
伊犁河谷不同管理模式稻田节肢动物群落结构及多样性
2017-11-01 12:01王小武丁新华吐尔逊付开赟班小莉付文君关志坚郭文超
新疆农业科学 2017年10期
王小武, 丁新华 ,吐尔逊,付开赟, 何 江 ,班小莉,付文君,关志坚,刘 文,郭文超
(1.新疆农业科学院微生物应用研究所,乌鲁木齐 830091;2.新疆农业科学院植物保护研究所/农业部西北荒漠绿洲作物有害生物综合治理重点实验室,乌鲁木齐 830091;3.伊犁州农业技术推广总站,新疆伊宁 835000;4.察布查尔县农业技术推广站,新疆察布查尔 835300;5.伊宁县农业技术推广中心,新疆伊宁 8351003)
伊犁河谷不同管理模式稻田节肢动物群落结构及多样性
王小武1, 丁新华2,吐尔逊2,付开赟2, 何 江2,班小莉3,付文君3,关志坚4,刘 文5,郭文超1
(1.新疆农业科学院微生物应用研究所,乌鲁木齐 830091;2.新疆农业科学院植物保护研究所/农业部西北荒漠绿洲作物有害生物综合治理重点实验室,乌鲁木齐 830091;3.伊犁州农业技术推广总站,新疆伊宁 835000;4.察布查尔县农业技术推广站,新疆察布查尔 835300;5.伊宁县农业技术推广中心,新疆伊宁 8351003)
目的研究伊犁河谷稻区节肢动物群落及各亚群落结构、时序格局和变化规律,为水稻生境昆虫综合治理提供依据。方法应用灰色关联度和主分量方法,分析该稻区不同模式节肢动物群落。结果共获节肢动物21 258头,隶属于2纲,10目,28科,31属,35种,生态调控型(记“A模式”)、常规管理型(记“B模式”)和粗放管理型(记“C模式”)三种模式下节肢动物物种数之间差异均不显著(P>0.05),但A、C模式物种数明显高于B模式。在发生量方面,A、B、C模式差异显著(P<0.05)。灰色关联度分析表明,水稻生境中,主要天敌为蜘蛛类且对稻水象甲具有一定的抑制作用,但其控害能力较弱;在群落特征指数和时序格局方面,A、C模式节肢动物群落较B模式,物种多样性指数、均匀度指数、丰富度指数均偏高,表明人工干预越少,节肢动物多样性、物种丰富度越大,生态系统越稳定;B模式下,多样性指数、丰富度指数、均匀度指数均最低,而优势集中性指数最高,表明该生境部分节肢动物种群分布较集中、数量大,生态系统趋于简单化,优势种害虫易爆发成灾。结论外界干涉越频繁,对节肢动物的分布、物种类群多样性均不利,节肢动物结构较单一,生态系统越不稳定;影响伊犁河谷稻区节肢动物群落结构变化主要因子为植食类、捕食类亚群落。
伊犁河谷稻区;群落结构;时序格局
0 引 言
【研究意义】新疆地处我国西北边陲、亚欧大陆腹地[1],以绿洲为载体,灌溉型农业为主,光热资源丰富,适宜多种农作物生长。素有“塞外江南”美誉的伊犁河谷既是新疆稻米的主产区,也是新疆实施“有机稻米生产”的主要示范区。目前该稻区水稻主要以直播和移栽2种栽培模式为主,因其管理方式的不同,移栽稻区又可分为常规移栽稻和有机移栽稻区。伊犁河谷在实施“有机稻米生产示范项目”之前,该稻区主要采用单一的化学药剂控制害虫,不仅使得稻水象甲等主要害虫反复猖獗,还对该稻区节肢动物群落结构造成严重破坏。近年来随着有机稻米生产基地的建成、项目的实施及水稻有机化发展,促使对稻田害虫的防治更注重生态系统自身的调控作用。节肢动物群落结构是研究群落多样性、稳定性、种间关系、演替以及种群发展的基础,也是充分发挥天敌自然控害作用、有效实施害虫综合治理的重要依据[2-3]。稻田节肢动物群落是以水稻为中心、食物链为基础多种节肢动物共存的动态、复杂网络系统[3-4],较自然其他生态系统,虽在营养结构、物种组成及稳定性等方面均有不同程度的降低,但天敌在有害生物种群控制中仍起到重要的作用[5],再者,稻田生态系统是人为干扰下的生态系统,其节肢动物群落多样性与稳定性,不仅仅取决于物种数、种间关系、食物网的复杂程度、水稻的生育阶段,还受人为因素影响较大;此外,害虫猖獗也往往与群落生态系统紊乱有关[6]。为此,把稻田节肢群落作为切入点,从群落生态学角度出发,探明节肢动物种群及亚群落间相互制约、相互联系、相互作用机制[7-12],不仅关系到有效实施害虫综合治理措施,而且也关系到现代新型农业可持续发展。【前人研究进展】目前,针对稻田节肢动物群落结构研究已有相关报道,涉及不同灌溉方式[11]、不同品种[3,13]、不同栽培方式[13]、不同生境[7-8,14]等方面,但自稻水象甲入侵新疆以来,其成功定殖该地区后对该地区节肢动物的影响,尤其是不同管理模式下节肢动物群落的影响,加之有关新疆荒漠绿洲稻区稻田节肢动物群落结构的研究及有关稻水象甲自然天敌控害方面的研究还尚未见报道,且节肢动物群落结构常因地理位置、耕作制度、气候条件、防治策略、品种、水肥管理等因素的干扰而存有差异[6-8],【本研究切入点】以伊犁河谷不同栽培管理模式的稻区为调查样点,调查并分析不同模式节肢动物及各亚节肢动物群落结构的多样性、稳定性及优势种天敌,在此基础上,通过分析不同模式益害比,揭示自然条件下天敌对主要害虫的控制作用,应用主成分法探讨不同栽培管理模式下影响节肢动物群落结构变化的主导因子和内部机制,为伊犁河谷水稻的有机化发展、对有害节肢动物最优综合治理及时准确的采取提供依据。
1 材料与方法
1.1 材 料
根据新疆伊犁河谷稻区当地生产实际中稻田的管理情况,将该稻区划分Ⅲ种模式,I、Ⅱ、Ⅲ模式水稻品种均为农林315,面积约为0.766 hm2,除水稻栽培及管理方式不同外,土质、海拔、地势等自然条件基本一致。
A生态调控型(记“A模式”):冬季清洁田埂杂草,4月中旬育秧,5月上旬炼苗,5月中下旬移栽,6月中旬施有机肥(速溶型),7月人工除草1~2次,7月中旬晒田,8月人工除草1~2次,10月上旬收获。年用鱼藤酮(内蒙古清源宝生物科技有限公司)、除虫菊素(内蒙古清源宝生物科技有限公司)、白僵菌(山西省科谷生物农药有限公司)等生物源药剂1~2次防除稻水象甲。
