
金黄色葡萄球菌肠毒素B激活TACE诱导人鼻黏膜上皮细胞分泌黏蛋白
2016-12-24 17:35,,,
中南医学科学杂志 2016年4期
,,,
(南华大学附属第一医院耳鼻喉科,湖南 衡阳 421001)
·基础医学·
金黄色葡萄球菌肠毒素B激活TACE诱导人鼻黏膜上皮细胞分泌黏蛋白
刘珺,廖剑绚*,唐洪波,周芝芳
(南华大学附属第一医院耳鼻喉科,湖南 衡阳 421001)
目的观察金黄色葡萄球菌肠毒素B(SEB)对人鼻黏膜上皮细胞活性氧(ROS)和肿瘤坏死因子转化酶(TACE)活性的影响,并探讨其在介导黏蛋白MUC5AC分泌中的作用。方法体外培养人鼻黏膜上皮细胞,用不同浓度的SEB孵育细胞0~24 h,采用荧光共振能量转移法检测TACE的酶活性,荧光探针H2DCFDA检测鼻黏膜上皮细胞中活性氧(ROS)的含量,ELISA检测培养上清中MUC5AC的分泌水平。同时采用TACE siRNA干扰或ROS抑制剂NAC处理细胞,观察其在介导MUC5AC分泌中的作用。结果100 ng/mL SEB作用鼻黏膜上皮细胞15 min后即可上调TACE的酶活性,30~60 min后达到峰值,并持续至2 h;SEB也能增加鼻黏膜上皮细胞内ROS的含量;采用NAPDH氧化酶抑制剂DPI处理后,ROS水平明显减少;ROS抑制剂NAC处理鼻黏膜上皮细胞后,可明显降低TACE的酶活性;30~100 ng/mL SEB能显著诱导鼻黏膜上皮细胞分泌MUC5AC,采用TACE siRNA干扰其表达或抑制剂处理后,可抑制MUC5AC分泌。结论SEB激活TACE诱导人鼻黏膜上皮细胞分泌MUC5AC。
金黄色葡萄球菌肠毒素B;肿瘤坏死因子α转化酶;活性氧;黏蛋白
慢性鼻—鼻窦炎(chronic rhinosinusitis,CRS)是一种多因素所致的鼻黏膜纤毛功能障碍,是耳鼻喉科最常见的多发病之一。其病因与发病机制复杂,病原微生物感染、鼻内解剖结构异常,遗传因素、以及环境因素均是CRS的重要诱因[1,2]。CRS作为一种炎症相关疾病,病原体感染在其发病中的作用并不完全明确。CRS显著的病理生理特征是鼻黏膜黏液分泌增加,纤毛清除功能减弱,后者可引起黏液淤积并继发性引起细菌感染,由此引起的炎症反应又可加重黏液—纤毛清除系统的损伤,形成恶性循环[3]。人体呼吸道黏液的主要成分是黏蛋白,它主要由鼻黏膜上皮杯状细胞和杯状细胞分泌,目前已知的黏蛋白基因近20余种,其中以MUC5AC最重要。生理条件下,MUC5AC在维持气道湿化、协助上皮细胞功能等方面发挥重要作用,因此是呼吸系统抵御外界刺激的非特异性屏障。但在某些病理条件下,黏蛋白的过度分泌可使CRS黏膜局部防御功能减弱和纤毛清除功能降低,导致局部反复感染[4-5]。研究表明,CRS患者鼻腔内金黄色葡萄球菌、表皮葡萄球菌的检出率较高[6-7],其中金黄色葡萄球菌肠毒素B(Staphylococcus aureus enterotoxin B,SEB)可能是CRS发病的重要病原相关分子模式[8]。但SEB究竟通过何种机制参与黏蛋白分泌目前尚未明了,本研究旨在初步探讨SEB诱导MUC5AC分泌的分子机制。
