
使用多因素易患性阈值模型评估双相情感障碍的遗传度缺失
2014-05-25 00:32李康许瑞环张洪德王前
遗传 2014年9期
李康,许瑞环,张洪德,王前
1. 南方医科大学南方医院检验医学中心,广州 510515;
2. 深圳市龙岗中心医院中心实验室,深圳 518116;
3. 深圳市龙岗中心医院检验科,深圳 518116
使用多因素易患性阈值模型评估双相情感障碍的遗传度缺失
李康1,2,许瑞环3,张洪德2,王前1
1. 南方医科大学南方医院检验医学中心,广州 510515;
2. 深圳市龙岗中心医院中心实验室,深圳 518116;
3. 深圳市龙岗中心医院检验科,深圳 518116
为了评估双向情感障碍的遗传度缺失,文章通过查询美国国家人类基因组研究所(National Human Genome Research Institute,NHGRI)的gwascatalog目录,检索出所有已发现的双相情感障碍易感变异,使用多因素易患性阈值模型计算每个易感变异对双相情感障碍遗传度的解释度。将所有易感变异遗传度解释度求和得到双相情感障碍已知易感变异对遗传度的总解释度,使用此总解释度评估双相情感障碍的遗传度缺失。结果显示,已知双相情感障碍易感变异对双相情感障碍遗传度的合计解释度为38.34%,尚有61.66%的遗传度无法被已有易感变异解释,属于遗传度缺失。双相情感障碍 38.34%的遗传度解释度较早前国外同类研究大幅度提高,表明随着新的双相情感障碍易感变异被不断发现,双相情感障碍遗传度缺失得到大幅度减小。但双相情感障碍遗传度缺失依然存在且数目较大的事实也表明双相情感障碍尚存在许多未知的分子遗传学机制有待进一步阐明。
双相情感障碍;遗传度;遗传度缺失;易患性阈值模型
双相情感障碍是一种典型的多基因遗传病,疾病的形成受多个易感变异控制,并且受环境因素的影响。Edvardsen等[1]研究发现双相情感障碍的遗传度为77.00%,表明遗传因素在双相情感障碍的发病机制中发挥着主要作用,也预示了双相情感障碍存在大量易感变异。
为阐明双相情感障碍的发病机制,国内外学者进行了大量的研究,取得了丰硕的成果。特别是全基因组关联分析(Genome-wide association study, GWAS)技术在双相情感障碍分子机制中的应用,大大地促进了双相情感障碍易感变异的研究进程,发现了大量的双相情感障碍易感变异。但随着研究的深入,已发现的大量双相情感障碍易感变异并不能完全解释双相情感障碍的高遗传度,即当前发现的双相情感障碍易感变异对双相情感障碍发病的贡献远远小于其遗传度,这种现象被称为“遗传度缺失(Missing heritability)”[2]。
