
视网膜色素变性患者视紫红质基因突变分析
2011-04-20 06:58贾晓林李进军卢立志刘晓玲
中国全科医学 2011年15期
刘 逾,贾晓林,李进军,卢立志,刘晓玲
视网膜色素变性 (Retinitis pigmentosa,RP)是视网膜感光细胞退化、视网膜色素上皮变性、外层视网膜进行性退化所致的常见遗传性致盲眼底病。在世界范围内该病发病率约为1/3 500[1],我国群体患病率约为 1/3 784[2],且呈逐年上升趋势,已成为临床上最常见且危害最严重的致盲性眼遗传病之一。RP的遗传方式复杂多样,非综合征RP中至少有4种遗传方式,其中30%为常染色体显性遗传 (ADRP),20%为常染色体隐性遗传 (ARRP),15%为X连锁遗传 (XLRP),另有5%早发型RP即隐性Leber's先天性黑蒙 (LCAⅡ型)。当患者因缺乏家族史而无法确定其遗传方式时,将其定义为散发型RP(SRP),约占30%[3]。多数RP为单基因遗传,但也存在二基因-双等位基因和二基因-三等位基因遗传方式[4]。迄今为止,通过连锁分析和候选基因筛选等方法已确定了19个ADRP、27个ARRP和6个XLRP基因位点,其中35个基因已被克隆。视紫红质 (RHO)基因是最早发现的ADRP致病基因,同时也是 ADRP患者最常见的致病基因。大约26.5%的ADRP患者由RHO基因突变引起[3],亚洲报道的突变率低于欧美,我国ADRP患者约有7%由RHO基因突变引起[5]。文献报道RHO基因突变也可导致ARRP,不过发生频率较小,不足1%[6]。为进一步明确RHO基因在中国人中的突变特征与频率,本研究对收集的RP患者进行RHO全基因多聚酶链式反应 (PCR)扩增,直接双向测序及基因突变检测分析,以期获得中国RP患者RHO基因突变频率及特征,探讨其在RP发病机制中的作用。
1 对象与方法
1.1 研究对象 选取2008年3月—2010年10月温州医学院附属眼视光医院门诊收治的55例RP先证者为研究对象,均为汉族,男31例,女24例,初诊年龄5~71岁,平均年龄35岁,其中ADRP 6例、ARRP 14例、SRP 35例,分布于中国华南地区。RP诊断的确立依据国内外通用标准:(1)夜盲史;(2)视力逐渐下降;(3)眼底检查可见骨细胞样或不规则的色素沉着,视网膜血管缩窄和视盘颜色蜡黄或变淡;(4)全视野视网膜电图 (ERG)明、暗视活动时,杆体细胞功能降低,有时可见锥体细胞功能轻度降低,晚期杆体、锥体细胞功能均可降低。(5)视野缩窄或有环行暗点特征性改变和 (或)适应曲线阀值升高,或视网膜杆体相缺如[7]。同时本研究随机收集55名健康成年人为对照组,其中男35例,女20例,年龄18~50岁,平均年龄32岁。
1.2 研究方法
1.2.1 临床研究 全面调查病史并绘制家系图,进行详细的眼科检查,行ERG及视野检查。
