
Nrf2在肺纤维化治疗中作用的研究进展
2018-03-19 03:28王玉敏
中国老年学杂志 2018年5期
陈 雁 王玉敏
(赤峰学院附属医院呼吸内科,内蒙 赤峰 024005)
肺纤维化(PF)是一种慢性、进展性、不可逆转的肺部疾病,临床表现为逐渐加重的呼吸困难,患者最终因肺功能严重受损发生呼吸衰竭而死亡。目前还没有方法可以完全阻断PF的发生。PF的发病机制仍未完全阐明,最近研究〔1〕显示氧化应激反应参与PF发病机制,因此,抑制氧化应激导致的肺损伤已成为防治PF的新策略。本文总结了肺部发生纤维化过程中氧化应激损伤的来源及其在PF形成中的作用,并对以转录因子(Nrf2)抗氧化通路为靶点治疗PF的进展进行了总结。
1 PF过程中氧化应激来源
氧化应激(oxidative stress)是指活性氧产生增多和/或抗氧化防御耗竭产生的一系列分子、细胞和组织的功能损害〔1〕。与机体其他器官相比,肺暴露于高水平的氧,使其更容易受到氧化应激的攻击从而产生损害,吸入的空气中氧的压力达到150 mmHg,而肺泡气氧的压力可达到100 mmHg。PF过程中氧化应激来源主要包括线粒体、还原型辅酶Ⅱ(NADPH)氧化酶和抗氧化防御体系酶的耗竭。
1.1线粒体介导产生活性氧(ROS) 线粒体产生ROS是介导PF产生的主要参与因素。线粒体功能障碍主要产生包括过氧化氢(H2O2)和超氧负离子(O2·-)在内的活性氧簇〔2〕。线粒体电子传递链(ETC) 是细胞内ROS产生的主要途径。ETC是O2·-的主要产生来源。大约2%的线粒体氧耗转变成O2·-。ETC是生物体三磷酸腺苷(ATP)能量的来源,在能量传递的过程中,一小部分电子发生泄漏,泄漏的电子被O2接收即生成O2·-。其中复合体Ⅰ和复合体Ⅳ产生ROS的能力较强。研究显示,肺泡巨噬细胞(AM)线粒体介导产生的H2O2在引起石棉沉滞症中的作用〔3〕,AM暴露于石棉样纤维(asbestos fibers)后可诱导H2O2的产生,而这一作用可被氧化氢酶(CAT)阻断,AM线粒体应激水平减弱。另外,Rho结合蛋白家族成员Rac1增加AM线粒体H2O2的产生,而敲低ETC复合体Ⅲ的ROS产生位点铁硫蛋白能够减弱石棉诱导的AM H2O2的产生。石棉沉滞症患者AM线粒体Rac1的表达水平升高,而条件性删除Rac1基因的小鼠肺内氧化应激水平较低和PF发生程度减轻。总之,石棉能够通过将电子由复合体Ⅲ转移给Rac1从而促进AM H2O2的产生,触发一系列下游事件,导致石棉沉滞症的发生。因此,推断Rac1可以作为PF的一个新的分子标志物。H2O2参与PF发生的另一项证据来源于CAT能够减弱石棉沉滞症啮齿类动物模型PF的发生〔4〕。另外,一些外源性毒素,如石棉样纤维能够激活参与PF发生的非内皮细胞线粒体ROS的产生〔5〕。
1.2NADPH氧化酶 NADPH氧化酶在肺内ROS的产生中发挥重要作用。NADPH氧化酶是一个与细胞膜结合的、含有多亚基的酶复合体,其主要功能是实现电子从NADPH跨细胞膜传递给分子氧,产生O2·-及其下游的ROS。NADPH氧化酶家族由NOX2,NOX1,NOX3,NOX4,NOX5,DUOX1和DUOX2 NADPH氧化酶7个成员组成,每个成员通过与细胞膜结合的催化亚基“Nox”或“Duox” 的不同进行区别。NADPH氧化酶催化亚基gp91phox/NOX2及其6个同源物统称为NOX家族蛋白。这些NOX家族蛋白在不同组织中具有独特的表达分布。当细胞受到刺激后4个细胞质成分募集到细胞膜上,与两个膜结合的成分组成有活性的NADPH氧化酶复合体,促进ROS的产生〔6〕。NADPH 氧化酶的主要功能是产生O2·和H2O2产生,从而参与细胞信号转导、杀死外来微生物的侵入等〔7〕。
