
鼠李糖乳杆菌KF7产单胺氧化酶抑制剂培养条件的优化*
2013-11-21 10:01孟春吴正钧郭本恒韩瑨
食品与发酵工业 2013年10期
孟春,吴正钧,郭本恒,韩瑨
1(光明乳业股份有限公司乳业研究院乳业生物技术国家重点实验室,上海,200436)2(上海海洋大学食品学院,上海,201306)
单胺氧化酶(MAO;EC 1.4.3.4)作为一种主要分布于线粒体外膜的黄素蛋白酶,在人体中能够对内源性和外源性胺类物质氧化脱氨产生醛类和水[1],按照对底物和抑制剂的特异性及分布位置不同,MAO可分为MAO A和MAO B两种亚型。一些报道称抑郁症、帕金森病和阿尔茨海默病等疾病与MAO在体内含量或活性过高有关,能够抑制酶活性的MAO抑制剂(MAOIs)可以作为二线药物来治疗此类精神疾病和神经系统疾病。MAOIs治疗忧郁症具有一定的优势,具有持久并且缓和的特点,不容易从兴奋转变成抑制作用,这与其他的中枢兴奋剂有所不同[2]。
目前天然的MAOIs主要是从不同的植物中提取得到,比如胡椒属的荜拔[3]、乌蔹莓属的乌蔹莓[4]、龙胆属的黄龙胆[5]和薄荷属的水薄荷[6]等一些植物,天然MAOIs可以推动合成药物的发展。对乳酸菌发酵液的MAO抑制作用的报道比较少,Shen[7]等从广西长寿之乡巴马的健康百岁老人粪便中分离出1株双歧杆菌,并研究这株乳酸菌的MRS发酵上清液、菌体和无细胞抽提液的抗氧化活性,体内实验对老龄小鼠进行灌胃,其中一个指标测定大脑中MAO的活性,发现MRS发酵上清液、菌体和无细胞抽提液均能有效降低MAO的活性。本实验室对乳酸菌抑制MAO活性的研究也有一定的成果,第一阶段[8]从健康人体粪便中分离出1株干酪乳酸菌JH23,其MRS发酵上清液对MAO具有较高的抑制率,确定具有MAO抑制作用的是小分子物质。第二阶段[9]从干酪乳杆菌JH23的MRS发酵液中分离纯化出具有MAO抑制作用的物质琥珀酸。由于干酪乳杆菌JH23在脱脂乳中生长缓慢,24h无凝乳现象,脱脂乳发酵液几乎无MAO抑制作用,所以选定几株实验室保藏的24h内能够在脱脂乳中凝乳的乳酸菌,对其发酵液的MAO抑制作用进行比较,最后选择一株具有优势的鼠李糖乳杆菌KF7(Lactobacillus rhamnosus KF7)作为目的菌株。
本实验在单因素实验的基础上,利用Design Expert8.0软件进行响应面实验设计和分析,确定最佳发酵条件,得到具有较高MAO抑制作用的鼠李糖乳杆菌KF7的发酵液。
1 材料和方法
1.1 材料与试剂
鼠李糖乳杆菌KF7(Lactobacillus rhamnosus KF7)等菌种,实验室保藏;单胺氧化酶(MAO),实验室自制;犬脲胺(Kynuramine dihydrobromide),Sigma公司;脱脂乳粉,光明新西兰奶粉厂;KH2PO4,分析纯;K2HPO4:分析纯;其他试剂均为实验室常规分析纯化学试剂。
1.2 仪器与设备
TH-CB-402超净工作台:LABCONCO公司;HVE-50高压蒸汽杀菌锅,日本HIRAYAMA公司;Bug Box厌氧培养箱,RUSKINN公司;GNP-9270隔水式恒温培养箱,上海精宏实验设备有限公司;MDF-U超低温冰箱,日本 SANYO公司;Spectra M5酶标仪,美国Molecular Devices公司;AVANTI J30I高速冷冻离心机,美国BECKMAN COULTER公司。
1.3 实验方法
1.3.1 单胺氧化酶的制备及单胺氧化酶抑制剂活性的测定