1 材料与方法
1.1试剂SEB购自Toxin Technology (Sarasota,FL),纯度>95%。胎牛血清、DMEM/Ham F12培养基购自Hyclone。二亚苯基碘(Diphenyleneiodonium,DPI)、N-乙酰-半胱氨酸(N-acetyl-cysteine,NAC)胰蛋白酶、2′,7′-二氯二氢荧光黄二乙酸酯(2′,7′-dichlorodihydrofluorescein diacetate,H2DCFDA)以及TAPI购自Sigma-Adrich。MUC5AC ELISA检测试剂盒购自武汉华美生物工程有限公司。肿瘤坏死因子转化酶(tumor necrosis factor converting enzyme,TACE)活性检测试剂盒购自Anaspec。TACE siRNA由广州ribobio公司合成。
1.2细胞培养本研究所用的鼻黏膜上皮细胞来自下鼻甲黏膜组织,为南华大学附属第一医院耳鼻喉科行鼻中隔手术患者的标本。开展研究前获得了患者的知情同意且研究方法符合本院医学伦理委员会标准。所有患者无变应性鼻炎、哮喘病史,且近期无糖皮质激素、抗组胺类药物及阿司匹林耐受不良史。即手术中获取的下鼻甲标本于含有100 IU/mL青霉素和100 μg/mL链霉素的磷酸盐缓冲液中,无菌条件下反复冲洗。随后置于5倍体积的胰蛋白酶中(0.25%),4 ℃消化16~18 h。用吸管吹打2~3 min,收集脱落的上皮细胞,加入终浓度为2.5%的胎牛血清终止消化。1 000 rpm离心5min收集并采用完全培养基(DMEM/Ham F12培养基中含10%胎牛血清、100 IU/mL青霉素、100 μg/mL链霉素)漂洗细胞2次。将细胞接种于培养板中,根据参考文献提供的方法去除鼻黏膜上皮细胞中的成纤维细胞和红细胞[9]。细胞于37 ℃、5% CO2条件下培养。待细胞80%以上融合后,加入1,10,100 ng/mL SEB处理不同时间后用于下一步研究。
1.3 ROS测定鼻黏膜上皮细胞处理完毕后,终浓度5 μmol/L的H2DCFDA染液,37 ℃避光孵育30 min,每3~5 min轻微振荡1次,使探针和细胞充分作用。孵育结束后用无菌PBS漂洗涤细胞3次以去除未进入细胞内的探针。设置荧光酶标仪激发波长485 nm,发射波长530 nm,测定其荧光强度。