B常规管理型(记“B模式”): 4月中旬育秧,5月上旬炼苗,5月中下旬移栽,6月下旬7月上旬两次田间化学除草,7月中旬晒田,9月中下旬收获。年用药2~3次,主要为康宽(氯虫苯甲酰胺)、福奇(氯虫·高氯氟)、敌稗(N-3,4-二氯苯基丙酰胺)等化学药剂。
C粗放管理型(记“C模式”): 4月中旬直播,生育期偶有化学防控虫草,无其他管理措施。
1.2 方 法
1.2.1 网捕法
采用网捕法,直径0.33 m,摆幅2 m左右,往返为一复网,共50复网,7 d调查一次,雨天顺延。将扫网采集到的标本放入内置乙醚棉球的保鲜盒中,带回实验室进行挑选、数量统计、鉴定。
1.2.2 目测法
目测调查并统计稻株叶冠、茎秆及水面上的节肢动物种类、数量,每次调查15~20穴[8,14]。
1.3 数据处理
用Excel2010计算群落丰富度Pi、Shannon-Wiener多样性指数H′、Pielou均匀度指数J、Simpson 优势集中性指数C、优势度I等参数,对稻田节肢动物群落结构进行分析,各参数具体计算如下:
Pi=ni/N;H′=-∑Pi(lnPi);J=H'/lnS;C=∑(Pi)2;I=Nmax/N
其中ni为i物种出现的个体数;N为稻田节肢动物群落的总个体数,S为稻田节肢动物群落物种数,Nmax为稻田节肢动物群落中数量最多的物种的个体数。
1.3.1 灰色关联度分析,具体计算公式参考宋备舟等[10]进行。
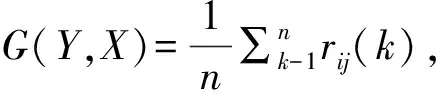
rij(k)=


1.3.2 主分量
为探明影响节肢动物群落结构的主导因子及内部机制[15],将各节肢动物( 按不同功能团) 的物种数量、个体数为变量,以植食类亚群落节肢动物物种数为X1,个体数为X2,捕食类节肢动物物种数为X3,个体数为X4;中性及寄生类节肢动物物种数为X5,个体数为X6,进行主分量分析[16]。
2 结果与分析
2.1伊犁河谷稻区节肢动物群落组成特征
调查共获节肢动物21 258头,隶属于2纲,10目,28科,31属,35种;其中,A模式 2纲,10目,28科,31属,35种;而双翅目(Diptera)、半翅目(Hemiptera)、鞘翅目(Coleoptera)、蜻蜓目(Odonata)和蜘蛛目(Araneida)等5个目节肢动物种数占该模式总种数的68.58%;B模式 2纲,10目,22科,24属,28种; 其中,双翅目(Diptera)、鞘翅目(Coleoptera)和蜘蛛目(Araneida)等3个目占总种数的46.44%;C模式 2纲,10目,24科,28属,31种;蜘蛛目(Araneida)、半翅目(Hemiptera)、鞘翅目(Coleoptera)和蜻蜓目(Odonata)等4个目节肢动物种数占总种数的54.83%。方差分析表明,三种模式下节肢动物物种数之间差异均不显著(P>0.05),但A、C模式物种数明显高于B模式。在发生量方面,A、B、C模式差异显著(P<0.05)。表1~3
2.2伊犁河谷稻区主要天敌与稻水象甲灰色关联度
确定了伊犁河谷稻区主要害虫类群为:稻水象甲。主要天敌类群为:瓢虫、食蚜蝇、蜘蛛、姬猎蝽和草蛉。A、B、C三种不同管理模式下,稻水象甲与主要天敌的关联度大小顺序分别为:蜘蛛>草蛉>食蚜蝇>姬猎蝽>瓢虫;蜘蛛>瓢虫>食蚜蝇>草蛉>姬猎蝽;蜘蛛>食蚜蝇>姬猎蝽>瓢虫>草蛉。可以看出,A、B、C三种不同模式下蜘蛛与和稻水象甲关联度最大,表明蜘蛛对稻水象甲具有控害效应。表4
表1 不同模式节肢动物群落组成
Table 1 Composed of arthropod community in different habitats in yili River Valley
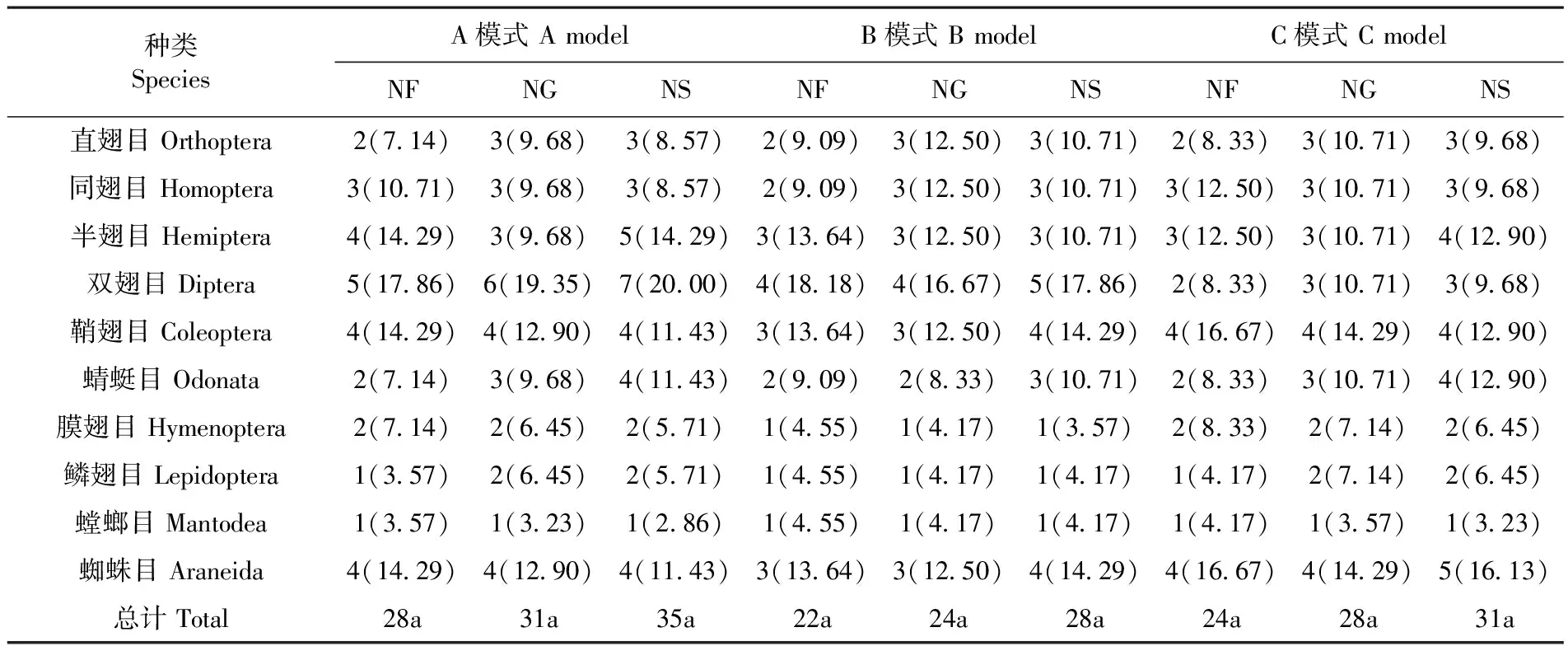
种类SpeciesA模式AmodelB模式BmodelC模式CmodelNFNGNSNFNGNSNFNGNS直翅目Orthoptera2(714)3(968)3(857)2(909)3(1250)3(1071)2(833)3(1071)3(968)同翅目Homoptera3(1071)3(968)3(857)2(909)3(1250)3(1071)3(1250)3(1071)3(968)半翅目Hemiptera4(1429)3(968)5(1429)3(1364)3(1250)3(1071)3(1250)3(1071)4(1290)双翅目Diptera5(1786)6(1935)7(2000)4(1818)4(1667)5(1786)2(833)3(1071)3(968)鞘翅目Coleoptera4(1429)4(1290)4(1143)3(1364)3(1250)4(1429)4(1667)4(1429)4(1290)蜻蜓目Odonata2(714)3(968)4(1143)2(909)2(833)3(1071)2(833)3(1071)4(1290)膜翅目Hymenoptera2(714)2(645)2(571)1(455)1(417)1(357)2(833)2(714)2(645)鳞翅目Lepidoptera1(357)2(645)2(571)1(455)1(417)1(417)1(417)2(714)2(645)螳螂目Mantodea1(357)1(323)1(286)1(455)1(417)1(417)1(417)1(357)1(323)蜘蛛目Araneida4(1429)4(1290)4(1143)3(1364)3(1250)4(1429)4(1667)4(1429)5(1613)总计Total28a31a35a22a24a28a24a28a31a