最近研究显示NADPH氧化酶介导产生的ROS在PF过程中发挥重要作用,尤其是NOX1,NOX2,NOX4-NADPH氧化酶参与PF发病机制〔7〕。转化生长因子-(TGF)-β诱导激活NOX4-NADPH 氧化酶,从而通过促进肌成纤维细胞分化和上皮再形成。TGF-β和氧化应激共同促进PF形成〔8〕。TGF-β是最强效促纤维化细胞因子,通过调控细胞的增殖、分化、黏附、迁移、凋亡及上皮细胞间质转型(EMT),进而在细胞外基质(ECM)沉积的过程中发挥重要作用。研究显示,NOX4-NADPH氧化酶介导产生的H2O2对于TGF-β诱导的成纤维细胞分化和ECM产生是必须的,NOX4基因敲除或药理学途径抑制NOX4可以阻止博来霉素诱导的小鼠PF的发生〔9〕。同时博来霉素能够诱导小鼠肺内NOX4的表达升高,PF患者肺内NOX4的表达也是升高的〔9〕。TGF-β1诱导的成纤维细胞分化与细胞外H2O2的产生相关,后者促进肺泡上皮细胞(AEC)死亡。AEC死亡是PF的重要始动事件和PF持续存在的主要事件〔10〕。NOX4在PF患者增生的肺泡上皮中强效表达,尤其在2型肺泡细胞(AT2)中表达明显〔10〕。NOX4基因缺失,可阻止博来霉素诱导的小鼠AT2细胞凋亡的发生〔11〕。NOX-4化学性抑制剂处理来源于NOX4基因缺失的内皮细胞或AEC,能够降低TGF-β1介导的ROS产生和AEC凋亡。综上所述,NOX4-NADPH氧化酶介导产生的ROS参与AEC凋亡机制,进而促进PF发生。
NOX2-NADPH氧化酶同时也参与PF的发生。最近研究发现,NOX2基因缺失的小鼠,能够阻止博来霉素诱导的小鼠PF的发生〔7〕。
1.3抗氧化防御体系酶的耗竭 肺本身表达一些抗氧化酶,从而保护肺脏因氧化应激引起损伤。抗氧化防御体系的酶的缺失能够促进氧化应激的发生,进而促进PF的发生。
1.3.1CAT CAT可促使H2O2分解为分子氧和水(2 H2O2→2 H2O+O2),清除体内的过氧化氢,从而使细胞免于遭受H2O2的毒害,是生物防御体系的关键酶之一。CAT 是AEC和炎症细胞内H2O2的清除剂,能够抑制H2O2介导的PF患者肺内成纤维细胞的活化〔12〕。持续静脉滴注聚乙二醇-过氧化氢酶能够阻止大鼠石棉沉滞症诱导的PF发生。
1.3.2谷胱甘肽(GSH) GSH是正常肺内的小分子抗氧化剂,PF患者肺内GSH含量减低。谷胱甘肽前体N-乙酰半胱氨酸(NAC)可以增加PF动物模型肺内的GSH水平,减弱博来霉素诱导的PF的发生。雾化吸入NAC,能够增强PF患者抗氧化/氧化平衡体系。一项3期临床实验表明,NAC显示出对PF的潜在治疗价值〔11〕。
2 Nrf2与PF的治疗