单胺氧化酶粗酶的实验室制备根据Stafford[10]等的方法,从Wistar雄性老年大鼠的肝组织匀浆中采用差速离心法在低温条件下分离获得,得到的粗酶液低温保存。
MAO 活性的测定[11-13]:以犬脲胺为底物,在 96孔板中分别加入120 μL磷酸钾缓冲液(0.1 mol/L,pH 7.4 ~7.6),30 μL 待测样品,20 μL 粗酶液和30 μL的犬脲胺底物(1 mmol/L),混匀。反应体系在37℃反应60 min,在波长360 nm下测得初始吸光值(I1)和反应结束后的吸收值(I2),空白对照用水替代待测样品(△I0)。
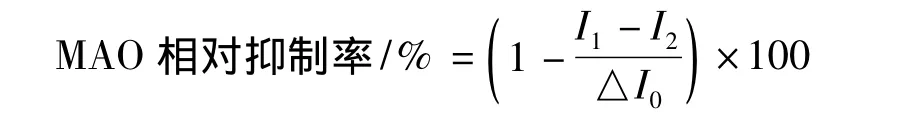
1.3.2 种子液及发酵液制备
将脱脂乳冻存的菌种通过划线法在MRS琼脂上活化,在37℃厌氧培养箱中培养48 h,待菌落长到一定大小后取出,用相同方法再活化一代备用。挑取新鲜培养的菌落接种于5 mL浓度为10%的脱脂乳中,37℃培养24 h左右作为种子液。
将种子液按一定量接种于100 mL浓度为10%的脱脂乳中,在指定温度下培养一定时间取出。将经过发酵后的脱脂乳以9 000 r/min离心20 min,取上清液,作为待测样品,存储于-20℃备用。
1.4 单因素实验
1.4.1 接种量对Lb.rhamnosus KF7产MAO抑制剂的影响
分别按1%、2%、3%、4%、5%的比例将新鲜种子液接种到10%(w/w)脱脂乳中,37℃培养48 h取出,测定发酵液上清对MAO的抑制率。
1.4.2 培养温度对Lb.rhamnosus KF7产MAO抑制剂的影响
将新鲜种子液按4%的比例接种到浓度为10%(w/w)脱脂乳中,分别在 28、31、34、37、40 ℃5 个不同温度培养48 h后取出,测定发酵液上清液对MAO的抑制率。
1.4.3 发酵时间对Lb.rhamnosus KF7产MAO抑制剂的影响
将新鲜种子液按4%的接种量接种到浓度为10%(w/w)的脱脂乳中,在37℃分别培养12、24、48、72、96、120 h后取出,测定发酵液上清对 MAO 的抑制率。
1.4.4 脱脂乳浓度对Lb.rhamnosus KF7产MAO抑制剂的影响
以4%的接种量将新鲜种子液分别接种到4%、6%、8%、10%、12%(w/w)的脱脂乳中,在37℃培养48 h后取出,测定发酵液上清对MAO的抑制率。
1.5 响应面法优化Lb.rhamnosus KF7产MAO抑制剂的发酵条件
采用Design Expert8.0软件进行Box-Behnken响应面实验设计,选取接种量、培养温度和培养时间3个因素,分别用A、B和C进行编码,选取的3个水平用-1、0、1编码,实验设计及水平见表1。
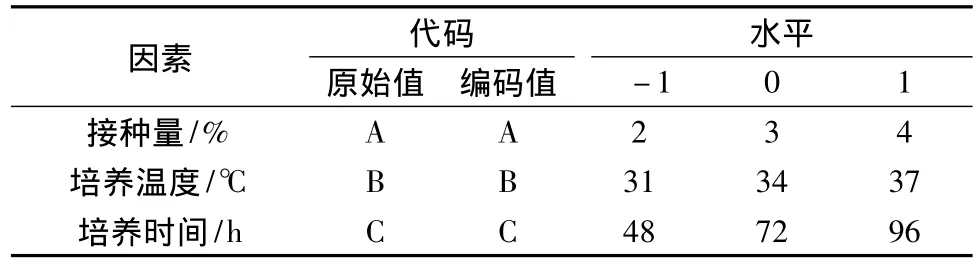
表1 响应面实验设计因素及水平表Table 1 The range of independent variables and their corresponding levels
2 结果与分析
2.1 几株乳酸菌的脱脂乳发酵液对MAO的抑制作用
选取1株干酪乳杆菌(LC2w)、1株鼠李糖乳杆菌(KF7)、1株嗜酸乳杆菌(Lb.acidophilus)、1株乳酸乳球菌(Lc.lactis)、1株瑞士乳杆菌(Lb.helveticus)和1株保加利亚乳杆菌(LB340),在MRS平板上活化后,以4%接种量接种到10%的脱脂乳中,培养48 h,取上清液,分别测定上清液和空白10%脱脂乳的MAO抑制作用,结果如图1。可以看出KF7相对于其他菌株具有更高的MAO抑制作用,抑制率为(59.75±1.61)%,因而选择KF7作为目的菌株进一步研究。