注:NF:科数;NG:属数;NS:种数,括号内为其所占该生境总科数、总属数或总种数比例,表中不同小写字母表示经0.05水平上的LSD差异检验(P<0.05 )
Note: NF:Number of Families;NG:Number of Genus;NS:Number of Species, Data in brackets were the ratio of families、genus and species, Data followed different lower and uppercase letters were significantly different atP< 0.05
表2 不同模式稻田节肢动物群落物种数方差
Table 2 Analysis of variance for different model number of arthropod species in paddy fields

变异来源Sourceofvariation平方和Quadraticsumdf均方差MeansquareerrorF值F-value显著水平Significancelevel组间amonggroups1337647266882352734817不显著组内intra-group1173882482445588总变异sumofvariation130764750
表3 不同模式稻田节肢动物群落个体数方差
Table 3 Number of arthropod community in rice field of different patterns of individual analysis of variance

变异来源Sourceofvariation平方和Quadraticsumdf均方差MeansquareerrorF值F-value显著水平Significancelevel组间amonggroups1745587287279375657005显著组内intra-group7405702481542855总变异sumofvariation91512950
表4 不同模式下稻水象甲与主要天敌的灰色关联度
Table 4 Relational grade of the Lissorhoptrus oryzophilus and their natural enemy groups in different mode

栽培模式Culturemode物种Species稻水象甲Lissorhoptrusoryzophilus关联Relationalgrade排序SortA模式Amode瓢虫Coccinellidae025865姬猎蝽NabispalliferusHsiao029684食蚜蝇Syrphidae031333蜘蛛Araneida054621草蛉Chrysopaperla039922B模式Bmode瓢虫Coccinellidae035952姬猎蝽NabispalliferusHsiao028975食蚜蝇Syrphidae033453蜘蛛Araneida047491草蛉Chrysopaperla030054C模式Cmode瓢虫Coccinellidae029974姬猎蝽NabispalliferusHsiao038263食蚜蝇Syrphidae039482蜘蛛Araneida041951草蛉Chrysopaperla028435
灰色关联度分析是基于灰色过程的一种相对的排序分析,其主要描述两个系统或因素发展过程中相对变化的情况,即其间的相似程度来判断其联系是否紧密。若两者在发展过程中相对变化基本一致或接近,则关联度就越大,反之就越小[17-18]。利用该理论研究并分析稻田稻水象甲与自然天敌之间的消长关系。若天敌与稻水象甲之间的关联度较高,说明该天敌对其的控制能力越强,反之,则弱。研究表明,伊犁河谷稻区蜘蛛与稻水象甲的关联度最大,对稻水象甲和蜘蛛进行进一步的益害比分析,6月11日前由于稻田蜘蛛个体数量较低,因而益害比也较低。6月18日以后随稻田稻水象甲数量增多,益害比也随之增大,A、C模式7月中旬达最大。因而益害比也随之迅速升高,并于7 月16日同时达到最高峰,而B模式因7月中旬进行田间化学控害,致使其益害比值最低。8月中旬,随着稻田稻水象甲羽化数量的逐渐增多,其益害比也逐渐增大。表4,图1

图1 不同模式下益害比时序动态
Fig.1 Temporal dynamics of predator prey ratio in different mode
2.3不同模式下节肢动物群落及亚群落特征指数和时序动态
2.3.1 不同模式下节肢动物群落及亚群落特征指数
根据食性关系,将伊犁河谷稻区节肢动物群落划分为,植食性亚群落、捕食性亚群群落、中性及寄生性亚群落3个功能类群[16]。研究表明,从节肢动物总群落来看,A、C模式节肢动物群落较B模式,物种多样性指数、均匀度指数、丰富度指数均偏高,而优势度指数、优势集中性指数偏低,表明人工干预越少,节肢动物多样性、物种丰富度越大,生态系统越稳定;B模式下,多样性指数、丰富度指数、均匀度指数均最低,而优势集中性指数最高,表明该生境节肢动物较A、C生境,部分节肢动物种群分布较集中、数量大,生态系统趋于简单化,优势种害虫易爆发成灾。从节肢动物亚群落来看,A、C模式捕食性节肢动物亚群落较B模式,优势度指数、丰富度指数及优势集中性指数均偏大,加之A、C模式下植食性亚群落多样性指数、丰富度指数及优势集中性指数较B模式均偏高,表明A、C模式捕食类节肢动物物种较多且聚集部分优势种天敌。而B模式下,植食性、捕食性亚群落均匀度指数和优势度指数均低于A、C模式,表明外界干涉越频繁,对节肢动物的分布、物种类群多样性均不利,节肢动物结构较单一,生态系统越不稳定。表5