2.1Nrf2与PF 机体细胞本身具有一套复杂的抗氧化系统,此系统组成Nrf2-抗氧化反应元件(ARE)-抗氧化酶通路。生理状态下,在胞质内的Nrf2与细胞骨架相关性抑制蛋白(Keap1)结合形成一个复合体,使Nrf2经泛素蛋白酶途径降解。当细胞或机体暴露于ROS时,引起Keap1的修饰,从而使Keap1募集Nrf2的功能发生改变,Keap1-Nrf2复合体解离,Nrf2从胞质转位到细胞核,与受其调控的抗氧化酶基因的ARE结合,从而启动编码解毒和抗氧化的基因表达,使其表达上调,促进细胞存活等功能。见图1。
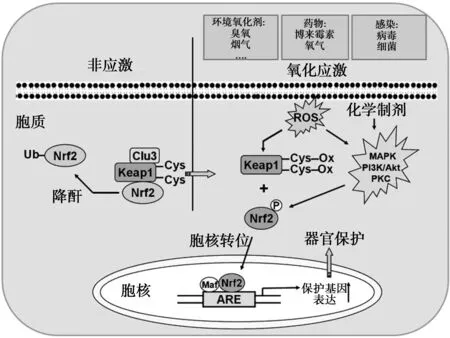
图1 Nrf2抗氧化系统
Nrf2在调节PF过程中的作用引起研究者的关注。Cho等〔11〕是最早研究Nrf2在PF中的作用,研究显示,Nrf2基因敲除(Nrf2-/-)的小鼠中,博来霉素诱导小鼠肺损伤和内皮细胞死亡较野生型小鼠(Nrf2+/+)加重。而反应PF化的指数,即羟脯氨酸含量、胶原沉积、纤维化评分,细胞增殖程度,Nrf2基因敲除(Nrf2-/-)的小鼠较野生型小鼠(Nrf2+/+)水平高。博来霉素诱导野生型小鼠(Nrf2+/+)Nrf2的表达和活性升高。博来霉素可以在野生型小鼠(Nrf2+/+)上调一些受Nrf2调控的细胞保护蛋白的表达。同时,与Nrf2基因敲除(Nrf2-/-)的小鼠比较,在野生型小鼠(Nrf2+/+)博来霉素诱导的反应肺损伤和PF的标志物的蛋白水平较低。综上研究表明,Nrf2在PF的发生发展过程中起到抑制PF发生的保护作用,这一保护作用通过上调特定细胞保护蛋白实现。因此,Nrf2成为治疗PF的新的潜在靶点。最近的一些研究,进一步证实了这一推断,在Nrf2基因敲除(Nrf2-/-)的小鼠中对博来霉素和百草枯(paraquat,PQ)诱导的PF发生更加敏感〔13〕。博来霉素可以通过调控ERK1/2进而抑制Nrf2活性,从而促进氧化应激的产生和PF的发生〔14〕。
2.2激活Nrf2在PF中的保护作用 激活Nrf2抗氧化通路在PF中具有保护作用,使其成为治疗PF的新靶点。以Nrf2抗氧化通路为靶点治疗PF方面,研究者做了有益的探索,取得了一些进展。
2.2.1抑制博来霉素诱导的PF发生 Sriram等〔15〕较早提供了调控Nrf2在PF中具有保护作用的研究证据,研究显示,表没食子儿茶素(EGCG) 可以抑制博来霉素诱导的PF发生,博来霉素可诱导大鼠肺内ROS产生、脂质过氧化作用、羟脯氨酸和髓过氧化物酶的活性增加,EGCG可以减弱这一作用,并恢复博来霉素引起的抗氧化状态。免疫组化分析发现,博来霉素可导致大鼠肺炎症反应增加和肺泡损伤,而EGCG可以降低博来霉素引起的胶原沉积。进一步分析发现,EGCG能够上调博来霉素引起的细胞保护酶基因的表达降低,即上调谷胱甘肽-S-转移酶(GST)和依赖还原型辅酶Ⅰ/Ⅱ醌氧化还原酶 1(NQO1)的表达。同时研究显示EGCG是Nrf2的激活剂。综上所述,EGCG通过激活Nrf2,进而促进GST和NQO1的表达来抑制博来霉素引起的PF发生。植物源性多酚类黄酮类抗氧化剂槲皮素能够通过激活Nrf2,进而上调血红素加氧酶(HO)-1的表达来抑制TGF-β诱导的胶原蛋白的产生,进而抑制PF的发生〔16〕。在雄性Wistar大鼠,黄连素能够抑制博来霉素引起的肥大细胞沉积和组胺的释放,并通过激活Nrf2来增加肺内抗氧化状态,抑制核转录因子(NF)-κB依赖的TGF-β通路激活,进而抑制PF的发生〔17〕。另外最近的研究显示,葫芦巴种子提取物苷〔18〕、丹酚酸〔19〕、南蛇藤醇〔20〕、利拉利汀〔21〕、 中华猕猴桃果仁非饱和脂肪酸〔22〕、苦参碱〔23〕、萝卜硫素〔24〕、广谱抗纤维化作用药物吡非尼酮(PFD)〔25〕均能够不同程度激活Nrf2通路,上调细胞保护蛋白的表达,抑制氧化应激的发生,进而抑制博来霉素引起的PF发生。