遗传度缺失现象普遍存在于双相情感障碍、精神分裂症、糖尿病、乳腺癌等多种复杂疾病。So[3]等报道了基于多因素易患性阈值模型评估已知易感突变对复杂疾病遗传度的解释度的方法,并对多种常见复杂疾病易感变异对遗传度的解释度进行了统计计算。结果显示已发现易感变异对双相情感障碍遗传度的解释度仅2.77%,有高达97.23%的双相情感障碍遗传度无法被已知易感变异解释,属于遗传度缺失。目前国内还未见关于双相情感障碍的遗传度缺失的研究。本研究使用So等[3]的研究方法计算双相情感障碍的易患性解释变异(Variance in liability explained, Vg),使用Vg与双相情感障碍遗传度的比值评估双相情感障碍所有已知易感变异对双相情感障碍遗传度的解释度,并对双相情感障碍的遗传度缺失进行评估。
1 对象和方法
1.1 研究对象
双相情感障碍的易感变异信息主要来源于美国国家人类基因组研究所(National Human Genome Research Institute, NHGRI)编制的已发表GWAS研究目录gwascatalog(http://www.genome.gov/gwastudies) (截止2013年9月30日),Hindorff等[4]对此目录做了详细的介绍。目录中与双相情感障碍相关的易感变异共有377个,其中324个易感变异因为未提供风险等位基因频率(Risk allele frequency)和比值比(OR)等数据而无法使用本方法计算其对双相情感障碍遗传度的解释度,未纳入本研究。本研究主要以剩余的53个双相情感障碍易感变异为研究对象,计算其对双相情感障碍遗传度的解释度,进而对双相情感障碍的遗传度缺失进行评估(表1)。
1.2 方法
假设每个易感变异均为一个风险等位基因A和一个保护等位基因 a组成的双等位基因。对于同一个人群,易感变异3种基因型AA、Aa、aa具有不同的平均易患性,相同的易患性阈值(图1)。等位基因A、a的基因频率分别表示为PA、Pa。基因型AA、Aa、aa的基因型频率分别表示为PAA、PAa、Paa。基因型AA、Aa、aa的外显率分别表示为fAA、fAa、faa。RR1代表基因型 Aa相对于基因型 aa的相对风险率(Relative risks, RR),RR2代表基因型AA相对于基因型aa的相对风险率。由于基因型Aa含有一个风险等位基因,而基因型AA含有两个风险等位基因,所以RR2=RR12。本研究使用易感基因的OR值作为其RR1的估计值。基因型AA、Aa、aa的易患性平均值分别表示为 μAA、μAa、μaa。T表示疾病的易患性阈值,K表示疾病总发病率。双相情感障碍的总发病率为0.021[5]。
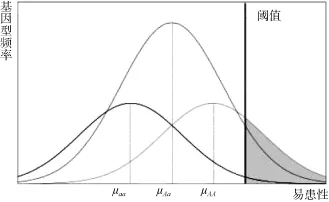
图1 易患性阈值模型示意图μAA、μAa、μaa分别为基因型AA、Aa、aa的易患性平均值,阴影部分表示双相情感障碍患者。
首先,根据哈迪温伯格定律,用PA推算出Pa、PAA、PAa、Paa。
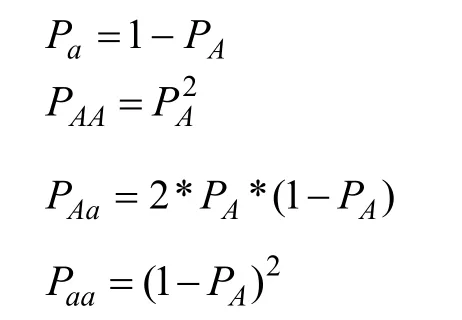
总患病率K与基因型频率,基因型外显率,相对风险比的关系可用下列公式表达:
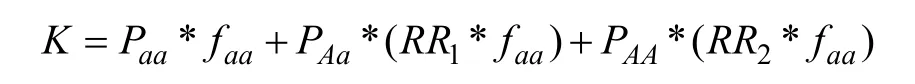
由此可推导出:
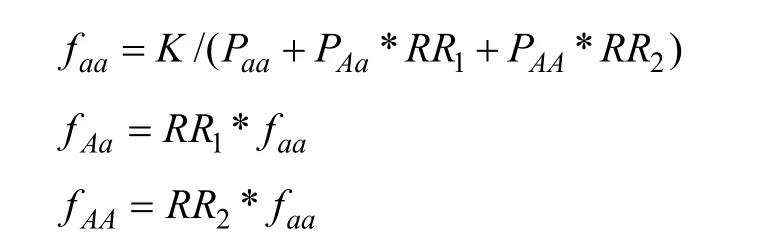
假设 3种基因型的残余方差均为 1,则基因型
特异性易患性均值与基因型外显率存在如下关系:
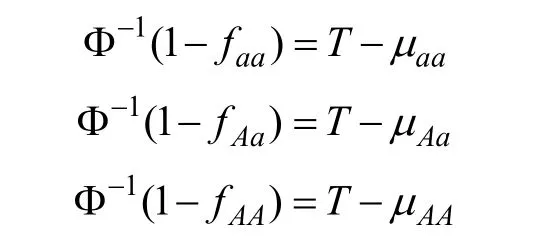
其中 Φ-1为返回正态分布百分位数的函数。可以假设μaa数值为0,这不会影响对Vg的计算。则T、μAa、μAA可计算如下:
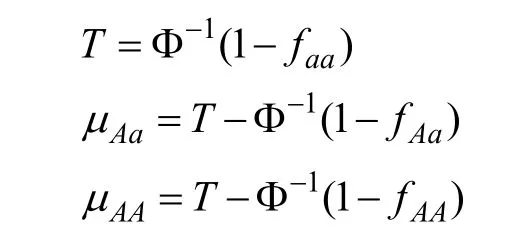
所有人群总的易患性平均值μall计算公式如下:
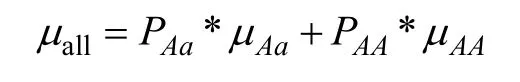
Vg计算公式如下:
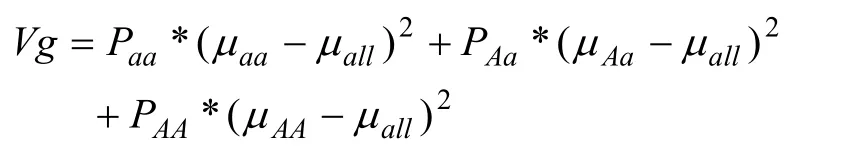
为方便与遗传度进行比较,将Vg按照如下公式进行变换,变换后的Vg表示为实际Vg(actual Vg)。
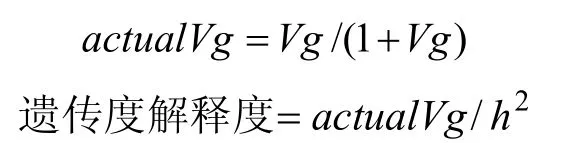
h2代表遗传度,双相情感障碍的遗传度为77.00%[1]。
So等[3]提供了此方法的程序软件,可免费下载使用(http://sites.google.com/site/ honcheongso/ software/varexp)。
2 结果与分析
运用多因素易患性阈值模型计算出 53个双相情感障碍易感变异对于双相情感障碍遗传度的解释度,结果见表 1。解释度低于 1.00%的易感变异有45个,对双相情感障碍遗传度的合计解释度为10.90%,其中解释度最低的是位于20p13的rs3761218,对双相情感障碍的遗传度解释度为 0.01%。解释度高于1.00%的易感基因有8个,对双相情感障碍遗传度的合计解释度为 27.43%,其中解释度最高的是位于1p31.3的 rs41350144,其对双相情感障碍的遗传度解释度为6.94%。53个易感变异对双相情感障碍遗传度的合计解释度为38.34%。双相情感障碍遗传度中尚存在 61.66%的部分无法被已知易感变异解释,这部分无法被已知易感变异解释的遗传度即可理解为双相情感障碍的遗传度缺失。
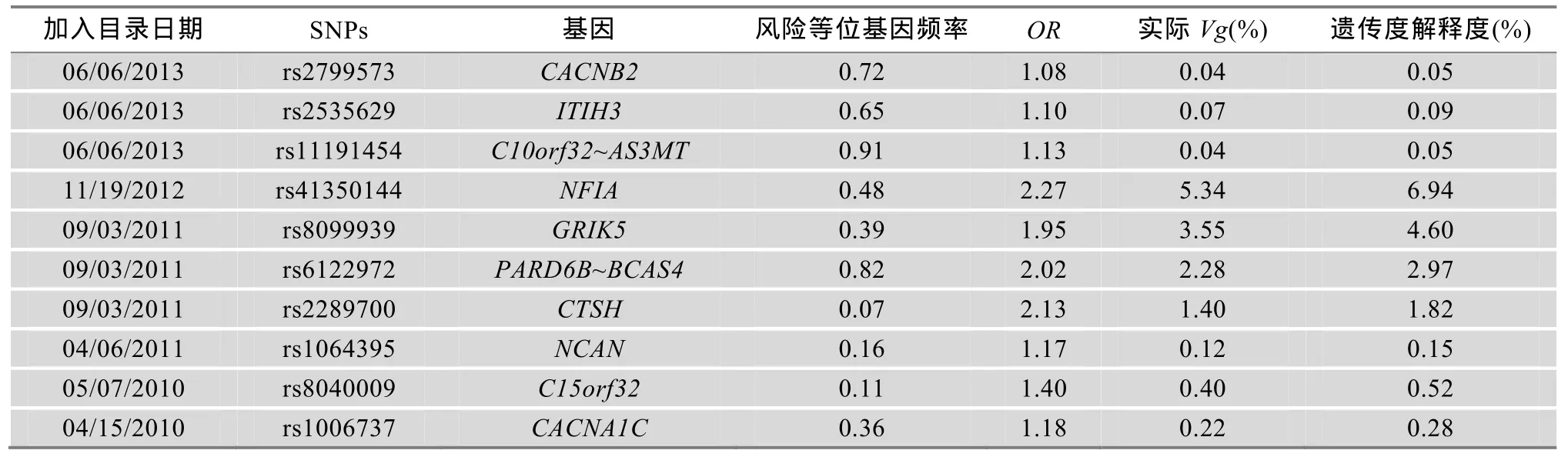
表1 已知双相情感障碍易感变异对双相情感障碍遗传度的解释度
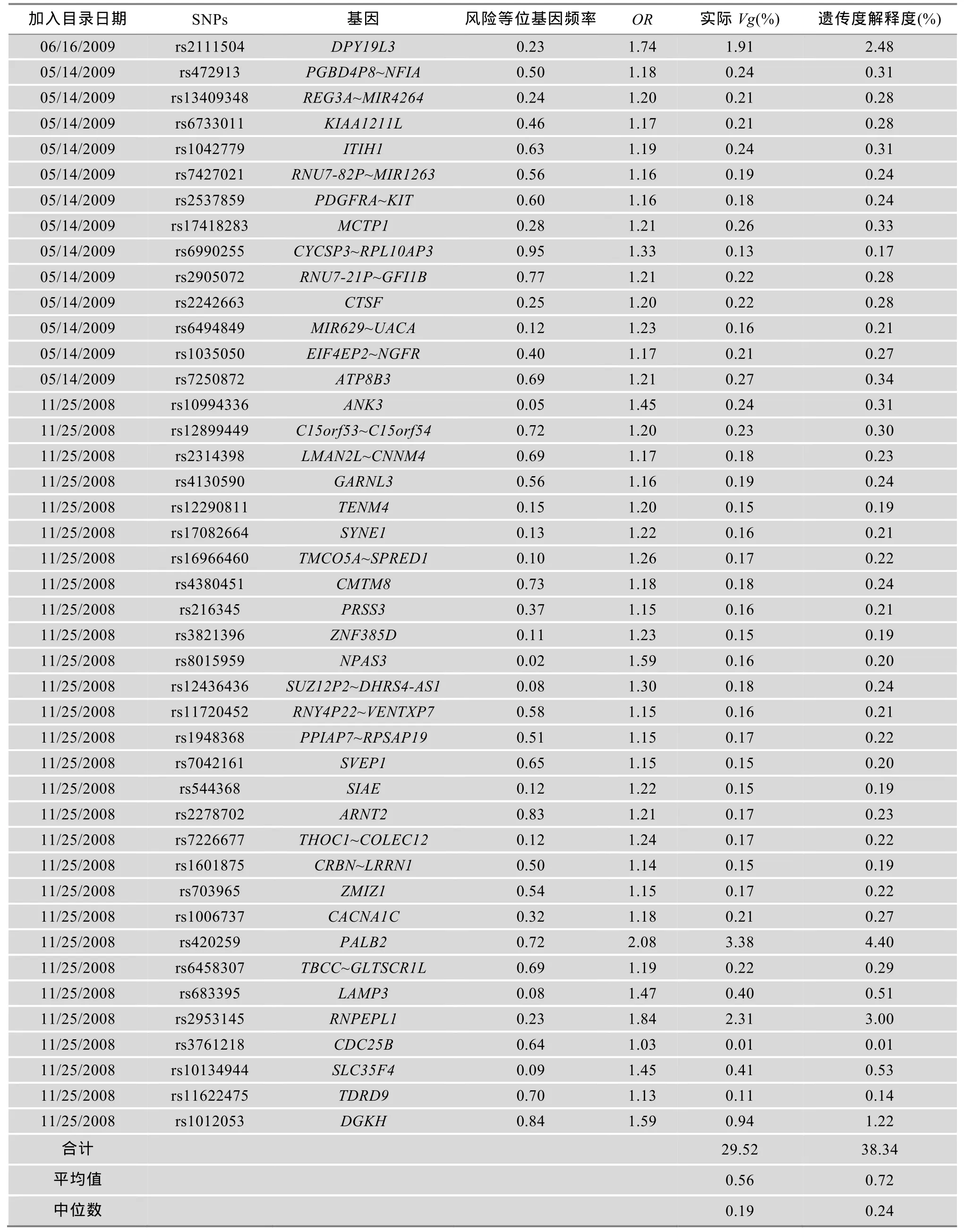
续表
3 讨 论
双相情感障碍是典型的多基因遗传病。随着GWAS技术越来越广泛的运用于双相情感障碍的研究,目前有超过300个双相情感障碍易感变异被发现。然而双相情感障碍众多易感变异的发现对于双相情感障碍发病机制的阐明及诊断治疗并未产生显著地推动作用。对比单基因疾病易感变异对于疾病发病机制及诊断治疗的巨大推动价值,双相情感障碍显示出更加复杂的遗传学发病机制。研究显示已发现的双相情感障碍易感变异无法完全解释疾病的遗传度,从而出现遗传度缺失现象。遗传度缺失现象普遍存在于多基因遗传病中,已成为多基因遗传病易感变异研究的主要瓶颈和热点研究领域。