表5 不同模式稻田节肢动物群落特征比较
Table 5 Comparison of different model characteristics of arthropod community in rice fields

处理Treatment功能团Functionalgroup多样性指数Diversityindex(H′)均匀度指数Evennessindex(J)优势度指数DominanceIndex(I)丰富度指数RichnessIndex(lnS)优势集中性指数ConcentrationIndex(C)A节肢动物群落2758608279015933555300861植食性亚群落1749908415028202639102044捕食性亚群落2315509318025592772601282中性和寄生性亚群落0958413827059531386304593B节肢动物群落2508407055018283332201170植食性亚群落1819906896025792079401951捕食性亚群落2297608287025482484901270中性和寄生性亚群落0624704506082440693106939C节肢动物群落2738007973015703434000867植食性亚群落1746607284027612397902036捕食性亚群落2274408618025592639101290中性和寄生性亚群落0986305505058131791804489
注:表中丰富度指数S取其自然对数lnS
Note: the richness index S in the table takes its natural logarithm lnS
2.3.2 不同模式下节肢动物群落及亚群落特征指数时序动态(图2)
A、C模式较B模式,节肢动物总群落及各亚群落多样性指数较平稳,全年波动较小(图2a,3a,4a)。B模式因农事操作及化学防虫控草,致使物种数、发生量均最少,多样性变化较大,波动明显。A模式物种数、发生量最大,种间制约程度高,波动小,C模式次之。
不同模式下节肢动物群落及亚群落均匀度波动(图2b,3b,4b),具体为整个生育期中,A、C模式较B模式,节肢动物总群落、植食类亚群落及捕食类亚群落均匀度波动程度不大且变化趋势相似,但不同模式下中性及寄生类亚群落均匀度波动明显。对于A模式,7月上中旬陆续开始晒田,8月中旬正值人工除草,田间间歇性灌水所致;C模式,因管理水平较差,稻田忽干忽湿,致使中性类双翅目等喜水性节肢动物数量变化幅度较大。 B模式,7月中旬节肢动物总群落和植食类亚群落均匀度不同程度呈现“V”字形变化,此现象由B模式田间管理措施(田间化学除草、控虫)所致。
不同模式下节肢动物群落及亚群落优势度指数和优势及中性指数变化趋势与多样性指数变化趋势相反,具体为:中性及寄生类亚群落>植食类亚群落>捕食类亚群落>节肢动物总群落(图2c,2d,3c,3d,4c,4d)。A、C模式,整个生育期优势度指数和优势及中性指数变化差异不显著,B模式6月上旬到7月下旬,因施肥、化学防控外界因素影响,使得优势度指数和优势及中性指数变化较大。8月中下旬,中性及寄生类亚群落优势度、优势集中性指数呈正态变化。此现象由于生育中后期水稻生境水位较高,致使中性类双翅目等喜水性节肢动物田间分布数量大且集中所致。
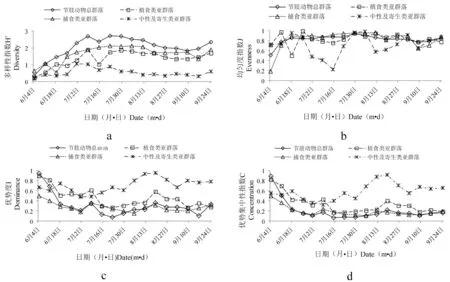
图2 A模式下节肢动物群落及亚群落特征指数时序动态
Fig.2 Temporal dynamics of characteristic index of arthropod community and its sub-community in the A mode
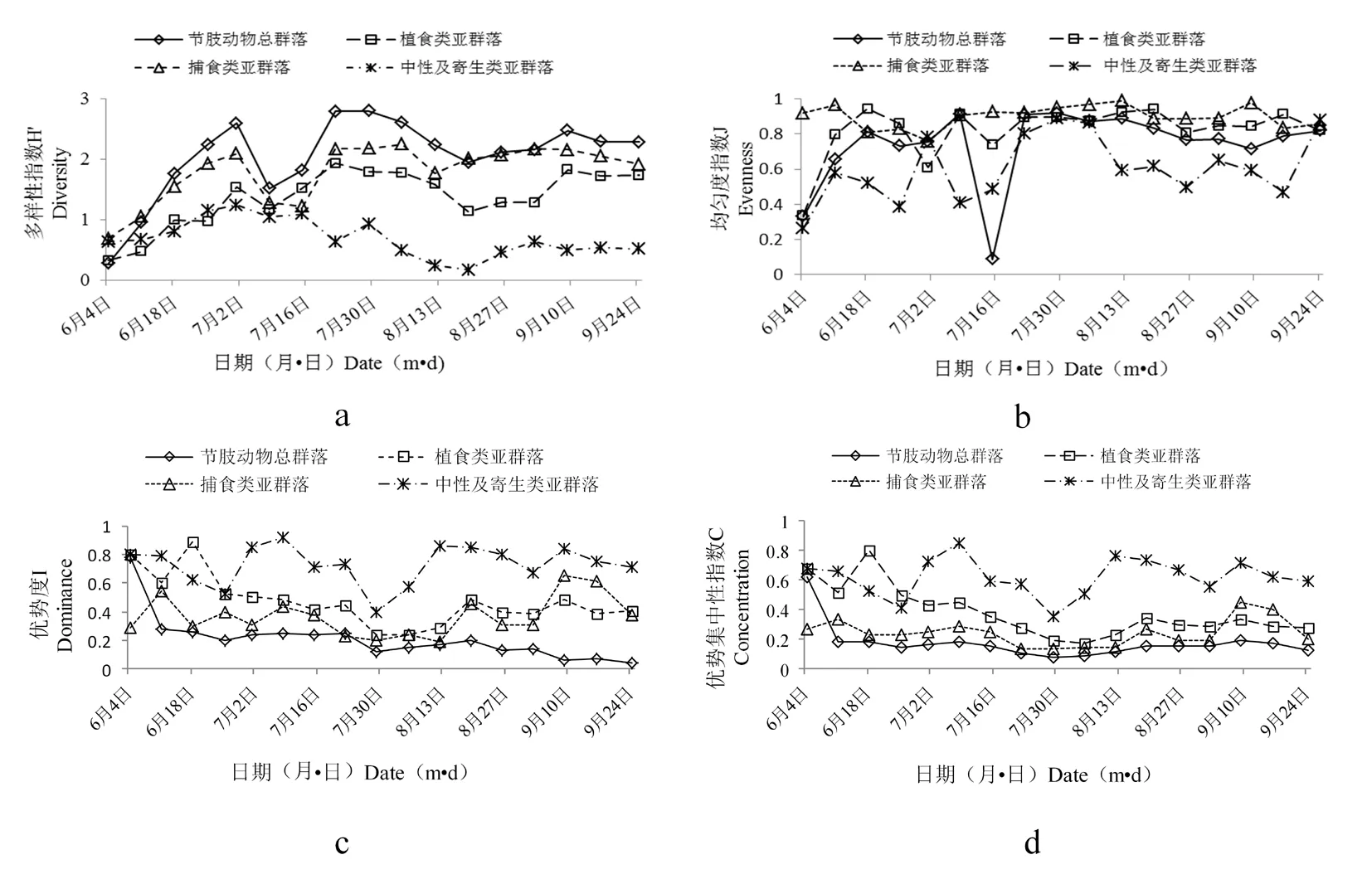
图3 B模式下节肢动物群落及亚群落特征指数时序动态
Fig.3 Temporal dynamics of characteristic index of arthropod community and its sub-community in the B mode

图4 C模式下节肢动物群落及亚群落特征指数时序动态
Fig.4 Temporal dynamics of characteristic index of arthropod community and its sub-community in the C mode
2.4 不同模式下节肢动物主分量
3种管理模式下各节肢动物亚群落3个主分量累计贡献率均高于85% , 其中,A模式节肢动物群落累计贡献率高达 90.1%,第一主分量代表了捕食类节肢动物个体数、物种数及植食类节肢动物个体数的综合因子,贡献率为50.6% ,说明了捕食类节肢动物物种数、个体数及植食类节肢动物个体数对群落的重要作用;第二主成分主要代表了植食类节肢动物物种数和中性及寄生类节肢动物物种数,贡献率达25.2%;第三主成分代表了中性及寄生类亚群落节肢动物物种数、个体数,贡献率达14.3%。因此A模式稻区植食类物种数、个体数、捕食类物种数、个体数及中性及寄生类节肢动物的物种数是本稻区节肢动物群落结构变化的主导因子。
B模式稻区第一主成分分量表示捕食类节肢动物物种数、个体数及植食类节肢动物物种数;第二主成分分量表示中性及寄生类亚群落节肢动物物种数、个体数;第三主成分分量表示植食类节肢动物数量和捕食类节肢动物物种数;3个主成分分量累计贡献率高达87.4%,表明B模式稻区捕食类、中性及寄生类亚群落物种数、个体数及植食类节肢动物物种数是该稻区节肢动物群落结构变化的主要因子。