2.2.2抑制百草枯诱导的PF发生 诺奥奶蓟草提取物复合胶囊能够在SD大鼠和A549细胞中通过激活Nrf2,进而促进HO-1和NQO1的表达,减弱炎症和氧化应激发生,从而抑制PQ引起的PF发生〔26〕。研究显示葛根素可以抑制PQ诱导的小鼠PF的发生,进一步研究表明,葛根素能够通过激活Nrf2,进而阻断Fstl 1通路、TGF-β1通路和氧化应激发生进而抑制PF的发生〔27〕。最近研究表明,丹酚酸B能够同过激活Nrf2,降低NOX4-NADPH 氧化酶水平,从而减弱PQ引起的小鼠肺内氧化应激的发生,并抑制TGF-β1的表达及其下游蛋白Smad3的磷酸化,最终抑制PQ引起的小鼠PF发生〔28〕。雷怕霉素同样也能够通过激活Nrf2通路抑制PQ引起的PF发生〔29〕。治疗疟疾急性发作的氯喹同样也可经过Nrf2通路的激活,上调HO-1和NQO1的表达,抑制氧化应激的发生,从而能抑制PQ引起的PF发生〔30〕。综上所述,激活Nrf2通路了,能够抑制百草枯诱导的PF发生。
2.2.3抑制硅诱导的PF发生 硅肺病是世界范围内最常见的职业性肺疾病。二氧化硅(SiO2)能够通过诱导大鼠肺内ROS和丙二醛(MDA)产生增加,激活Nrf2及其下游硫氧还蛋白(Trx)。硅抑制Trx活性并下调硫氧还蛋白还原酶(TrxR)的表达,表明硅抑制Trx系统。所以硅通过抑制Trx系统,诱导氧化应激发生,进而诱导大鼠肺PF的发生〔31〕。
最近研究显示,蚯蚓提取物能够通过激活Nrf2依赖的机制抑制二氧化硅诱导的小鼠肺PF的发生〔32〕。蚯蚓提取物可以抑制二氧化硅引起的氧化应激、线粒体依赖的凋亡发生和上皮细胞-间充质细胞转换(EMT)。因此,蚯蚓提取物是潜在的通过激活Nrf2而在PF治疗具有前景的药物。
2.2.4抑制烟气诱导的PF发生 最近研究显示,硫化氢(H2S)能够通过激活Nrf2,进而上调HO-1和硫氧还蛋白(Trx)-1的表达来抑制慢性烟气暴露氧化应激的发生和炎症的产生,进而抑制大鼠肺PF的发生〔33〕。H2S能够减少慢性烟气暴露大鼠肺内氧化应激水平,减少超敏C反应蛋白、TNF-α、IL-1β和IL-6。
3 问题与展望
目前研究表明,Nrf2在抑制肺纤维化过程中具有保护作用,但Nrf2依赖的下游效应因子与PF之间的相互作用仍阐释不清。Nrf2及受其调控蛋白在PF中作用的进一步阐明,对以Nrf2为靶点筛选治疗PF的药物至关重要,对于PF的预防和治疗具有深远意义。
1Faner R,Rojas M,Macnee W,etal.Abnormal lung aging in chronic obstructive pulmonary disease and idiopathic pulmonary fibrosis〔J〕.Am J Respir Crit Care Med,2012;186(4):306-13.