1.2.2 样品采集 对RP患者及对照者行全面系统检查,排除其他系统疾病,采集静脉血5 ml,乙二胺四乙酸二钾 (EDTA-K2)抗凝、-70℃保存。根据世界医学会《赫尔辛基宣言》涉及人类受试者的医学研究的伦理原则及知情同意原则,所有研究对象均签署知情同意书。
1.2.3 RP组DNA提取 采用蛋白酶K法提取白细胞DNA,干燥回收的DNA用TE缓冲液溶解备用,-20℃保存。
1.2.4 引物设计 运用primer 5.00软件 (PREMIER Biosoft International)对RHO基因序列 (Genbank NC_000003.11)进行引物设计,扩增RHO基因全序列,交上海英俊生物技术公司合成 (见表1)。
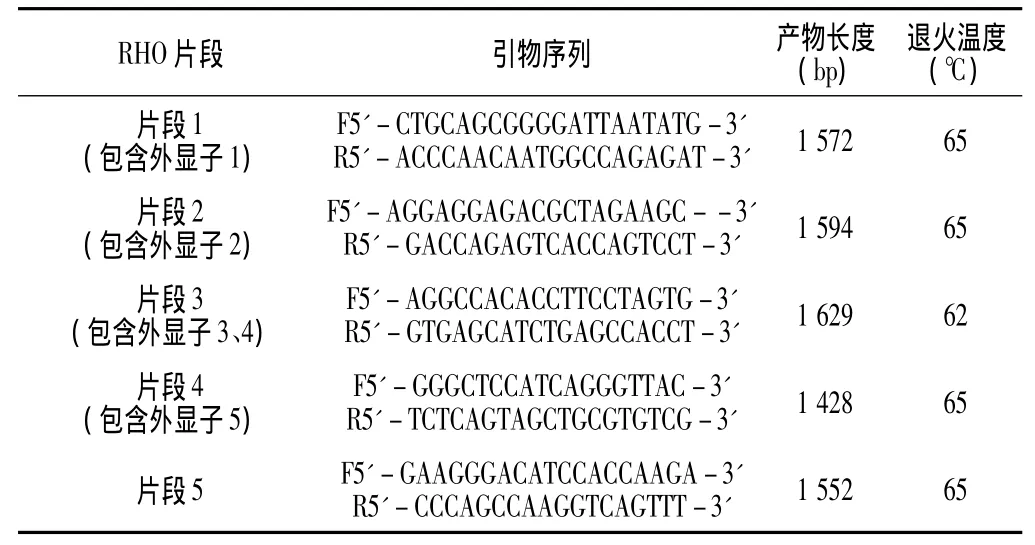
表1 RHO基因PCR引物碱基序列Table 1 PCR primer sequences for the RHO gene
1.2.5 PCR反应 PCR反应体系:10×Buffer 2.5μl,MgCl2(含 MgCl210 mmol/L)1.5 μl,dNTP(2.5 mmol/L)1.5 μl,ExTaq(5 U/μl)0.25μl,上下游引物 (10 mmol/L)各0.4 μl,DNA模板1.5μl,加超纯水至25μl。PCR反应条件:95℃预变性4 min,95℃变性30 s,63.5~66.0℃退火40 s,72℃延伸150 s,35个循环,后72℃延伸10min。PCR反应均在德国Biometra仪器上进行,反应试剂均来自大连宝生物公司(TaKaRa公司)。PCR反应结束后,将反应产物进行2%琼脂糖电泳,同时加Maker标记DNA相对分子量,透射紫外线观察DNA电泳带凝胶成像仪照相,计算机分析结果,以证实该扩增片段。
1.2.6 DNA测序分析 对扩增出的PCR扩增产物经纯化试剂盒纯化后进行直接的正、反双向测序分析 (上海英俊生物技术公司ABI3170自动测序仪)。采用Chromas2.31软件查看测序结果,并采用DNAStar软件 (DNAStar version 6,0,3,99)进行比对分析。突变的命名参照国际统一的人类基因突变命名系统[8]。
1.3 统计学方法 采用SPSS 15.0统计软件进行统计学处理。多重比较的P值由Bonferroni检验校正。采用χ2检验对每一个检测到的单核苷酸多态性 (SNP)是否符合Hardy-Weinberg平衡进行分析。基因型频率的比较采用χ2检验或Fisher精确概率法。以P<0.05为差异有统计学意义。
2 结果
2.1 PCR扩增产物 所有研究对象DNA经PCR扩增及2%琼脂糖凝胶电泳后,均显示单一条带,所扩增的片段长度与预期一致,表明是所需的产物 (见图1)。
