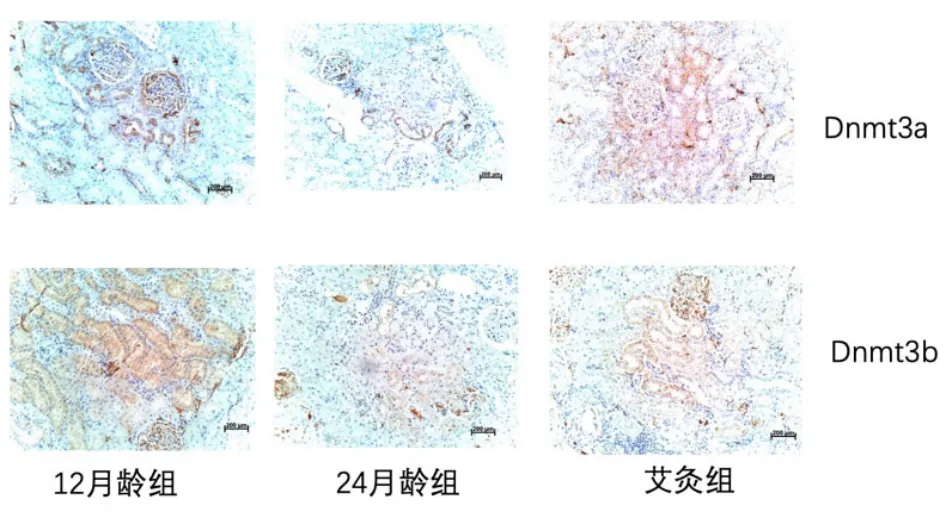
图7 3 组大鼠肾组织Dnmt3a 和Dnmt3b 表达(免疫组化,×200)
2.5.2 3 组大鼠肾组织衰老相关甲基转移酶mRNA 表达
3 组大鼠肾组织衰老相关甲基转移酶Dnmt3a 和Dnmt3b mRNA 表达经qRT-PCR 检测结果显示,与12 月龄组比较,24 月龄组大鼠Dnmt3a 和Dnmt3b mRNA 表达降低,差异具有统计学意义(P<0.05);与24 月龄组比较,艾灸组大鼠Dnmt3a 和Dnmt3b mRNA 表达增加,差异具有统计学意义(P<0.05)。详见图8 和图9。
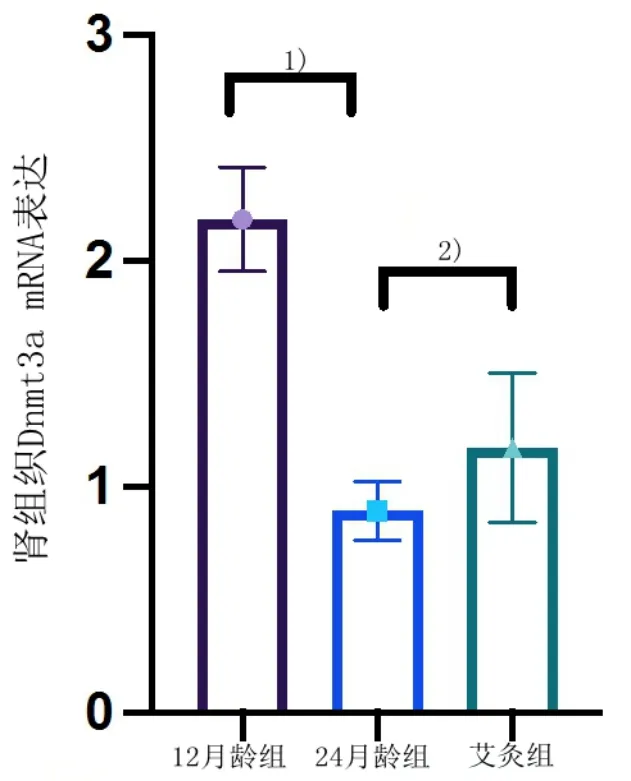
图8 3 组大鼠肾组织Dnmt3a mRNA 表达(12 月龄组n=9,24 月龄组n=8,艾灸组n=9)

图9 3 组大鼠肾组织Dnmt3b mRNA 表达(12 月龄组n=9,24 月龄组n=8,艾灸组n=9)
3 讨论
3.1 艾灸可影响衰老标志物的表达
随着人口日趋老龄化,衰老成为目前人们研究的热门话题。衰老本身是生命发展过程中不可逆的过程,但自古以来诸多医家不断寻求多种方式延缓衰老进程。肾为先天之本,早在《黄帝内经》中就提到肾气盛衰主宰人体生长、发育、衰老的生命过程,其中肾虚、肾阴阳不足是衰老的主要病机。传统灸法可通过借助灸火或辅以药物燃烧的热度及药物作用于相关腧穴以达到治未病和延年的作用。
艾灸一方面可使得穴位局部组织温度升高、能量代谢增加以及改善局部血液循环;另一方面可以影响机体远部,通过刺激体表经穴发挥作用。此外,艾灸可调节体内自由基代谢[17],增强机体免疫功能[18],调节神经内分泌[19-20],改善脂质代谢[21-22],调控不同组织内c-fos、Bcl-2、Bax 等衰老相关基因的表达[18],延缓衰老进程。因艾灸燃烧时产生温热作用影响局部经穴,结合操作情况,考虑到43 ℃以上的热刺激信号会对机体产生潜在性危害,动物在接受这个程度的热刺激时会产生逃避、跳跃等伤害性反应,因此本实验在经穴局部使用电子数显温度计进行温度监测,控制在(40±1)℃,进一步关注温和灸对衰老相关标志物及相关基因甲基化程度的影响。
随着人口老龄化进展,多种肾脏疾病发病率日益增高,针对肾脏衰老的研究近年来热度高涨,同时肾脏是受到衰老影响最明显的器官之一。特异性的衰老标志物的出现说明细胞进入了衰老状态,对衰老相关β-半乳糖苷酶(SA-β-Gal)的检测是能够说明细胞进入衰老周期强有力的证据。