2.2 单因素实验结果
2.2.1 接种量对MAO抑制率的影响
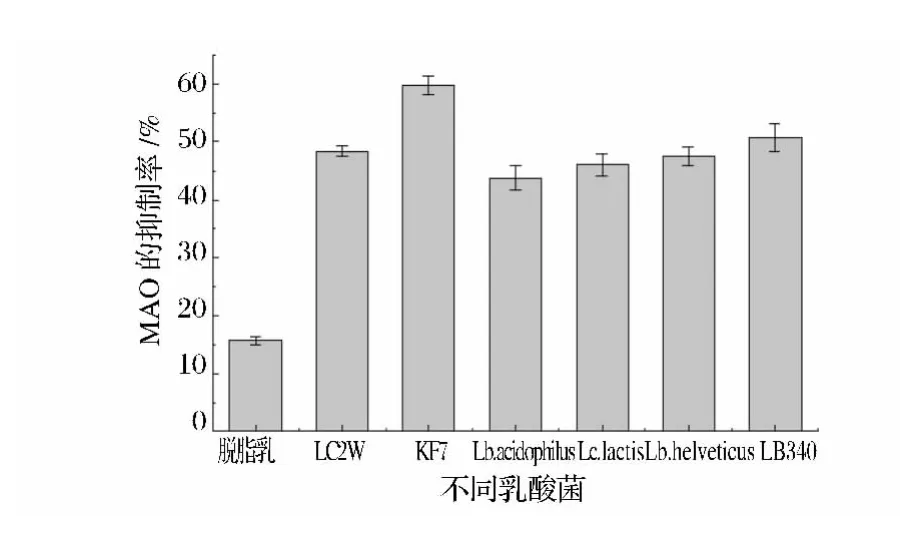
图1 几株乳酸菌发酵脱脂乳上清液和空白脱脂乳对MAO的抑制作用Fig.1 MAO inhibitory effect of fermented skim milk supernatant by the tested strains and blank skim milk
接种量对于微生物的生长和代谢产物的积累具有一定的影响。将Lb.rhamnosus KF7按不同比例接种到10%(w/w)脱脂乳中,在37℃下培养48 h后,发酵脱脂乳对MAO的抑制作用如图2所示。当接种量较小时,发酵脱脂乳对MAO的抑制作用随着接种量的增加而提高,当接种量为3%时,发酵液对MAO的抑制率达到最高;之后,随接种量的继续增加,抑制率出现小幅下降。可能是发酵体系中活菌数量的大量增多,导致营养物质的快速消耗,同时乳酸等不利于菌体生长的有害代谢产物积累过快等因素,影响对MAO具有抑制作用的代谢产物形成和积累。
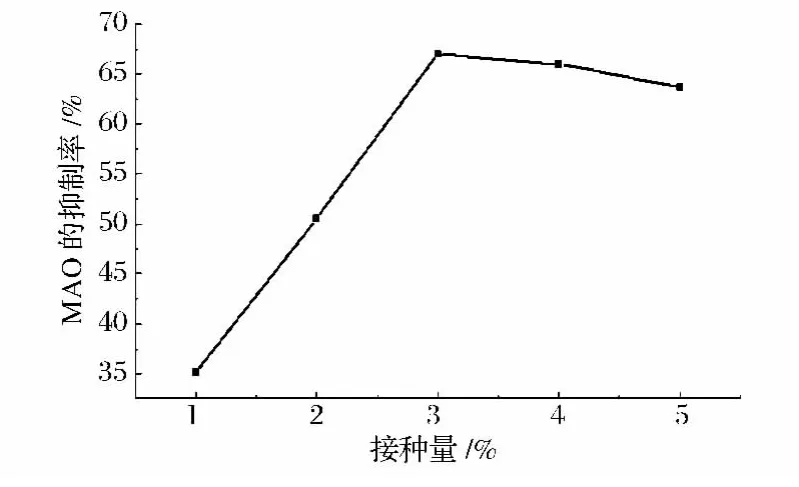
图2 接种量对MAO抑制率的影响Fig.2 Effect of inoculum ration on inhibition of MAO by Lb.rhamnosus KF7 fermented skim milk
2.2.2 培养温度对MAO抑制率的影响
培养温度是影响菌体生长的重要因素之一,适合的温度有利于菌体的生长,菌体代谢活跃,促使代谢产物生成。培养温度对Lb.rhamnosus KF-7在脱脂乳中发酵液抑制MAO活性的影响如图3,温度在28~34℃之间时,随温度的升高,发酵液对MAO的抑制率逐渐升高,在34℃达到最高,当温度再继续升高时,抑制率则会呈现下降趋势。产生这种趋势的可能原因是温度低于或高于34℃时,菌体生长缓慢,因而菌体代谢产物的产生量相对菌体的较适生长温度条件下的量会少。

图3 培养温度对MAO抑制率的影响Fig.3 Effect of culture temperature on inhibition of MAO by Lb.rhamnosus KF7 fermented skim milk
2.2.3 培养时间对MAO抑制率的影响
培养时间对微生物积累代谢产物具有重要的影响,按照4%的接种量在37℃培养,随时间变化发酵液对MAO抑制率的影响。从图4可以得出,在72 h时发酵液对MAO的抑制率较高。随时间增加抑制率逐渐降低,可能是对MAO具有抑制作用的有效物质发生降解。

图4 培养时间对MAO抑制率的影响Fig.4 Effect of culture time on inhibition of MAO by Lb.rhamnosus KF7 fermented skim milk
2.2.4 脱脂乳浓度对MAO抑制率的影响
脱脂乳浓度也是影响菌体生长的一个因素,为菌体生长提供碳源、氮源等生长所必须的营养物质,从图5可以看出脱脂乳浓度为12%时,抑制率最高,与浓度为10%时的抑制率接近,从经济角度考虑,为降低成本,选择10%浓度的脱脂乳。
2.3 响应面分析优化鼠李糖乳杆菌KF7产单胺氧化酶抑制剂的培养条件
2.3.1 响应面实验设计及结果
通过Design Expert8.0软件的Box-Behnken实验设计,选择接种量、培养温度和培养时间作为3个因素,进行3因素3水平设计,见表2。