So等[3]对包含双相情感障碍在内的十种常见多基因遗传病的已发现易感变异对遗传度的解释度进行定量计算,结果显示解释度最高的是前列腺癌,其解释度为 31.16%,解释度最低的精神分裂症仅0.39%,双向情感障碍的解释度为2.77%。本研究显示,已发现双相情感障碍易感变异对遗传度的解释度为 38.34%,显著高于先前报道。分析其中原因,主要有以下几点:首先,大量新近发现的双相情感障碍易感变异的纳入提高了本研究的总遗传度解释度。So等[3]的研究参考了NHGRI截至2010年1月31日的gwascatalog。当时gwascatalog中收录了47个双相情感障碍易感变异,排除 4个未提供风险等位基因频率(Risk allele frequency)和(或)比值比(OR)的易感变异后剩余43个易感变异。剩余的43个易感变异再按照 p值小于 7.20×10-8的标准进行筛选,排除39个p值大于 7.20×10-8的易感变异,剩余4个易感变异。So等[3]使用gwascatalog目录中这4个剩余的易感变异加上1个gwascatalog外的易感变异合计 5个双相情感障碍易感变异用于评估双相情感障碍遗传度解释度。2010年后,全球范围内开展了大量的新的双相情感障碍GWAS研究,有大量新的双相情感障碍易感变异被发现。2010年2月1日至今,gwascatalog新录入的双相情感障碍易感变异多达 330个,本研究选择了其中 10个易感变异。这10个易感变异的合计遗传度解释度达到17.48%,其中Lee等[6]发现的rs41350144和Yosifova等[7]发现的 rs8099939对双相情感障碍遗传度的解释度分别为6.94%和4.60%,是目前发现的对双相情感障碍遗传度解释度最高的两个易感变异。这些新发现的易感变异的纳入大幅提高了易感变异对双相情感障碍的总遗传度解释度。其次,对基因组显著性阈值的要求适当放宽,使得更多的易感变异纳入本研究。So等[3]的研究为了保证结果准确度,仅纳入p值小于7.20×10-8的易感变异,从而使gwascatalog中39个双相情感障碍易感变异未纳入研究,仅有 4个易感变异被纳入研究。考虑到GWAS研究者在发现易感变异研究中,均已根据自身研究特点,选择了合适的基因组显著性阈值,为了全面的反映所有已发现双相情感障碍易感变异对其遗传度的解释度,本文未采用7.20×10-8的显著性阈值对易感变异进行筛选,将所有53个易感变异全部纳入研究,从而使得双相情感障碍的总遗传度解释度大幅度提高。
本研究对已知易感变异对双相情感障碍遗传度的解释度的计算也存在一定的局限性。第一,本研究主要选择的双相情感障碍易感变异主要来源于GWAS研究,而GWAS自身的一些局限性,例如较难识别低最小基因频率突变(0.5%<MAF<5%)和稀有突变(MAF<0.5%),对拷贝数变异(CNVs)和中性拷贝变异(copy neutral variation)等结构性变异较难识别也从一定程度上制约了本研究。