1.4 TACE活性检测采用荧光共振能量转移原理间接测定TACE的酶活性,按照AnaSpec公司提供的试剂盒的操作步骤进行。该试剂盒含有底物QXLTM520/ 5-FAM,该底物能被TACE特异性降解,从而使QXLTM520无法淬灭荧光分子5-FAM。因此当样品中若含有TACE后,它通过降解该底物而产生5-FAM,其荧光强度与TACE的活性成正比。在荧光酶标仪(Synergy HT,Bio-Tec)上设置激发波长490 nm,发射波长520 nm,测定各组荧光强度,结果以相对活性表示,计算公式:处理组活性/对照组活性。
1.5转染与RNA干扰将生长状态旺盛的鼻黏膜上皮细胞接种于6孔板中(含1.5 mL无血清培养基)培养24 h。按Lipofectamine 2000提供的操作步骤配制好A液和B液。其中A液将10 μL siRNA用无血清培养基溶解(本文所用的TACE siRNA序列分别为:正义链5′-GGUUUUAAAGGCUAUGGAAtt-3′;反义链5′-UUCCAUAGCCUUUAAAACCtg-3′),终体积为250 μL。B液将5 μL Lipofectamine 2000用250 μL无血清培养基稀释。充分混匀后室温静置20 min;随后将上述A、B混合液加至细胞培养液中,使TACE siRNA终浓度约为100 nmol/L。4 h后弃上清,更换为完全培养液,随后用于下一步研究。
1.6 ELISA检测MUC5AC分泌当生长于6孔板中的鼻黏膜上皮细胞长满80%~90%视野时,弃培养基,并加入不同浓度的SEB作用相应时间。处理结束后,采用连续冻融细胞方式裂解细胞,获取裂解产物,1 000 rpm离心5 min后,将上清用于MUC5AC检测。根据试剂盒提供的操作步骤,在包被有MUC5AC抗体的微孔板中加入100 μL待测液,37 ℃孵育2 h。经洗涤后,按照试剂盒的步骤分别加入鼠抗人MUC5AC抗体以HRP标记的羊抗鼠二抗孵育。显色后,测定450 nm处的吸光度值,并根据标准曲线计算MUC5AC的浓度。
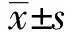
2 结 果
2.1 SEB增强鼻黏膜上皮细胞TACE的酶活性如图1A所示,采用不同浓度SEB处理鼻黏膜上皮细胞30 min后,随着浓度的递增,TACE的酶活性逐渐增高,当SEB浓度为100 ng/mL时,TACE的酶活性增加了2.8倍。此外,SEB对TACE酶活性的影响也呈一定的时间依赖性,即100 ng/mL SEB作用15 min后即可明显增强TACE的酶活性,30~60 min后达到峰值,并持续至2 h(图1B)
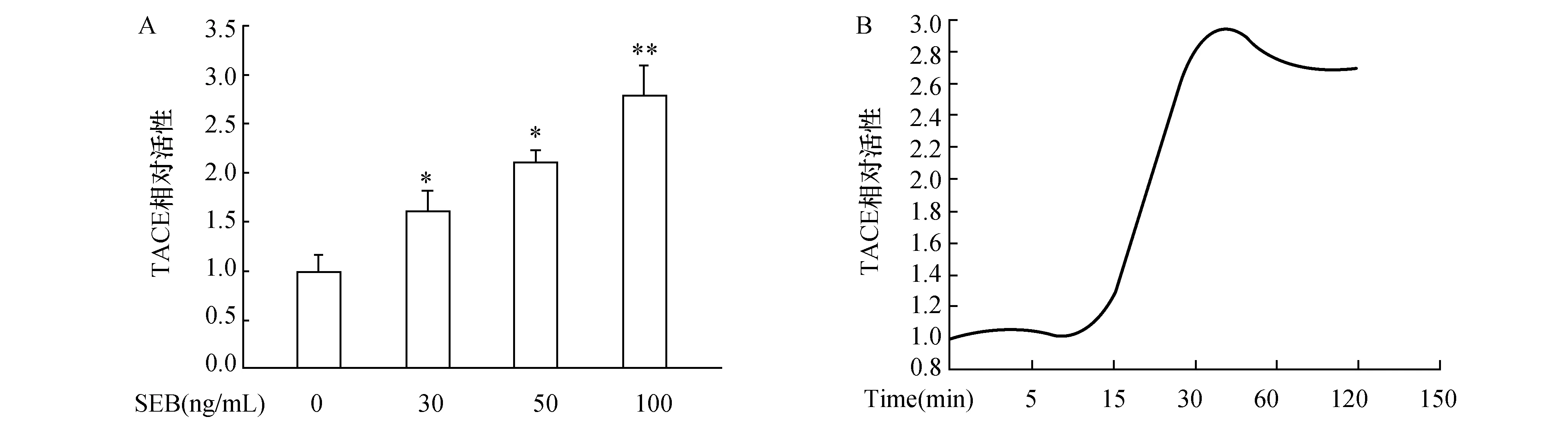
图1 SEB对鼻黏膜上皮细胞TACE酶活性的影响 A:不同浓度SEB对TACE活性的影响.与0ng/mL SEB组相比,*:P<0.05,**:P<0.01;B:100ng/mL SEB作用不同时间对TACE活性的影响