2.2 DNA测序及分析结果 应用PCR产物直接测序技术,采用Chromas2.31软件查看测序结果,测序结果经DNAStar软件进行比对分析 (见图2),共检出7种碱基变异,均符合Hardy-Weinberg平衡。其中2种为错义突变,V104I仅在RP组出现,A299S在RP组及对照组中各有1例。其余5种为非编码区突变,且在对照组中也发现相应碱基突变,考虑为SNP,在NCBI-SNP数据库中进行检索,两组各SNP位点突变频率比较,差异均无统计学意义 (P>0.05,见表2)。
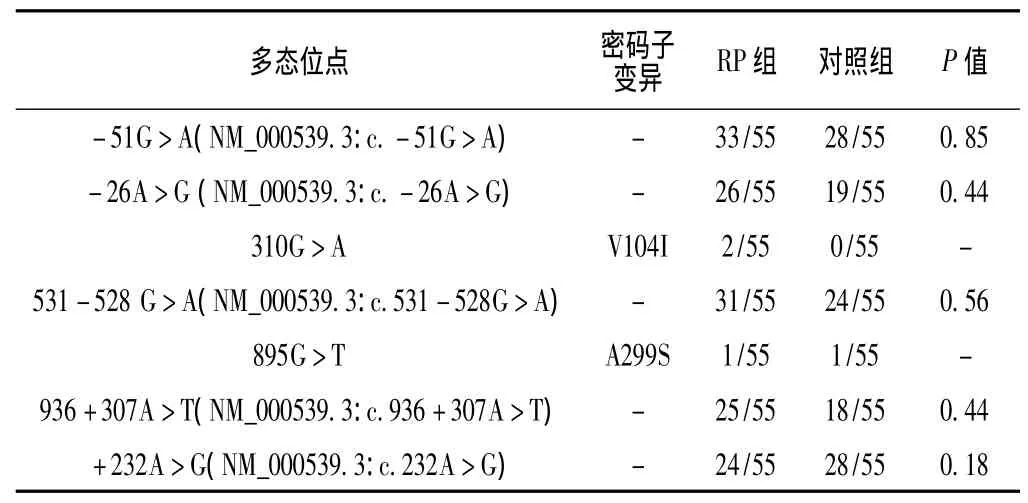
表2 检出的RHO基因点突变Table 2 RHO sequence alterations detected in the study
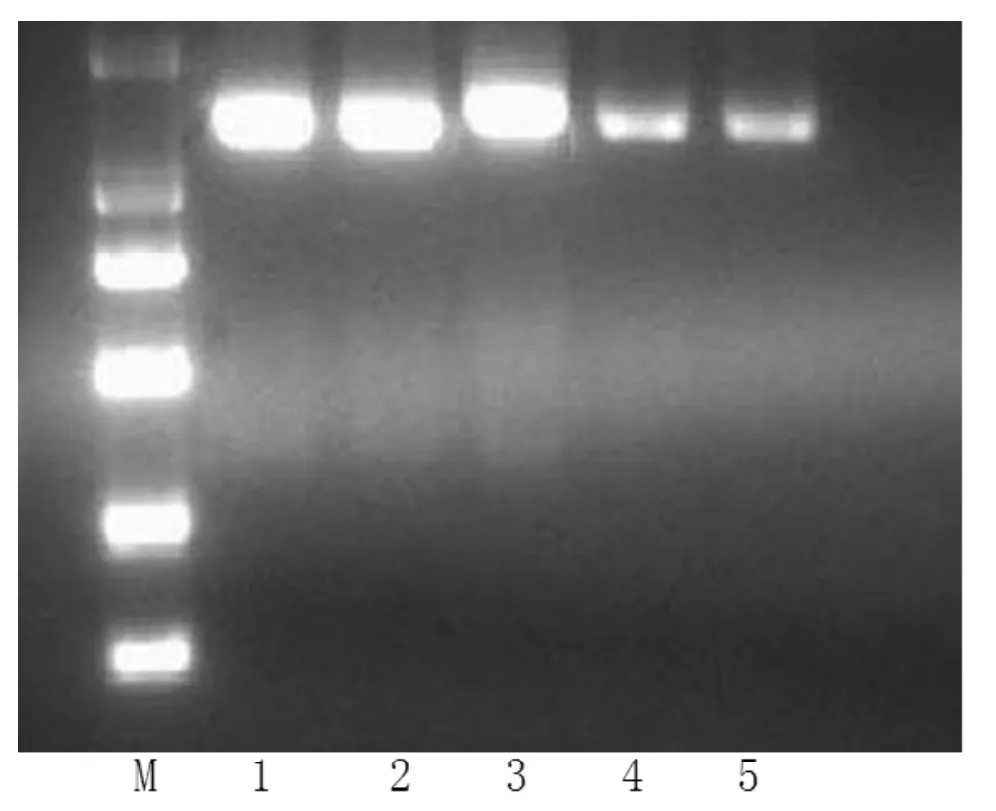
图1 PCR扩增产物2%琼脂糖凝胶电泳图Figure 1 Electrophoretogram of agarose gel amplified by PCR
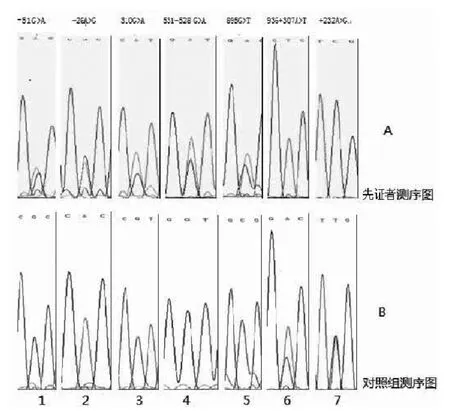
图2 先证者RHO基因7个突变序列和对照者RHO基因7个碱基位点测序结果Figure 2 A graph of7mutation sequence of rhodopsin gene in proband and graph of 7 bases of rhodopsin gene in normal control
3 讨论
RHO基因位于人染色体3q21~24,由4个内含子和5个外显子组成,编码一个由348个氨基酸组成的视蛋白,这是一个在视觉光电转化过程中起作用的G-蛋白偶联受体。RHO基因突变导致结构异常或糖基化改变的视蛋白的产生,该蛋白滞留在感光细胞的内质网中,不能运输到视杆细胞外节盘膜中,或者该视蛋白由于结构异常不能折叠,而不能整合入盘膜或引起盘膜结构不稳定而发生变性,导致RP[9]。自1990年Thaddeus等[10]报道RHO基因突变与某些 ADRP的发病有关以来,目前已发现100余种RHO基因突变,其中约90%为单个碱基置换的点突变,改变了RHO蛋白分子的单个氨基酸而致病,少数是微小缺失或插入突变。
本研究在1例SRP患者和1例ADRP患者中检出错义突变V104I,对照组55名健康成年人未检出此突变。Macke等[11]曾通过家系分析报道该突变为非致病的错义突变。本研究在1例42岁男性ADRP患者及1例对照者RHO基因上检出错义突变A299S。张晓莉等[12]曾报道在1例RP患者及2例正常对照组成员中检出该突变,也系非致病的错义突变。迄今为止国内外已报道的非致病错义突变共3个,除本研究检出的V104I、A299S外,另一个相似突变为P220L[10]。