第二,对于双相情感障碍易感变异,本研究主要参考美国 NHGRI编制的已发表GWAS研究目录gwascatalog。可能存在一些双相情感障碍易感变异由于未收录入gwascatalog而未被纳入本研究,从而使计算所得的双相情感障碍遗传度解释度偏低。第三,本研究使用的计算方法要求易感变异必须有相应的风险等位基因频率和 OR方能计算其遗传度解释度。Gwascatalog目录中与双相情感障碍相关的总计377个易感变异中,仅53个同时提供了风险等位基因频率和OR,324个易感变异因为未提供风险等位基因频率和(或)OR等数据而未纳入本研究。这一方法学局限性使得超过 80%的已发现双相情感障碍易感变异对双相情感障碍遗传度的解释度无法得到估算。第四,本研究使用比值比作为相对危险率的估计值,但当疾病患病率较高时,相对危险率将小于比值比,这样使用相对危险率计算所得的总遗传度解释度将小于使用比值比计算所得的遗传度解释度。
38.34 %的遗传度解释度说明双相情感障碍依然有大部分遗传度无法得到解释,存在较大的遗传度缺失。随着全球范围内双相情感障碍GWAS研究的不断开展,新的双相情感障碍易感变异的不断发现将使得双相情感障碍遗传度解释度进一步得到提升。但基于GWAS如上所述的局限性,仅仅依赖GWAS研究无法完全解决双相情感障碍的遗传度缺失。Manolio等[8]建议的复杂疾病遗传度缺失研究策略包括:对极端表型患者进行目标区域或全基因组测序;使用扩大的基因组变异参考品以增大GWAS研究的覆盖度;挖掘现有GWAS研究,研究基因结构变异与疾病的关联及基因间相互作用的证据;通过扩大单个研究和 Meta分析来增加多种复杂性疾病研究的样本量,包括对非欧裔人群的研究。相信随着研究样本量的不断增大,基因-基因、基因-环境间相互作用的不断阐明,对稀有变异和结构性变异与复杂疾病相关性的深入研究,复杂疾病的遗传度缺失问题将得到进一步解决。
Lee等[9]建立了名为基因组复杂性状分析(Genome-wide complex trait analysis, GCTA)的基于线性混合模型定量计算复杂疾病遗传度缺失的统计学方法,目前已被迅速应用于包括帕金森病[10]在内的众多复杂疾病遗传度缺失研究,并显示出良好的研究价值。我们计划在下一步的研究中使用GCTA方法对双相情感障碍遗传度缺失做进一步研究。
[1] Edvardsen J, Torgersen S, Røysanb E, Lygren S, Skre I, Onstad S, Øien PA. Heritability of bipolar spectrum disorders. Unity or heterogeneity? J Affect Disord, 2008, 106(3): 229-240.
[2] Maher B. Personal genomes: The case of the missing heritability. Nature, 2008, 456(7218): 18-21.