2.2 SEB经ROS增强TACE酶活性SEB处理鼻黏膜上皮细胞30 min后,即可明显促进细胞内产生ROS。而给予NADPH氧化酶抑制剂DPI(10 μmol/L)预处理细胞后,ROS含量明显减少(图2A)。此外,采用不同浓度ROS抑制剂NAC预处后,TACE酶活性随之降低(图2A)。
2.3 SEB促进鼻黏膜上皮细胞分泌MUC5AC ELISA结果显示,采用不同浓度SEB处理24 h后,随着浓度的递增,MUC5AC分泌水平逐渐增多,当SEB浓度为100 ng/mL时,MUC5AC高达(312±31)ng/mL(图3A)。此外,MUC5AC的分泌也呈一定的时间依赖性,即100 ng/mL SEB作用8 h后,MUC5AC显著增多,16~24 h后达到峰值,并持续至48 h(图3B)
2.4 TACE参与SEB诱导鼻黏膜上皮细胞分泌MUC5AC 鼻黏膜上皮细胞经而给予1.0、5.0和10.0 μmol/L TACE抑制剂预TAPI处理细胞后,随着浓度的增加,MUC5AC分泌逐渐减少(图3A)。采用siRNA干扰TACE表达后, MUC5AC分泌明显降低(图3B、C)。
3 讨 论
黏液高分泌是慢性鼻及鼻窦炎的主要病理学特征之一,黏蛋白在维持气道湿化、协助上皮细胞功能等方面发挥重要作用,因此是呼吸系统抵御外界刺激的非特异性屏障。黏液的异常分泌是导致鼻腔黏液纤毛清除系统功能损害的重要因素,继而极易引发细菌感染。细菌感染是CRS黏液高分泌的常见原因,其机制涉及多重信号通路的调控,包括丝裂原活化蛋白激酶(MAPKs)、核转录因子NF-κB和AP-1等。不同的刺激因素所激活的信号通路有所不同。目前认为,MUC5AC分泌主要受TACE和EGFR调控[10,11]。TACE是金属水解蛋白(adamalysin)家族的膜结合型整合素样金属蛋白酶(a disintegrin and metallpprotase,ADAM),其功能主要是催化膜结合型TNF-α前体水解为具有生物活性TNF-α。除此之外,它也能促进TGF-α等前体分子,从而释放出可溶性TGF-α,后者随后结合至EGFR上,通过配体-受体反应激活EGFR而诱导MUC5AC表达。本研究也证实,鼻黏膜上皮细胞在静息状态下,TACE酶活性较低,而给予100 ng/mL SEB作用15 min后即可诱导其激活,并持续至2 h以上,而通过RNA干扰或其抑制剂处理后,SEB诱导MUC5AC的水平显著减少,这表明TACE参与了SEB作用后MUC5AC的表达。为了进一步探讨SEB诱导MUC5AC的调控机制,我们对TACE的上游通路进行了探讨。TACE是ADAM家族成员,其结构中含有一个抑制性前结构域,1个催化活性区,1个解聚素结构域,1个富含脯氨酸的铰链区以及一个跨膜结构域和一个胞浆结构域。