相关研究发现[23-24],在细胞所处环境pH 为6 时,其SA-β-Gal 表达随年龄增加而上调,且SA-β-Gal 敏感程度较高,在未发生衰老的细胞中检测不到,因此SA-β-Gal 是目前被广泛认可的检测细胞衰老的金标准[25-27]。足三里穴是常用的保健要穴,是足阳明胃经的合穴、胃之下合穴,为回阳九针穴之一,土经中的土穴,为后天之本。本实验在前人基础上,采用自然衰老模型鼠为研究对象,通过温和灸足三里穴,观察衰老相关指标及相关基因甲基化程度,从p16 DNA 甲基化角度观察温和灸足三里穴对自然衰老模型鼠肾组织的影响,探究其延缓衰老的机制。此外,衰老的进展过程中伴随端粒长度的损耗,研究发现端粒长度变化不仅影响肾脏衰老进程,而且与肾脏疾病、肾脏移植等有密切关系[28-29]。本研究发现,温和灸可降低肾组织内衰老相关β-半乳糖苷酶的表达,降低端粒长度的损耗。随着年龄的增长,肾脏的宏观和微观结构均会相应地发生变化。在微观结构水平上,肾脏的衰老主要表现在出现肾间质纤维化、肾小球硬化、基底膜增厚、肾小管萎缩和动脉硬化等病变。本课题组前期研究发现,艾灸可有效降低衰老大鼠肾组织衰老标志物SA-β-Gal 的表达,改善自然衰老大鼠的肾小球结构,减轻肾组织炎症,抑制肾间质纤维化;与此同时,温和灸可同时可降低衰老大鼠24 h尿蛋白含量及血清BUN 水平,通过抑制FN 表达及TGF-β1 基因转录发挥肾脏保护作用[7-8],对肾脏衰老进程产生影响。
3.2 艾灸可对衰老相关甲基转移酶表达的调控作用
DNA 甲基化是目前发现最早、研究较深入的重要表观遗传修饰作用之一,这一遗传方式在胚胎发育、染色体失活、细胞功能的维持、基因印记等过程中具有十分重要的作用。DNA 甲基化的实现主要依赖于Dnmts的催化作用,相关Dnmts 的表达变化可直接或间接对DNA 甲基化状态产生影响。哺乳动物的Dnmts 包含两个结构和功能不同的家族,即Dnmt1 和Dnmt3。Dnmt1在生物体内表达广泛,参与甲基化模式的复制过程;Dnmt3 家族包括Dnmt3a 和Dnmt3b,其表达在胚胎发生的不同阶段,具有组织特异性,同时Dnmt3b 作为重要的从头甲基化酶,可催化基因甲基化的新生位点[30-32]。Dnmt3b 可结合至分泌性卷曲相关蛋白1(secreted frizzled-related protein 1, SFRP1)基因启动子区域,诱导CpG 岛发生高甲基化,抑制SFRP1 基因转录,参与肾组织纤维化进程的调控[33]。此外,吴丽云等[34]通过研究发现,左金丸干预可以影响人体肠组织中Dnmts 的表达,对其异常表达可以有一定的调整作用。殷秋忆[35]通过研究发现,六味地黄方可调整雄性大鼠血清同型半胱氨酸水平进一步对其血管内皮细胞Dnmts 表达,起到抗内皮损伤的作用。