C模式稻区第一主成分分量表示捕食类节肢动物个体数及植食类节肢动物物个体数;第二主成分分量表示捕食类节肢动物和中性及寄生类亚群落节肢动物物种数;第三主成分分量表示植食类节肢动物物种数和中性及寄生类亚群落节肢动物个体数;3个主成分分量累计贡献率达85.43%,表明C模式稻区植食类、捕食类亚群落节肢动物物种数和个体数是该稻区节肢动物群落结构变化的主要因子。表6
表6 新疆伊犁河谷不同模式节肢动物群落主成分
Table 6 Principal component analysis of arthropod community in the different mode

模式Mode主成分PrincipalcomponentX1X2X3X4X5X6累计贡献率CumulativeA10513074408750970-0204068750620767-04780356-00450715-02327583-02570182-0117-016706290566901B108620360094509330665-0.0885142-0165-04400054-0046055008467223-023108070091-0080-00970427874C106660824033109270609066748442021904400857000406010195713830663010801450017018105828543
3 讨 论
稻田节肢动物群落结构及多样性是稻田有害节肢动物治理的重要依据[19-20],用灰色关联度分析法和主分量分析法对伊犁河谷稻区节肢动物群落结构进行分析,结合水稻生长发育时期,按照群落的时序动态特点针对性采取以“生态调控”为核心的综合治理措施,是实现水稻有害节肢动物可持续治理的重要基础[21]。
为植食性类肢动物提供良好的生存环境。再者,其以“绿洲为载体”,这也为双翅目、蜻蜓目等节肢动物的生存创造了优良的条件。此外,植食类节肢动物亚群落的繁荣也为捕食类、寄生类节肢动物亚群落及河谷其他动物提供了良好的营养基础[16],是该稻区节肢动物群落及各亚群落结构稳定的前提条件,为群落的多样性奠定了基础。
天敌与害虫间的相互制约、依存的关系是在长期协同进化所致[9]。在植物生育期中,主要害虫可能存有多种天敌,而不同天敌因物种数、捕食量、食性、时空格局和搜索行为等方面的差异性[10],致使对主要害虫的制约呈现出复杂的相互关系。研究通过灰色关联度分析发现, A、B、C三种不同模式下蜘蛛与和稻水象甲关联度最大,表明蜘蛛对稻水象甲具有一定的控害效应,这一结果在关志坚等[22], 于凤泉等[23]研究中得以印证;此外,调查发现蜘蛛对稻水象甲其自然控害能力较弱,为避免打破该生境节肢动物之间的动态平衡,因此在稻水象甲发生量高于其防治临界值5.82头/m2时[24],建议采取生物措施来抑制其发生[25],造成这一结果的原因可能与稻水象甲的生活习性有关,需今后进一步研究。
节肢动物多样性指数是反映群落组织水平的主要指标,同时也反映不同生境节肢动物群落的发展状况[16,26]。“生态调控”是实现水稻有害节肢动物可持续治理的重要基础[21]。以功能团为单位,探讨节肢动物群落的结构及多样性,可使其复杂的群落网络结构简单明了,节肢动物各亚群间的关系更加明确[5]。万方浩等[26]对综防区和化防区稻田害虫-天敌群落组成及多样性研究中,发现综合防区节肢动物的多样性和丰富度均高于化防区;金翠霞等[27]对稻田节肢动物群落多样性研究,其认为杀虫剂的施用扩大了害虫亚群落和天敌亚群落之间的稳定性差距,打破其同步发展的关系,再者,杀虫剂的施用对天敌亚群落的物种数、个体数均具明显的抑制作用,尤其是对寄生性天敌影响更为显著,致使害虫再猖獗;王凯学等[14]、张清泉等[28]对生态稻田及常规稻田节肢动物群落结构特征比较研究中,发现生态稻田(生境多样性稻田)天敌亚群落的发展快于害虫亚群落,其物种数和个体数均早于或与害虫同步发生,在一定程度上对害虫具有一定的控制作用,而常规稻田各亚群落的多样性、丰富度及物种数均低于生态稻田。顾伟等[16]、郁晓蓓[19]和宋文军等[29]均认为,在研究节肢动物群落结构及时序动态时,只需综合考虑节肢动物群落多样性指数和均匀度指数,即可判定节肢动物群落的稳定性。韩争伟等[30](2013)认为节肢动动物群落的稳定性需要多个指标综合反映才能确定其稳定性。研究表明, 外界干涉越频繁,对节肢动物的分布、物种类群多样性均不利,致使节肢动物结构较单一,生态系统趋于简单化,优势种害虫易爆发成灾,与前人[14,26-28]研究结果较为一致。
4 结 论
研究于伊犁河谷稻区捕获节肢动物共35种,隶属于2纲,10目,28科,31属。优势节肢动物类群为鞘翅目、半翅目、同翅目、双翅目、蜻蜓目及蜘蛛目等。A、B、C三种不同管理模式下,稻水象甲与主要天敌的关联度大小顺序分别为:蜘蛛>草蛉>食蚜蝇>姬猎蝽>瓢虫;蜘蛛>瓢虫>食蚜蝇>草蛉>姬猎蝽;蜘蛛>食蚜蝇>姬猎蝽>瓢虫>草蛉。伊犁河谷稻区A、C模式下节肢动物总群落及各亚群落物种多样性指数、均匀度指数、丰富度指数较B模式均偏高,B模式,植食性、捕食性亚群落均匀度指数和优势度指数均低于A、C模式,主分量分析表明,不同模式节肢动物群落结构变化的主要因子为植食类、捕食类亚群落。表明该稻区节肢动物群落较复杂,外界干扰后,自我恢复能力较强。
References)
[1] 胡见义,彭苏萍,李建忠, 等. 新疆能源产业发展战略与思考 [J]. 中国能源,2012,34(1):8-13.