图5 脱脂乳浓度对MAO抑制率的影响Fig.5 Effect of concentration of skim milk on inhibition of MAO by Lb.rhamnosus KF7 fermented skim milk
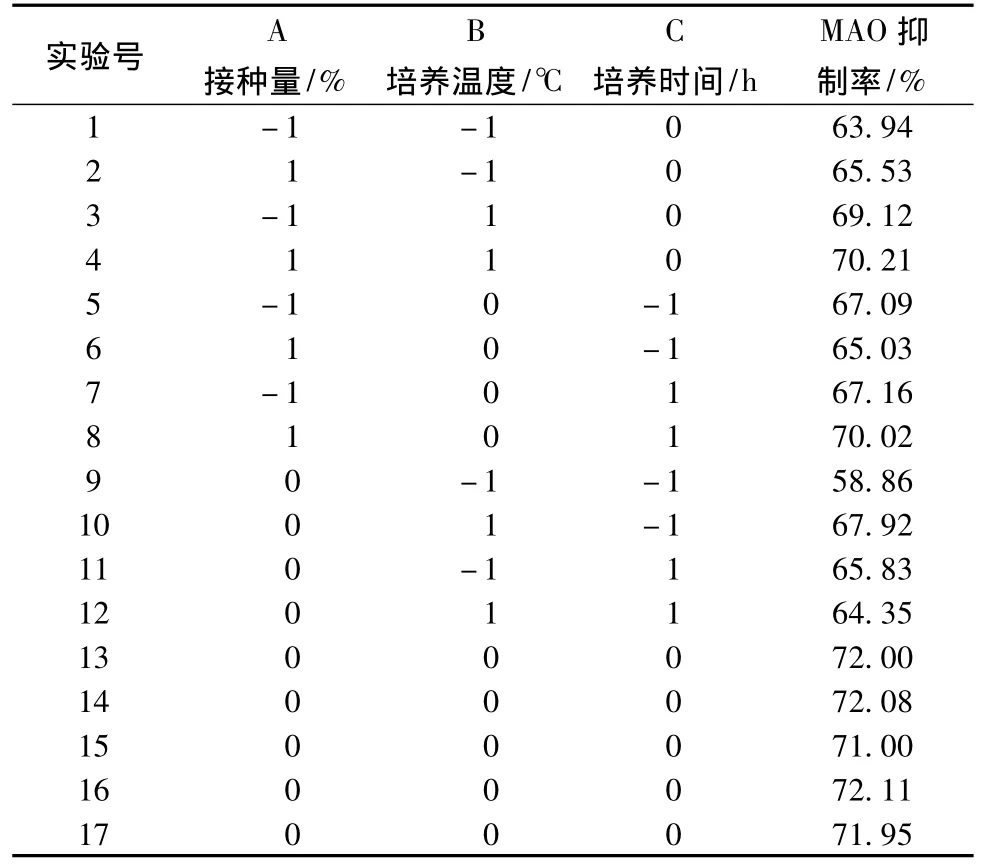
表2 响应面分析实验设计和结果Table 2 Experimental design and result of Response surface analysis
2.3.2 响应面实验方差分析及各因素显著性分析
由表3方差分析可知该模型具有极高的显著性(Pmodel<0.0001),说明本实验选用的二次多项模型是可靠的,其决定系数(R2)=0.982 4,表明发酵液上清对MAO抑制率的实测值与预测值拟合较好。校正决定系数(R2Adj)=0.959 8,表明该模型能较好地解释不同条件下发酵液上清对MAO抑制率的变化。另外,失拟项不显著,因此可以用该模型得到的二次回归方程对实验结果进行分析[14]。
对实验数据进行多项回归拟合,得到发酵上清液的MAO抑制率对编码自变量的二次多项回归方程:
抑制率(%)=71.83+0.44A+2.06B+0.93C-0.13AB+1.23AC-2.39BC-0.90A2-3.73B2-3.61C2
另外,从表3可知,一次项B、C对抑制率分别具有极显著性影响,A影响不显著,交互项AB影响不显著,AC影响显著,BC影响极为显著,A2影响显著,B2、C2影响极显著。说明响应值是受各因素交互影响的,而不是仅仅某一因素的影响。

表3 响应面回归方程的方差分析Table 3 The analysis of variance for the polynomial model
2.3.3 响应面分析
响应面分析得到的三维曲面图和等高线图可以直观地反映各因素对响应值的影响及各参数之间的相互作用,从响应曲面的最高点和等高线可以看出在所选的范围内存在极值。如图6所示,随接种量和培养温度的增大,抑制率是逐渐增大的,达到某一个值后,随2个自变量的增大,抑制率则呈现下降趋势。图7和图8显示,抑制率随2个自变量的变化同图6具有相同的趋势。
通过分析得到最佳条件的理论值为,接种量3.29%,培养温度34.71℃,培养时间74.4 h,此培养条件得到MAO抑制率的理论值为72.18%。考虑到实际条件,验证实验的最佳优化条件:接种量3.3%,培养温度34.7℃,培养时间74 h。实验重复3次,取平均值得MAO抑制率的验证值为(71.98±0.16)%,与理论值接近(相对误差为0.3%),结果表明采用响应面分析法得到的发酵培养条件基本准确,具有实用价值。

图6 接种量和培养温度交互影响MAO抑制率的等高线图及其曲面图Fig.6 Contour plot and respective response surface plot of MAO inhibition rate of Lb.rhamnosus KF7 fermented skim milk affected by inoculum ratio and culture temperature
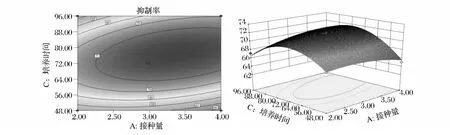
图7 接种量和培养时间交互影响MAO抑制率的等高线图及其曲面图Fig.7 Contour plot and respective response surface plot of MAO inhibition rate of Lb.rhamnosus KF7 fermented skim milk affected by inoculum ratio and incubation time