文献报道65种脊椎动物的RHO蛋白中有27种第299位氨基酸为丙氨酸[13],其余均为丝氨酸[14],而丝氨酸也可以看做是丙氨酸的羟基化形式,以此解释A299S突变在RHO基因上属正常的多态变化,但对于其他两种非致病错义突变的机制尚无文献报道。这些错义突变改变了RHO基因的氨基酸序列,引起RHO分子结构和功能异常,虽然已有家系报道为非致病突变,但其机制不明。本研究未检出其他错义突变,我们预测RHO基因在中国华南地区RP患者中的突变率较低。
除错义突变,本研究在RHO非编码区检出多个突变位点,在NCBI-SNP数据库中检索均为已报道的SNP位点,RP组与对照组各SNP位点发生频率均无显著差异。RP遗传学机制复杂,其候选基因确切致病机制及相互作用关系尚不清楚。随着分子生物学的进展,人们认为在内含子与外显子交界处的内含子突变会导致外显子的缺失或内含子不被剪切,发生在内含子中间的变异也多会因为激活了隐性剪切位点而影响了正常剪切从而致病[15]。王丹艺等[16]对151例香港地区汉族 RP先证者和150例对照者进行了RHO基因和RP1基因全编码区和邻近剪切位点的内含子区域序列突变的检测。发现RHO基因非编码区-26G>A突变仅在RP组中出现,其发生率在RP组中显著高于对照组,提示SNP与RP的危险性增加有关。本研究中也检出-26G>A突变,但其在RP组和对照组中的发生率无显著差异。虽然本研究所检测出的非编码区突变与对照组差异不显著,但仍然应该对内含子在RP发病机制中所起的作用引起重视。迄今人们对RP的研究和认识越来越多[17],特别是对内含子及其功能、发生在内含子的基因突变与视网膜色素变性相关性的研究给RP的遗传病因学研究带来了全新的概念。如果发生在内含子的基因突变与RP发病的相互作用被证实,对于RP的发病机制将有更进一步的认识。
本研究检出两种已报道的非致病错义突变,5种非编码区SNP位点,未发现新的错义突变,我们由此预测RHO基因在中国华南地区RP患者中的突变率低于国外报道。本研究尚有不足之处,首先样本量较小,没有发现新的突变位点,其次未对检出错义突变的先证者进行进一步家系研究,以确定检出的错义突变在该先证者家系中是否垂直传递,仅能参考文献报道。进一步研究应尽可能的扩大样本量,争取全基因组检测,对检测错义突变患者进行家系研究,以期发现新的突变位点。
1 Rivolta C,Sharon D,De AngelisMM,et al.Retinitis pigmentosa and allied diseases:numerous diseases,genes,and inheritance patterns hum mol genet[J].Hum Mol Genet,2003,12(5):583 -584.