一般认为,TACE主要有两种激活机制,一是以ROS为代表,通过氧化前结构域中的半胱氨酸后,可暴露出TACE的催化位点。第二中机制以佛波脂为代表,它通过激活蛋白激酶C,引起TACE胞外结构域构象改变,从而暴露出催化结构域[12,13]。本研究发现,SEB处理后30 min后,即可明显促进细胞内产生ROS,采用ROS抑制剂处理后,TACE活性明显降低。而ROS的产生又受NADPH氧化酶的调控,我们采用其抑制剂DPI处理后,发现胞内ROS的含量也明显减少。以上结果表明TACE的激活依赖于上游的NADPH氧化酶和ROS的产生。
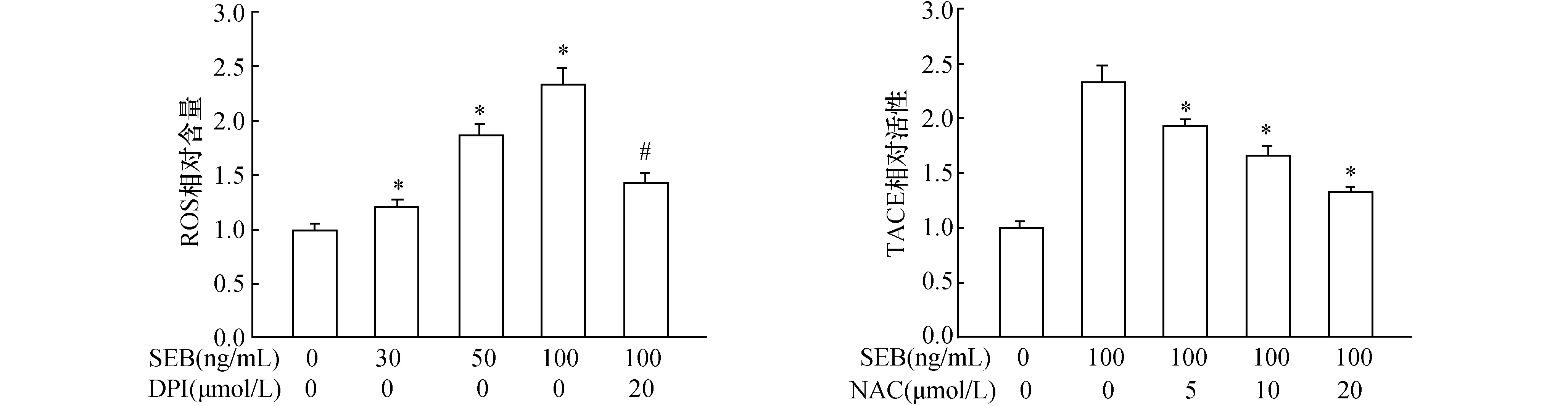
图2 SEB诱导经ROS增强鼻黏膜上皮细胞中TACE酶活性 A:SEB对鼻黏膜上皮细胞中ROS产生的影响.与0ng/mL SEB相比,*:P<0.05;与100ng/mL SEB相比,#:P<0.05;B:ROS抑制剂对TACE酶活性的影响.与100 ng/mL SEB单独处理组相比,*:P<0.05
图3 SEB诱导鼻黏膜上皮细胞分泌MUC5AC 与0 ng/mL SEB组相比,*:P<0.05,**:P<0.01
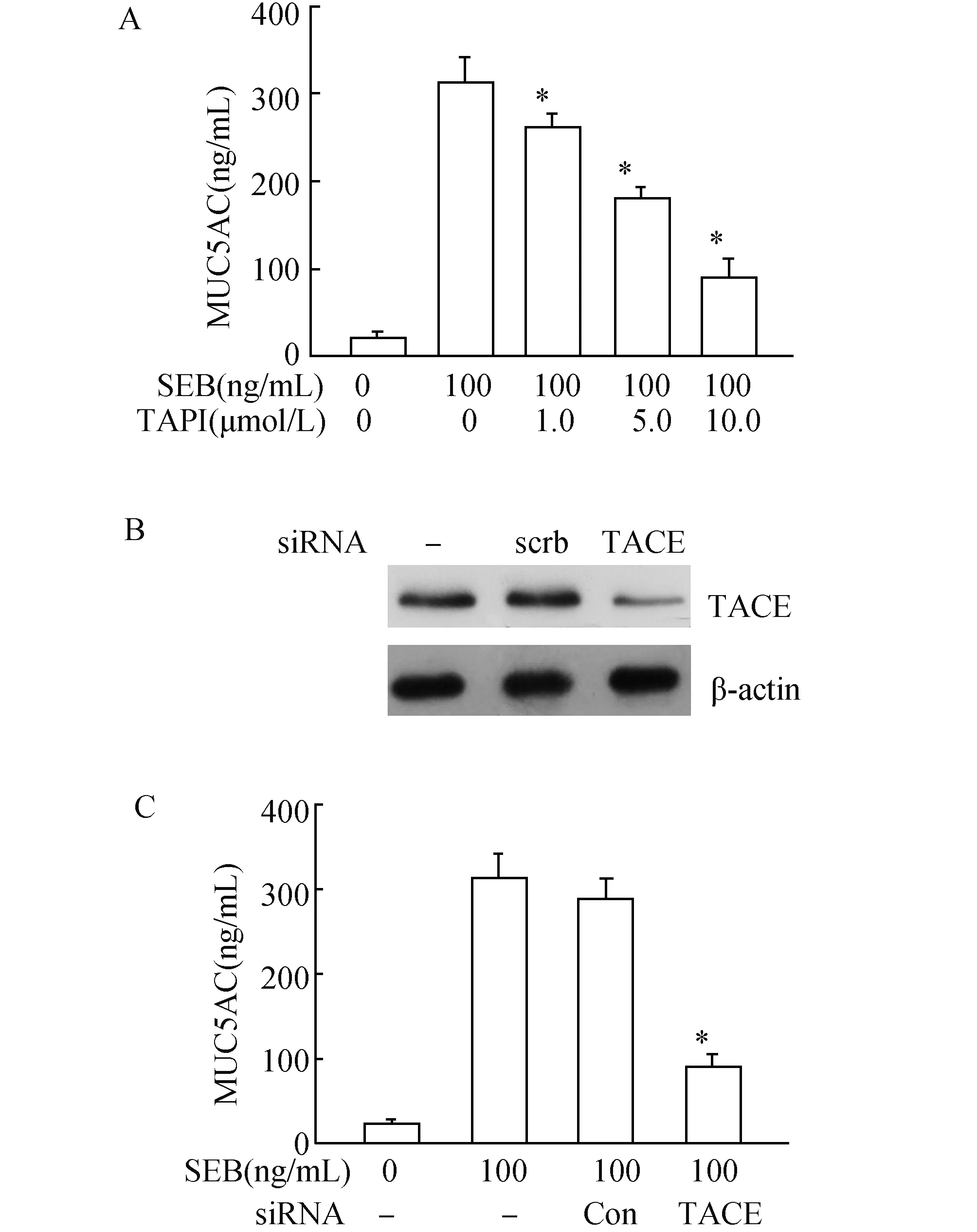
图4 TACE诱导SEB诱导鼻黏膜上皮细胞分泌MUC5ACA:与100 ng/mL+0 μmol/L TAPI相比,*:P<0.05;B:TACE siRNA干扰效果;C:与对照siRNA组(Con)相比,*:P<0.05
总之,本研究证实SEB可鼻黏膜上皮细胞分泌MUC5AC。这表明金黄色葡萄球菌感染后,可能通过诱导MUC5AC过度分泌而加重COPD病情。若内炎症状态持续存在,黏液合成和分泌增多造成气流受限,同时过多积聚的黏液又可破坏气道的非特异性防御功能,有利于病原菌定植,从而加重感染。此外,本研究也证实SEB通过ROS/TACE途径诱导MUC5AC分泌,这是一条新的途径,但不是唯一途径。因为MUC5AC上也存在NF-κB、Sp1等核转录因子的结合位点,这些分析可能最终也参与了MUC5AC的分泌。在随后的研究当中,我们将对EGFR的其他分子开展更深入探讨,从而进一步明确CRS的发病机制。
[1] Stevens WW,Lee RJ,Schleimer RP,et al.Chronic rhinosinusitis pathogenesis[J].J Allergy Clin Immunol,2015,136(6):1442-1453.
[2] 蔡建良,陈浩,达瑞,等.尘螨变应原疫苗治疗变应性鼻炎的临床动态观察[J].中南医学科学杂志,2014,42(6):585-588.