2Galluzzi L,Kepp O,Trojel-Hansen C,etal.Non-apoptotic functions of apoptosis-regulatory proteins〔J〕.EMBO Rep,2012;13(4):322-30.
3Osborn-Heaford HL,Ryan AJ,Murthy S,etal.Mitochondrial Rac1 GTPase import and electron transfer from cytochrome c are required for pulmonary fibrosis〔J〕.J Biol Chem,2012;287(5):3301-12.
4Mossman BT,Marsh JP,Sesko A,etal.Inhibition of lung injury,inflammation,and interstitial pulmonary fibrosis by polyethylene glycol-conjugated catalase in a rapid inhalation model of asbestosis〔J〕.Am Rev Respir Dis,1990;141(5 Pt 1):1266-71.
5Liu G,Cheresh P,Kamp DW.Molecular basis of asbestos-induced lung disease〔J〕.Annu Rev Pathol,2013;8:161-87.
6王玉敏,王月华,杜冠华,等.靶向调控HO-1/NOX2治疗帕金森病的新进展〔J〕.中国药学杂志,2015;50(23):2024-7.
7Hecker L,Cheng J,Thannickal VJ.Targeting NOX enzymes in pulmonary fibrosis〔J〕.Cell Mol Life Sci,2012;69(14):2365-71.
8Liu RM,Gaston-Pravia PKA.Oxidative stress and glutathione in TGF-beta-mediated fibrogenesis〔J〕.Free Radic Biol Med,2010;48(1):1-15.
9Hecker L,Vittal R,Jones T,etal.NADPH oxidase-4 mediates myofibroblast activation and fibrogenic responses to lung injury〔J〕.Nat Med,2009;15(9):1077-81.
10Carnesecchi S,Deffert C,Donati Y,etal.A key role for NOX4 in epithelial cell death during development of lung fibrosis〔J〕.Antioxid Redox Signal,2011;15(3):607-19.
11Cho HY,Reddy SP,Yamamoto M,etal.The transcription factor NRF2 protects against pulmonary fibrosis〔J〕.FASEB J,2004;18(11):1258-60.
12Kliment CR,Oury TD.Oxidative stress,extracellular matrix targets,and idiopathic pulmonary fibrosis〔J〕.Free Radic Biol Med,2010;49(5):707-17.
13Kikuchi N,Ishii Y,Morishima Y,etal.Nrf2 protects against pulmonary fibrosis by regulating the lung oxidant level and Th1/Th2 balance〔J〕.Respir Res,2010;11:31.
14Liu R,Chen H,Bai H,etal.Suppression of nuclear factor erythroid 2-related factor 2 via extracellular signal-regulated kinase contributes to bleomycin-induced oxidative stress and fibrogenesis〔J〕.Toxicol Lett,2013;220(1):15-25.
15Sriram N,Kalayarasan S,Sudhandiran G.Epigallocatechin-3-gallate augments antioxidant activities and inhibits inflammation during bleomycin-induced experimental pulmonary fibrosis through Nrf2-Keap1 signaling〔J〕.Pulm Pharmacol Ther,2009;22(3):221-36.
16Nakamura T,Matsushima M,Hayashi Y,etal.Attenuation of transforming growth factor-β-stimulated collagen production in fibroblasts by quercetin-induced heme oxygenase-1〔J〕.Am J Respir Cell Mol Biol,2011;44(5):614-20.
17Chitra P,Saiprasad G,Manikandan R,etal.Berberine attenuates bleomycin induced pulmonary toxicity and fibrosis via suppressing NF-κB dependant TGF-β activation:a biphasic experimental study〔J〕.Toxicol Lett,2013;219(2):178-93.
18Kandhare AD,Bodhankar SL,Mohan V,etal.Effect of glycosides based standardized fenugreek seed extract in bleomycin-induced pulmonary fibrosis in rats:Decisive role of Bax,Nrf2,NF-κB,Muc5ac,TNF-α and IL-1β〔J〕.Chem Biol Interact,2015;237(2):151-65.