[3] So HC, Gui AHS, Cherny SS, Sham PC. Evaluating the heritability explained by known susceptibility variants: a survey of ten complex diseases. Genet Epidemiol, 2011, 35(5): 310-317.
[4] Hindorff LA, Sethupathy P, Junkins HA, Ramos EM, Mehta JP, Collins FS, Manolio TA. Potential etiologic and functional implications of genome-wide association loci for human diseases and traits. Proc Natl Acad Sci USA, 2009, 106(23): 9362-9367.
[5] Merikangas KR, Akiskal HS, Angst J, Greenberg PE, Hirschfeld RMA, Petukhova M, Kessler RC. Lifetime and 12-month prevalence of bipolar spectrum disorder in the National Comorbidity Survey replication. Arch Gen Psychiatry, 2007, 64(5): 543-552.
[6] Lee HJ, Woo HG, Greenwood TA, Kripke DF, Kelsoe JR. A genome-wide association study of seasonal pattern mania identifies NF1A as a possible susceptibility gene for bipolar disorder. J Affect Disord, 2013, 145(2): 200-207.
[7] Yosifova A, Mushiroda T, Kubo M, Takahashi A, Kamatani Y, Kamatani N, Stoianov D, Vazharova R, Karachanak S, Zaharieva I, Dimova I, Hadjidekova S, Milanova V, Madjirova N, Gerdjikov I, Tolev T, Poryazova N, O'Donovan MC, Owen MJ, Kirov G, Toncheva D, Nakamura Y. Genome-wide association study on bipolar disorder in the Bulgarian population. Genes Brain Behav, 2011, 10(7): 789-797.