[3] Bachert C,Zhang L,Gevaert P.Current and future treatment options for adult chronic rhinosinusitis:Focus on nasal polyposis[J].J Allergy Clin Immunol,2015,136(6):1431-1440.
[4] Seshadri S,Lu X,Purkey MR,et al.Increased expression of the epithelial anion transporter pendrin/SLC26A4 in nasal polyps of patients with chronic rhinosinusitis[J].J Allergy Clin Immunol,2015,136(6):1548-1558.e7.
[5] Kim DH,Chu HS,Lee JY,et al.Up-regulation of MUC5AC and MUC5B mucin genes in chronic rhinosinusitis[J].Arch Otolaryngol Head Neck Surg,2004,130(6):747-752.
[6] Brook I.Bacteriology of chronic sinusitis and acute exacerbation of chronic sinusitis[J].Arch Otolaryngol Head Neck Surg,2006,132(10):1099-1101.
[7] Busaba NY,Siegel NS,Salman SD.Microbiology of chronic ethmoid sinusitis:is this a bacterial disease[J].Am J Otolaryngol,2004,25(6):379-384.
[8] Kim YM,Jin J,Choi JA,et al.Staphylococcus aureus enterotoxin B-induced endoplasmic reticulum stress response is associated with chronic rhinosinusitis with nasal polyposis[J].Clin Biochem,2014,47(1-2):96-103.
[9] 鄂征.组织培养技术及其在医学研究中的应用[M].北京:中国协和医科大学出版社,2004:151.
[10] Liu Z,Tian F,Feng X,et al.LPS increases MUC5AC by TACE/TGF-alpha/EGFR pathway in human intrahepatic biliary epithelial cell[J].Biomed Res Int,2013,2013:165715.