图8 培养温度和培养时间交互影响MAO抑制率的等高线图及其曲面图Fig.8 Contour plot and respective response surface plot of MAO inhibition rate of Lb.rhamnosus KF7 fermented skim milk affected by culture temperature and time
3 结论与讨论
单因素实验表明随接种量、培养温度和培养时间的增大,发酵脱脂乳上清的MAO抑制率的趋势是先增大后减小,通过单因素实验找到每一个因素的较优点,从而在响应面实验中能够选取合适的水平。
本文通过Box-Behnken中心组合实验对发酵条件进行优化,优化过程具有较高的准确性和精确性。通过对模型进行方差分析,显示该模型能够用于鼠李糖乳杆菌KF7产单胺氧化酶抑制剂的发酵条件的预测和分析。最终得到的最佳发酵条件为:接种量3.3%,培养温度34.7℃,培养时间74 h,验证得到的MAO抑制率为71.98%。
目前具有MAO抑制作用的天然产物主要是从植物中得到,采用微生物发酵方法制备MAO抑制剂的研究相对较少。人体多种精神和神经系统疾病都与MAO的失调有关,MAO抑制剂则起到缓解和治疗作用,MAO抑制剂的研究和开发是具有医学价值的,从天然产物到化学合成,得到了不同种类具有价值的MAO抑制剂。针对这一现状,在前期的研究中,我们通过体外实验发现鼠李糖乳杆菌KF7发酵脱脂乳的产物具有较高的MAO抑制作用。为了进一步的研究,首先通过对Lb.rhamnosus KF7产单胺氧化酶抑制剂的发酵条件进行优化,以提高发酵脱脂乳对MAO的抑制作用,从而为接下来的工作提供依据。
[1]Malorni W,Giammarioli A M,Matarrese P,et al.Protection against apoptosis by monoamine oxidase A inhibitors[J].FEBS Letters,1998,426(1):155-159.
[2]金国章,鄒岡,胥彬.治疗精神病药物单胺氧化酶抑制剂的进展概况[J].药学学报,1963,10(1):51-63.
[3]Lee S A,Hwang J S,Han X H,et al.Methylpiperate derivatives from Piper longum and their inhibition of monoamine oxidase [J].Archives of Pharmacal Research,2008,31(6):679-683.
[4]Han X H,Hong S S,Hwang J S,et al.Monoamine oxidase inhibitory components from Cayratia japonica[J].Archives of Pharmacal Research,2007,30(1):13-17.
[5]Haraguchi H,Tanaka Y,Kabbash A,et al.Monoamine oxidase inhibitors from Gentiana lutea[J].Phytochemistry,2004,65(15):2 255-2 260.