2 胡诞宁.几种主要遗传病在我国的发病情况与遗传规律 [J].遗传学报,1988,15(3):231-236.
3 Stephen PD,Sara JB,Lori SS.Perspective on genes and mutations causing retinitis pigmentosa [J].Arch Ophthalmol,2007,125(2):151-158.
4 Kajiwara K,Berson EL,Dryla TP.Digenic retinitis pigmentosa due to mutations at the unlinked peripherin/RDS and ROM1 loci[J].Science,1994,264(5165):1604-1608.
5 Zhang XL,Liu M,Meng XH,et al.Mutational analysis of the rhodopsin gene in Chinese ADRP families by conformation sensitive gel electrophoresis[J].Life Sci,2006,78(13):1494 -1498.
6 Kajiwara K,Sandberg MA,Berson EL,et al.A nullmutation in the human peripherin/RDSgene in a family with autosomal dominant retinitis punctata albescens[J].Nat Genet,1993,3(3):208 -212.
7 熊世红,王擎.视网膜色素变性患者视紫红质E341ter基因突变及临床表型分析[J].中华眼科杂志,2002,38(4):224-227.
8 Saga M,Mashima Y,Akeo K,et al.Autosomal dominant retinitis pigmentosa.A mutation in codon 181(Glu- >Lys)of the rhodopsin gene in a Japanese family[J].Ophthalmol Genet,1994,15(2):61 - 67.
9 刘晶,肖林,王薇.常染色体隐性遗传视网膜色素变性家系视紫红质基因突变的检测分析[J].中华眼底病杂志,2004,20(3):145-148.
10 Thaddeus PD,Terri LM,Elias Reichel,et al.A point mutation of the rhodopsin gene in one form of retinitis pigmentosa[J].Nature,1990,25(1):343,364 -366.
11 Macke JP,Davenport CM,Jacobson SG,etal.Identification of novel rhodopsinmutations responsible for retinitis pigmentosa:Implications for the structure and function of rhodopsin[J].Am J Hum Genet,1993,53(1):80-89.
12 张晓莉,府伟灵,彭智培,等.100例视网膜色素变性患者中视紫红质基因突变的筛选与检测[J].中华医学遗传学杂志,2002,19(6):463-466.
13 Deretic D,Schmerl S,Hargrave PA,et al.Regulation of sorting and post-golgi trafficking of rhodopsin by its C-terminal sequence QVS(A)PA[J].Proc Natl Acad Sci USA,1998,95(18):10620 -10625.
14 Horn F,Weare J,Beuker MW,et al.GPCRDB:an informationsystem for G protein 2 coupled receptors[J].Nucleic Acids Res,1998,26(1):275-179.
15 Logsdon JM.The recent origins of spliceosomal introns revisited[J].Curr Opin Genet Dev,1998,8(6):637 -648.
16 王丹艺,范宝剑,陈伟民,等.香港地区汉族人视紫红质基因和视网膜色素变性17基因与视网膜色素变性的双基因关联分析[J].中华医学杂志,2005,85(23):1613 -1618.
17 武晓燕,张彬,苏晓力,等.4+1疗法治疗原发性视网膜色素变性的临床研究[J].中国全科医学,2008,11(5):844.
猜你喜欢
中国造纸(2022年9期)2022-11-25
中国造纸(2022年8期)2022-11-24
广西医科大学学报(2022年5期)2022-06-07
昆明医科大学学报(2022年3期)2022-04-19
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
中南医学科学杂志(2019年6期)2019-12-05
生物工程学报(2019年6期)2019-07-10
环球时报(2019-04-03)2019-04-03
医药前沿(2019年18期)2019-01-04
中国棉花(2018年6期)2018-07-06