[11] Barbier D,Garcia-Verdugo I,Pothlichet J,et al.Influenza A induces the major secreted airway mucin MUC5AC in a protease-EGFR-extracellular regulated kinase-Sp1-dependent pathway[J].Am J Respir Cell Mol Biol,2012,47(2):149-157.
[12] Bass R,Edwards DR.ADAMs and protein disulfide isomerase:the key to regulated cell-surface protein ectodomain shedding[J].Biochem J,2010,428(3):e3-5.
[13] Zhang Z,Oliver P,Lancaster JR Jr,et al.Reactive oxygen species mediate tumor necrosis factor alpha-converting,enzyme-dependent ectodomain shedding induced by phorbol myristate acetate[J].FASEB J,2001,15(2):303-305.
[14] Schamberger AC,Staab-Weijnitz CA,Mise-Racek N,et al.Cigarette smoke alters primary human bronchial epithelial cell differentiation at the air-liquid interface[J].Sci Rep,2015,5:8163.
[15] Lee SU,Sung MH,Ryu HW,et al.Verproside inhibits TNF-alpha-induced MUC5AC expression through suppression of the TNF-alpha/NF-kappaB pathway in human airway epithelial cells[J].Cytokine,2016,77:168-175.
[16] Deshmukh HS,Shaver C,Case LM,et al.Acrolein-activated matrix metalloproteinase 9 contributes to persistent mucin production[J].Am J Respir Cell Mol Biol,2008,38(4):446-454.
StaphylococcusAureusEnterotoxinBInducesHumanNasalEpithelialCellsSecretionofMUC5ACviaActivationofTACE
LIU Jun,LIAO Jianxuan,TANG Hongbo,et al
(Depart.Otorhinolaryngology,TheFirstaffiliatedhospitalofUniversityofSouthChina,Hengyang421001,China)
ObjectiveTo observe the effect of staphylococcus aureus enterotoxin B (SEB) on reactive oxygen species (ROS) production and tumor necrosis factor converting enzyme (TACE) activity in human nasal epithelial cells,and explore its role in MUC5AC secretion.MethodsHuman nasal epithelial cells were cultured in vitro,and then were stimulated with different concentrations of SEB for 0~24 h.The activity of TACE was measured by fluorescence resonance energy transfer method,SEB-induced production of ROS was detected by using the fluorescent probe H2DCFDA.To investivate the role of TACE and ROS in MUC5AC secretion,nasal epithelial cells was transfected by TACE siRNA or the ROS inhibitor NAC,and Secretion of MUC5AC in the culture supernatant was detected by ELISA.ResultThe enzymic activity of TACE was up-regulated followed by treatment of 100ng/mL of SEB for 15min,peaked at 30~60min and lasted for 2 h.In addition,SEB could induce ROS accumulation in human nasal epithelial cells,and this ability could be abrogated by DPI,an inhibitor of NADPH oxidase.Moreover,treatment of the ROS inhibitor NAC significantly decreased the enzymic activity of TACE.Furthermore,secretion of MUC5AC were abrogated when the cells were transfected the TACE siRNA or treated by its inhibitor TAPI.ConclusionsSEB induces MUC5AC secretion via ROS and TACE in human nasal epithelial cells.
staphylococcal aureus enterotoxin B; tumor necrosis factor-α converting enzyme; reactive oxygen species; MUC5AC
10.15972/j.cnki.43-1509/r.2016.04.011
2016-03-06;
2016-05-19
湖南省科技计划项目(2013SK3114).
*通讯作者,E-mail:278691897@qq.com.
R378.1
A
秦旭平)
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
河北农业大学学报(2022年2期)2022-04-26
食品安全导刊(2021年20期)2021-08-30
实用肿瘤学杂志(2020年4期)2020-12-08
中华养生保健(2020年3期)2020-11-16
科研成果与传播(2019年3期)2019-09-10
中国临床医学影像杂志(2019年5期)2019-08-27
中国临床医学影像杂志(2019年2期)2019-04-25
中成药(2017年8期)2017-11-22
中国医学影像学杂志(2015年9期)2015-12-15