[6]Olsen H T,Stafford G I,Van S J,et al.Isolation of the MAO-inhibitor naringenin from Mentha aquatica L [J].Journal of Ethnopharmacology,2008,117(3):500-502.
[7]Shen Q,Shang N,Li P L.In vitro and in vivo antioxidant activity of Bifidobacterium animalis 01 isolated from centenarians[J].Current microbiology,2011,62(4):1 097-1 103.
[8]王豪,郭本恒,王荫榆,等.具有抑制单胺氧化酶活性的干酪乳杆菌JH-23生长特性及生理功能研究[J].食品工业科技,2010,31(11):168-171.
[9]何一薇,郭本恒,陈卫,等.干酪乳杆菌JH23代谢产物对单胺氧化酶的抑制作用[J].天然产物研究与开发,2012,24(7):882-887.
[10]Stafford G I,Pedersen P D,Jager A K,et al.Monoamine oxidase inhibition by southern African traditional medicinal plants[J].South African Journal of Botany,2007,73(3):384-390.
[11]Krajl M.A rapid microfluorimetric determination of monoamine oxidase[J].Biochemical Pharmacology,1965,14(11):1 684.
[12]Schmidt K,LI Z,Schubert B,et al.Screening of entomopathogenic Deuteromycetes for activities on targets involved in degenerative diseases of the central nervous system[J].Journal of Ethnopharmacology,2003,89(2):251-260.
[13]Ashby J R C R,Carr L A,Cook C L,et al.Alteration of platelet serotonergic mechanisms and monoamine oxidase activity in premenstrual syndrome[J].Biological Psychiatry,1988,24(2):225-233.
[14]胡明明,张国文,何力.响应面法优化超声提取花生壳多酚[J].南昌大学学报:理科版,2011,35(3):241-246.
猜你喜欢
当代水产(2022年1期)2022-04-26
昆明医科大学学报(2020年12期)2021-01-26
乳业科学与技术(2019年6期)2019-12-22
中国乳品工业(2017年9期)2017-10-12
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年3期)2016-11-15
中国粮油学报(2016年5期)2016-01-23
食品工业科技(2014年9期)2014-03-11
茶叶通讯(2014年2期)2014-02-27
食品科学(2013年15期)2013-03-11