因此传统中药可能通过多种路径对组织器官内甲基转移酶的表达进行调控,进一步对相应基因的表达产生干预作用,对细胞生长增殖产生影响。相关研究发现,年龄相关的去甲基化与甲基转移酶表达的下调有关。DNA 甲基化内源性调控因素主要包括DNA 甲基转移酶和去甲基化酶活性水平及体内其他因素对DNA 甲基化活性的调节。随着年龄增长,甲基转移酶的表达量显著减少,此外体内重要激素如生长激素、性激素水平的变化也会对甲基转移酶的活性起到重要干预作用并影响衰老相关进程。本研究发现,艾灸干预可明显增加衰老大鼠肾组织内Dnmt3a 和Dnmt3b 的表达,提示艾灸足三里穴可提高衰老大鼠肾组织内甲基转移酶的表达,这可能是艾灸延缓衰老的重要机制。
3.3 艾灸可通过调控p16 DNA CpG 岛甲基化发挥延缓衰老进程的作用
DNA 甲基化作为一种复制后修饰,主要出现在原核生物和真核生物基因组中,体内多种重要的生理过程均有其参与,对基因表达的调节,染色体完整性的维持及染色体灭活等都起到了十分重要的干预作用。DNA甲基化与肿瘤、遗传性疾病和衰老的发生具有十分密切的关系。甲基化模式非常不稳定,并受多种刺激如理化刺激、衰老、疾病等多种因素的影响。甲基化的DNA与甲基CpG结合蛋白二者结合(甲基CpG结合蛋白具有与组蛋白去乙酰化酶蛋白复合体相结合的性质),因此在两种蛋白参与的情况下,细胞染色体聚簇成团,导致DNA 转录活性丧失,细胞生长阻滞,出现细胞衰老[36]。本实验研究发现,12 月龄组大鼠p16 DNA CpG 岛整体甲基化水平与24 月龄组未见明显差异,提示肾组织衰老相关p16 DNA CpG 岛甲基化程度在大鼠老年前期即趋于稳定,从老年早期开始的持续的温和灸足三里穴可提高p16 DNA CpG 岛整体甲基化水平,降低p16 mRNA及p16 蛋白在衰老机体的表达,这可能是温和灸参与调控衰老进程的重要机制之一。
4 结论
艾灸可通过影响衰老相关甲基转移酶的表达,调控p16 DNA CpG 岛整体甲基化水平,降低p16 mRNA 及p16 蛋白在衰老机体的表达,从而发挥延缓衰老的生物学效应。
猜你喜欢
保健与生活(2022年11期)2022-06-09
服饰导报·鞋世界(2021年4期)2021-05-17
生物学通报(2020年10期)2020-08-13
癌变·畸变·突变(2019年3期)2019-06-03
中国药理学通报(2019年5期)2019-01-11
国际妇产科学杂志(2016年2期)2016-06-16
健康管理(2016年2期)2016-05-30
恋爱婚姻家庭·养生版(2016年5期)2016-05-06
分析测试学报(2015年8期)2016-01-13
河南医学研究(2012年3期)2012-08-15