HU Xian-yi, PEN Su-ping, LI Jian-zhong,et al. (2012). The Development strategy of Energy industry in Xinjiang [J].EnergyofChina, 2012, 34(1) : 8-13. (in Chinese)
[2] 周玉锋. 苇状羊茅人工草地节肢动物群落研究 [D]. 贵阳:贵州大学博士研究生学位论文, 2009.
ZHOU Yu-feng. (2009).Researchonarthropodcommunityintallfescueartificialgrassland[D]. PhD Thesis. Guizhou University, Guiyang. (in Chinese)
[3] 郭玉人.沈阳地区稻田节肢动物群落结构及群落生态研究 [J] .生态学报,2001,21(11):1 854-1 862.
GUO Yu-ren. (2001). The constructions and ecology of arthropod community of paddies in Shenyang [J] .ActaEcologicaSinica,21(11): 1,854-1,862. (in Chinese)
[4] 刘志诚,叶恭银,胡萃,等. 转cry1 Ab/cry1 Ac基因籼稻对稻田节肢动物群落影响 [J] .昆虫学报,2003,46(4):454-465.
LIU Zhi-cheng, YE Gong-yin, HU Cu, et al. (2003). Impact of transgenic indica rice with a fused gene of cry1Ab cry1Ac on the rice paddy arthropod community [J] .ActaEntomologicaSinica, 46(4) : 454-465. (in Chinese)
[5] 郝树广,张孝羲. 稻田节肢动物群落优势功能集团的垂直分布,数量动态及天敌作用估计[J].应用生态学报,2000,11(1):103-107.
HAO Hu-guang, ZHANG Xiao-yi. (2000). Vertical distribution and quantitative dynamics of dominant functional groups of arthropod community in rice fields and estimation of natural enemy effects [J] .ChineseJournalofAppliedEcology, 11 (1) : 103-107. (in Chinese)
[6] 陶方玲,梁广文,庞雄飞. 不同生境区稻田节肢动物群落相似性分析 [J] .生态科学,1995,(2):54-59.
TAO Fang-ling, LIANG Guang-wen, PANG Xionbg-fei. (1995). The Analysis about the similarity of arthropod conmulities in rice field in different habitats [J].EcologicalScience, (2): 54-59. (in Chinese)
[7] 杨国庆, 吴进才, 张士新, 等. 三种类型稻田节肢动物群落结构、亚群落内禀增长率与链节数的关系[J]. 生态学报, 2004, 24(4): 686-692.
YANG Guo-qing, WU Jin-cai, ZHANG Shi-xin, et al. (2004). The relationship among structures, intrinsic rate of increase of functional guilds, and link numbers of arthropod community in three types of rice field under organic rice production system [J].ActaEcologicaSinica, 24(4): 686-692. (in Chinese)
[8] 王丽艳,朱莹,丛斌, 等.寒地稻田节肢动物群落多样性调查 [J] . 植物保护,2009,35(3):102-105.
WANG Li-yan, ZHU Ying, CONG Bing, et al. (2009). Investigation of the arthropod community diversity in cold rice fields [J] .PlantProtection,35(3): 102-105. (in Chinese)
[9] 巫厚长, 魏重生, 王方晓, 等. 各种天敌对烟蚜种群数量影响效果的灰色关联度分析[J]. 安徽农业大学学报,2002,29(3):224-229.
WU Hou-chang, WEI Chong-sheng, WANG Fang-xiao, et al. (2002). Grey relationalgrade analysis on natural enemies influencing Myzus persicae population.JournalofAnhuiAgriculturalUniversity, 29(3): 224-229. (in Chinese)
[10] 宋备舟, 王美超, 孔云,等. 梨园芳香植物间作区主要害虫及其天敌的相互关系[J]. 中国农业科学, 2010, 43(17):3 590-3 601.