[8] Manolio TA, Collins FS, Cox NJ, Goldstein DB, Hindorff LA, Hunter DJ, McCarthy MI, Ramos EM, Cardon LR, Chakravarti A, Cho JH, Guttmacher AE, Kong A, Kruglyak L, Mardis E, Rotimi CN, Slatkin M, Valle D, Whittemore AS, Boehnke M, Clark AG, Eichler EE, Gibson G, Haines JL, Mackay TFC, McCarroll SA, Visscher PM. Finding the missing heritability of complex diseases. Nature, 2009, 461(7265): 747-753.
[9] Lee SH, Wray NR, Goddard ME, Visscher PM. Estimating missing heritability for disease from genome-wide association studies. Am J Hum Genet, 2011, 88(3): 294-305.
[10] Keller MF, Saad M, Bras J, Bettella F, Nicolaou N, Simón-Sánchez J, Mittag F, Büchel F, Sharma M, Gibbs JR, Schulte C, Moskvina V, Durr A, Holmans P, Kilarski LL, Guerreiro R, Hernandez DG, Brice A, Ylikotila P, Stefánsson H, Majamaa K, Morris HR, Williams N, Gasser T, Heutink P, Wood NW, Hardy J, Martinez M, Singleton AB, Nalls MA; International Parkinson's Disease Genomics Consortium (IPDGC); Wellcome Trust Case Control Consortium 2 (WTCCC2). Using genome-wide complex trait analysis to quantify ‘missing heritability’ in Parkinson’s disease. Hum Mol Genet, 2012, 21(22): 4996-5009.
(责任编委: 周钢桥)
Evaluating the missing heritability of bipolar disorder using the multifactorial liability threshold model
Kang Li1,2, Ruihuan Xu3, Hongde Zhang2, Qian Wang1
1. Laboratory Medical Center, Nanfang Hospital, Southern Medical University, Guangzhou 510515, China;
2. Central Laboratory, Longgang District Central Hospital, Shenzhen 518116, China;
3. Clinical Laboratory, Longgang District Central Hospital, Shenzhen 518116, China
In order to evaluate the missing heritability of bipolar disorder, we queried the GWAS catalog of National Human Genome Research Institute, retrieve all the susceptible gene variation of bipolar disorder, and calculate the heritability explanation degree of each susceptibility variant using the multifactorial liability threshold model. The total heritability explanation degree of bipolar disorder was obtained through summing up the heritability explanation degree of each susceptibility variant. Then, we evaluated the missing heritability of bipolar disorder based on the total heritability explanation degree.The results showed that the total heritability explanation degree of bipolar disorder explained by known susceptible variants was 38.34%, and the other 61.66% of heritability can’t be explained by known susceptibility variants, which belong to the missing heritability of bipolar disorder. The total heritability explanation degree of bipolar disorder in this study was significantly increased compared to earlier similar studies abroad. With constant discovery of new bipolar disorder susceptibility variants, the missing heritability of bipolar disorder has been greatly reduced, but the missing heritability of bipolar disorder still exists and occupies a large part of the bipolar disorder heritability, indicating that the molecular genetic mechanisms of bipolar disorder need to be further clarified.
bipolar disorder; heritability; missing heritability; liability threshold model
2014-01-31;
2014-07-07
广东省科技厅课题项目(编号:2011B061200003)资助
李康,硕士,主管技师,研究方向:复杂疾病的基因组和蛋白质组学。E-mail:kangerem@163.com
王前,博士,教授,研究方向:复杂疾病的快速诊断。E-mail:wangqian@smu.edu.cn
10.3724/SP.J.1005.2014.0897
时间: 2014-7-30 11:30:44
URL: http://www.cnki.net/kcms/detail/11.1913.R.20140730.1130.003.html
猜你喜欢
数学物理学报(2022年5期)2022-10-09
山东冶金(2022年2期)2022-08-08
现代临床医学(2021年4期)2021-07-31
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
中成药(2018年12期)2018-12-29
百科知识(2015年18期)2015-09-10
现代检验医学杂志(2015年6期)2015-02-06
作物研究(2014年6期)2014-03-01
中国糖料(2013年1期)2013-01-22