19Tang H,Gao L,Mao J,etal.Salidroside protects against bleomycin-induced pulmonary fibrosis:activation of Nrf2-antioxidant signaling,and inhibition of NF-κB and TGF-β1/Smad-2/-3 pathways〔J〕.Cell Stress Chaperones,2016;21(2):239-49.
20Divya T,Dineshbabu V,Soumyakrishnan S,etal.Celastrol enhances Nrf2 mediated antioxidant enzymes and exhibits anti-fibrotic effect through regulation of collagen production against bleomycin-induced pulmonary fibrosis〔J〕.Chem Biol Interact,2016;246(1):52-62.
21Kabel AM,Abd-Elmaboud MA,Atef A,etal.Ameliorative potential of linagliptin and/or calcipotriol on bleomycin-induced lung fibrosis:In vivo and in vitro study〔J〕.Environ Toxicol Pharmacol,2017;50(2):216-26.
22刘理静,钱 红,尹辉明,等.中华猕猴桃果仁非饱和脂肪酸通过激活Keap1/Nrf2信号通路增强肺纤维化大鼠抗氧化能力〔J〕.细胞与分子免疫学杂志,2016;32(4):479-83.
23罗庆凯,何振华,张秀峰,等.苦参碱对博莱霉素诱导的肺纤维化大鼠中Nrf2表达的影响〔J〕.中国现代医学杂志,2012;22(13):16-20.
24韩 丽,江 涛.萝卜硫素激活Nrf2/ARE通路对博来霉素致大鼠肺纤维化的作用〔J〕.中国新药与临床杂志,2016;35(12):899-905.
25Liu Y,Lu F,Kang L,etal.Pirfenidone attenuates bleomycin-induced pulmonary fibrosis in mice by regulating Nrf2/Bach1 equilibrium〔J〕.BMC Pulm Med,2017;17(1):63.
26Zhao F,Shi D,Li T,etal.Silymarin attenuates paraquat-induced lung injury via Nrf2-mediated pathway in vivo and in vitro〔J〕.Clin Exp Pharmacol Physiol,2015.
27Liu MW,Liu R,Wu HY,etal.Radix puerariae extracts ameliorate paraquat-induced pulmonary fibrosis by attenuating follistatin-like 1 and nuclear factor erythroid 2p45-related factor-2 signalling pathways through downregulation of miRNA-21 expression〔J〕.BMC Comp Alte Med,2016;16:11.
28Liu B,Cao B,Zhang D,etal.Salvianolic acid B protects against paraquat-induced pulmonary injury by mediating Nrf2/Nox4 redox balance and TGF-β1/Smad3 signaling〔J〕.Toxicol Appl Pharmacol,2016;309:111-20.
29Xu Y,Tai W,Qu X,etal.Rapamycin protects against paraquat-induced pulmonary fibrosis:Activation of Nrf2 signaling pathway〔J〕.Biochem Biophys Res Commun,2017;490(2):535-40.
30Shen H,Wu N,Wang Y,etal.Chloroquine attenuates paraquat-induced lung injury in mice by altering inflammation,oxidative stress and fibrosis〔J〕.Int Immunopharmacol,2017;46:16-22.
31Zhu Z,Yang G,Wang Y,etal.Suppression of thioredoxin system contributes to silica-induced oxidative stress and pulmonary fibrogenesis in rats〔J〕.Toxicol Lett,2013;222(3):289-94.
32Yang J,Wang T,Li Y,etal.Earthworm extract attenuates silica-induced pulmonary fibrosis through Nrf2-dependent mechanisms〔J〕.Lab Invest,2016;96(12):1279-300.
33Zhou X,An G,Chen J.Inhibitory effects of hydrogen sulphide on pulmonary fibrosis in smoking rats via attenuation of oxidative stress and inflammation〔J〕.J Cell Mol Med,2014;18(6):1098-103.
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
昆明医科大学学报(2020年12期)2021-01-26
食品与生物技术学报(2021年4期)2021-01-17
世界科学技术-中医药现代化(2020年2期)2020-07-25
中成药(2018年10期)2018-10-26
天然产物研究与开发(2018年8期)2018-09-10
中成药(2018年6期)2018-07-11
中成药(2018年5期)2018-06-06