SONG Bei-zhou, WANG Mei-chao, KONG Yun, et al. (2010). Interaction of the Dominant Pests and Natural Enemies in the Experimental Plots of the Intercropping Aromatic Plants in Pear Orchard [J].ScientiaAgriculturaSinica, 43(17): 3,590-3,601. (in Chinese)
[11] 邱佩,崔远来,罗玉峰, 等.淹灌和间歇灌溉对晚稻田节肢动物群落多样性影响[J].中国农村水利水电,2016,(8):113-117.
QIU Pei, CUI Yuan-lei, LUO Yu-feng, et al. (2016). Effect of flooding irrigation and intermiteent irrigation patterns on arthropod community diversity in late rice fields [J] .ChinaRuralWaterandHydropower, (8): 113-117. (in Chinese)
[12] 罗淑萍,张永强,黄寿山.不同抗性品种稻田捕食性节肢动物的群落结构[J]. 应用昆虫学报,2006,43(4):453-460.
LUO Shu-ping, ZHANG Yong-qiang, HUANG Shou-shan. (2006).Community structure of predatory arthropods in different resistant variety rice fields [J].ChineseJournalofAppliedEntomology, 43(4): 453-460. (in Chinese)
[13] 蔡万伦,石尚柏,杨长举,等.不同种植方式下转Bt基因水稻对稻田节肢动物群落的影响[J].昆虫学报,2005,48(4):537-543.
CAI Wan-long, SHI Shang-bai, YANG Chang-ju, et al. (2005). Difference of arthropod communities in Bt rice paddies under different cropping patterns [J].ActaEntomologicaSinica, 48(4): 537-543. (in Chinese)
[14] 王凯学,张清泉,陈丽丽, 等.生态稻田及常规稻田节肢动物群落结构特征的比较研究[J]. 植物保护,2013,39(3):31-35.
WANG Kai-xue, ZHANG Qing-quan, CHEN Li-li, et al. (2013). Comparative studies on arthropod community structure characteristics between ecological rice paddies and conventional rice paddies [J].PlantProtection, 39(3): 31-35. (in Chinese)
[15] 郝树广,张孝羲,程遐年,等. 稻田节肢动物群落营养层及优势功能集团的组成与多样性动态[J]. 昆虫学报,1998,(4):343-353.
HAO Shu-guang, ZHANG Xiao-yi, CHEN Xia-nian, et al. (1998). The Dynamics of biodiversity and the composition of nutrition classes and dominant guilds of arthropoda community in paddy field [J].ActaEntomologicaSinica, (4): 343-353. (in Chinese)
[16] 顾伟,马玲,丁新华,等. 扎龙湿地不同生境的昆虫多样性[J]. 应用生态学报,2011,22(9):2 405-2 412.
GU Wei, MA Ling, DING Xin-hua, et al. (2011). Insect diversity of different habitat types in Zhalong Wetland Northeast China [J].ChineseJournalofAppliedEcology, 22(9): 2,405-2,412. (in Chinese)
[17] 唐启义. DPS 数据处理系统-实验设计、统计分析及数据挖掘[M]. 北京:科学出版社,2010:1 060-1 061.
TANG Qi-yi.(2010).DPSDateprocessingSystem-ExperimentalDesign,StatisticalAnalysisAndDataMining[M].Beijing: Science Press: 1,060-1,061. (in Chinese)
[18] 贾伟宽, 王慧, 丁世飞,等. 基于熵权的灰色关联度分析天敌对棉铃虫的控制作用[J]. 长江大学学报(自然科学版), 2008, 5(4):4-7.
JIA Wei-kuan , WANG Hui, DING Shi-fei, et al. (2008). Analysis of Control of Cotton Bollworm by Natural Enemies Using Gray Incidence Degree Based on Entropy [J].JournalofYangtzeUniversity(NatSciEd.) , 5(4): 4-7. (in Chinese)
[19] 郁晓蓓. 两品种枣园节肢动物群落动态及枣瘿蚊与其天敌的关系[D]. 合肥:安徽农业大学硕士学位论文, 2010.
YU Xiao-pei. (2010).StudiesonthedynamicsofarthropodcommunitiesandtherelationsofCecidamyiidaewithitsnaturalenemiesintwojujubeOrchards[D]. Master Thesis. Anhui Agricultural University, Hefei. (in Chinese)
[20] 胡竞辉,王美超,孔云,等. 梨园芳香植物间作区节肢动物群落时序格局[J]. 生态学报,2010,30(17):4 578-4 589.
HU Jing-hui, WANG Mei-chao, KONG Yun, et al. (2010). Temporal structures of arthropod community of intercropping aromatic plants in pear orchard [J].ActaEcologicaSinica, 30(17): 4,578-4,589. (in Chinese)
[21] 江俊起. 不同类型和防治措施稻田害虫和天敌群落结构的研究[D]. 合肥:安徽农业大学硕士学位论文, 2004.
JIANG Qi-jun. (2004).StudiesontheStructuresofInsectPestandNaturalEnemyCommunitiesinpaddyfieldsofDifferentRiceTypesandunderDifferentInsecticideTreatments[D]. Master Thesis. Anhui Agricultural University, Hefei. (in Chinese)
[22] 关志坚,丁新华,付文君,等. 伊犁河谷地区稻水象甲生物学特性及其种群田间迁移规律的研究[J]. 新疆农业科学,2014,51(7):1 312-1 318.
GUANG Zhi-jian, DING Xin-hua, FU Wen-jun, et al. (2014). Study on the Occurrence Regulation of Rice Water Weevil (Lissorhoptrusoryzophilus)in the Ili River Valley[J].XinjiangAcademicofAgricultureScience, 51(7): 1,312-1,318. (in Chinese)
[23] 于凤泉, 李志强, 刘培斌,等. 稻水象甲生物防治研究进展[J]. 辽宁农业科学, 2003,(6):19-20.
YU Feng-quan, LI Zhi-qiang, LIU Pei-bing, et al. (2003). Research progress on biological control of rice water weevil [J].Liaoningagriculturalsciences, (6):19-20. (in Chinese)
[24] 丁新华, 王小武, 吐尔逊,等. 新疆荒漠绿洲稻区稻水象甲危害损失及防治阈值研究[J]. 生物安全学报, 2017, 26(1): 63-67.
DING Xin-hua, WANG Xiao-wu, Tursun, et al. (2017). Establishing the harm loss and economic threshold ofLissorhoptrusoryzophilusKuschel for Xinjiang desert oases [J]JournalofBiosafety, 26(1):63-67. (in Chinese)
[25] 王小武, 丁新华, 吐尔逊,等. 不同生物药剂对稻水象甲的毒力、拒食活性及防效分析[J]. 生物安全学报, 2017, 26(1):68-74.
WANG Xiao-wu,DING Xin-hua, Tursun, et al. (2017). Toxicity antifeedant activity and control effect of different biological insecticides toLissorhoptrusoryzophilusKuschel [J].Journalofbiosafety, 26(1): 68-74. (in Chinese)
[26] 万方浩, 陈常铭. 综防区和化防区稻田害虫-天敌群落组成及多样性的研究[J]. 生态学报,1986,(2):159-170.
WAN Fang-hao, CHEN Chan-ming. (1986). Studies on the structure of the rice pest-natural enemy community and diversity wnder IPM Area and chemical control area [J].ActaEcologicaSinica, (2): 159-170. (in Chinese)
[27] 金翠霞,吴亚,王冬兰. 稻田节肢动物群落多样性[J]. 昆虫学报,1990,(3):287-295.
JIN Cui-xia, WU Ya, WANG Dong-lan. (1990). Diversity of arthropod communities in paddy fields [J].ActaEntomologicaSinica, (3): 287-295. (in Chinese)
[28] 张清泉,王华生,覃保荣,等. 生态稻田节肢动物群落结构及其多样性研究[J]. 中国植保导刊,2014,34(4):19-24.
ZHANG Qing-quan, WANG Hua-sheng, QIN Bao-run, et al. (2014). Study on community structure and diversity of arthropods in ecological rice paddy fields [J].ChinaPlantProtection, 34(4): 19-24. (in Chinese)
[29] 宋文军,马玲,王慧,等. 扎龙自然保护区湖泊边昆虫群落多样性[J]. 东北林业大学学报,2007,35(7):80-81.
SON Wen-jun, MA Ling, WANG Hui, et al. (2017).Diversity of Insect Communities by the Lakeside in Zhalong Nature Reserve [J].JournalofNortheastForestryUniversity, 35(7): 80-87. (in Chinese)
[30] 韩争伟,马玲,曹传旺,等. 太湖湿地昆虫群落结构及多样性[J]. 生态学报,2013,33(14):4 387-4 397.
HAN Zheng-wei,MA Ling, CAO Chuan-wang, et al. (2013).The structure and diversity of insect community in Taihu Wetland [J].ActaEcologicaSinica, 33(14): 4,387-4,397. (in Chinese)
TheStructureandDiversityofArthropodCommunitiesinPaddyFieldsunderDifferentManagementModelsinYiliRiverValley
WANG Xiao-wu1,DING Xin-hua2,Tursun2,FU Kai-yun2,HE Jiang2,BAN Xiao-li3,FU Wen-jun3,GUAN Zhi-jian4,LIU Wen5, GUO Wen-chao1
(1.ResearchInstituteofAppliedMicrobiology,XinjiangAcademyofAgriculturalSciences,Urumqi830091,China; 2.ResearchInstituteofPlantProtection/KeyLaboratoryofIntegratedPestManagementofCropsinChinaNorth-westernOasis,MinistryofAgriculture,P.R.China,XinjiangAcademyofAgriculturalSciences,Urumqi830091,China; 3.AgriculturalTechnologyExtensionMasterStationofIliPrefecture,YiningXinjiang835000,China; 4.AgriculturalTechnologyExtensionStationofQapqalCounty,QapqalXinjiang835000,China; 5.AgriculturalTechnologyExtensionStationofYiningCounty,YiningXinjiang835100,China)
ObjectiveIn order to explore the structure, the temporal dynamics and the change regularity of arthropod communities and its sub-communities in paddy fields of Ili river valley.MethodArthropods on different modes were investigated by gray correlation degree analysis, principal component analysis and. clustering analysis.ResultA total of 21,258 arthropods were collected, which belonged to 2 classsess,10 orders, 28 families and 35 species. There was no significant difference between the number of different model arthropod species (P>0.05), while their reproduction quantity had significant difference (P<0.05). Gray correlation degree analysis showed that spiders had a certain effect on rice water weevil, but its natural control ability to rice water weevil was weak. In terms of community characteristics index and temporal pattern, the arthropod communities in Aand C models were higher than those in the B model and species diversity index, evenness index and richness index were all higher, which indicated that when the less artificial intervention, the greater arthropod diversity and species richness would be and he more stable the ecosystem would be, too. Under the B model, the diversity index, richness index and evenness index were the lowest while the advantage concentration index was the highest, which showed that the arthropod population in the habitat was more concentrated, the quantity was large, the ecosystem tended to be simple, and the dominant species were easy to outbreak.ConclusionThe more frequent external interference, the less unfaborable distribution of arthropod and diversity of species groups would be. The more simple and single the arthropod structure was, the less stable the ecosystem would be. The main factors affecting arthropod animal community structure changes in Yili River Valley are phytophagous and predacious sub community.
Ili river valley; arthropod community; temporal dynamics
GUO Wen-chao(1966-),male,born in Hebei,Research,doctoral supervisor . Research area:Agricultural invasive pests biological control and biological control. (E-mail)gwc1966@163.com
S562;S435.1
A
1001-4330(2017)10-1875-12
10.6048/j.issn.1001-4330.2017.10.013
2017-08-12
新疆维吾尔自治区创新环境(人才、基地)建设专项-自然科学基金(2017D01B29)
王小武(1990-),男,宁夏海原人,硕士研究生,研究方向为农业昆虫与害虫防治,(E-mail)wxw303528@163.com
郭文超(1966-),男,河北人,研究员,博士生导师,研究方向为害虫生物防治和农业外来入侵生物防控,(E-mail)gwc1966@163.com
Supported by: Special Fund for Innovative Environment (Talent & Base) Construction Project of Xinjiang Natural Science Fundation (2017D01B29)
猜你喜欢
科学大众(中学)(2021年3期)2021-05-17
落叶果树(2020年2期)2020-04-20
——稻(二)
种业导刊(2019年3期)2019-05-21
课外生活(小学1-3年级)(2018年10期)2018-11-19
小猕猴学习画刊(2018年9期)2018-09-22
现代园艺(2017年22期)2018-01-19
现代园艺(2017年21期)2018-01-03
山东农业科学(2017年3期)2017-03-29
种业导刊(2015年3期)2015-01-22
种业导刊(2014年3期)2014-01-23
