
四种变温层积对老山芹种子胚发育及生理特性的影响
2018-06-11 02:24李富恒张雪霞赵恒田吴旭飞吴晶晶刘玉美张晓雯
东北农业大学学报 2018年5期
李富恒,张雪霞,赵恒田,于 萍,吴旭飞,吴晶晶,刘玉美,张晓雯
(1.东北农业大学生命科学学院,哈尔滨 150030;2.中国科学院东北地理与农业生态研究所,哈尔滨 150081)
老山芹(Heracleum moellendorffii Hance)是伞形科牛防风属多年生宿根草本植物,学名东北牛防风,又名山芹菜、短果回芹和大叶芹等[1]。老山芹为耐阴植物,多分布于针阔叶混交林、杂木林下及沟谷湿地等处,是东北林区主要山野菜品种之一。老山芹富含维生素C、核黄素和氨基酸等营养物质[2],幼嫩茎叶可食,全株可作为优质饲料。老山芹可入药,具有医疗保健价值,帮助胃肠蠕动、清理肠道垃圾,治疗风湿性关节炎、腰膝酸痛、头痛、高血压、高血糖及高血脂等疾病[3]。
作为野生植物,老山芹种子具有休眠特性,生产上存在种子发芽率低、催芽难度大、出苗不整齐、育苗技术不成熟、育苗周期过长等问题,限制老山芹生产规模扩大与发展。在老山芹需求量不断增加、野生资源日益枯竭情况下,开展老山芹种子休眠机理及打破休眠技术研究十分必要,可保护林区生态环境、发展林下经济、实现老山芹大面积人工栽培。
种子休眠原因复杂,其中胚后熟是一个重要原因。胚后熟又可分为三种类型:形态后熟型、生理后熟型及复合后熟型。银杏形态后熟型种子采收时外部形态已近成熟[4],但胚尚未分化完全,仍需从胚乳中吸收养料,继续分化发育,直至完全成熟才能发芽;樱桃、山楂、梨、苹果等果树种子属生理后熟型[5],采收时胚外部形态虽已具备成熟特征,但生理上必须通过后熟过程,在种子内部完成一系列生理生化变化后才能萌发;许多种子休眠兼具形态后熟及生理后熟,是一种复合后熟类型,如短梗五加[6]、山茱萸[7]、西洋参[8]、华重楼[9]、东北红豆杉[10]、北五味子[11]、三七[12]、大叶白蜡[13]等。目前老山芹研究多局限于栽培技术[14]、植株及种子生长发育特性[15]、层积条件下种子形态变化[16]等方面,缺乏胚后熟及种子休眠机理深入研究。因此,有必要开展老山芹种子形态及生理后熟综合研究。
本研究以老山芹种子为材料,共设四种不同变温层积处理,定期取样,制作装片观察种胚发育情况并显微摄影,测定种子形态及生理生化指标,分析两类指标动态变化规律及其与种子后熟关系,为揭示老山芹种子休眠机理、打破种子休眠、发展老山芹产业、调整林区经济结构提供理论依据和技术支撑。
1 材料与方法
1.1 材料
试验材料为五年生老山芹植株收获种子,2016年8月采于中国科学院东北地理与农业生态研究所(哈尔滨市)试验田,自然晾干备用。
1.2 方法
1.2.1 种子层积处理及取样
清水冲洗细砂并除去杂质,置于100℃烘箱高温灭菌3 h,取出晾至室温,加入适量蒸馏水搅拌至细砂可握成团但不滴水为止。选籽粒饱满种子,75%乙醇消毒30 s后再置于蒸馏水中浸种6 h。将处理好种子与细砂按照2∶3混匀后放入盛有湿沙塑料盒(长×宽×高,19 cm×13 cm×11 cm),盖子打小孔以利空气交换,按照以下四种方法变温层积处理:
① 20℃,20 d→4℃,30 d→-20℃,10 d→4℃,30 d→20℃,15 d
② 20℃,15 d→4℃,35 d→-20℃,10 d→4℃,35 d→20℃,10 d
③ 20℃,10 d→4℃,90 d→20℃,5 d
④ 20℃,25 d→4℃,35 d→-20℃,5 d→4℃,35 d→20℃,5 d
层积过程中第0、15、30、45、60、75、90、105天分别取样,每隔10 d清洗一次种子和细砂。
1.2.2 种胚形态观察及指标测定
将不同层积处理每隔15 d取样洗净,挑选籽粒饱满、尺寸相近种子去除种被,游标卡尺测定胚乳长。解剖刀剥离种胚制作临时装片,显微镜下用直线式测微尺测定胚长及显微摄影。重复测量10次,取平均值,计算胚率。测量标准如下:
胚乳长:游标卡尺测量胚乳长度最大值;
胚长:直线测微尺测量种胚长度的最大值;
胚率:胚率(%)=胚长/胚乳长×100%。
1.2.3 种子生理生化指标测定
可溶性糖、淀粉含量采用蒽酮法测定,可溶性蛋白质含量采用考马斯亮蓝G250法测定,粗脂肪含量采用索氏提取法测定[17]。
1.2.4 数据分析
运用Excel 2003绘图,DPS7.05统计分析软件处理与分析数据。
2 结果与分析
2.1 不同变温层积处理对老山芹种胚发育进程影响
老山芹种子采收时大部分种子种胚处于心形胚后期阶段(见图1a),层积过程中,由于后熟作用种胚逐渐生长发育,但四种变温层积处理条件下种胚发育进程差异较大。
层积15 d时除处理三有5 d为4℃低温外,其他三个处理均为20℃暖温层积条件,四个处理种胚形态和体积均未发生显著变化,与未处理时基本一致。
层积30 d时观察种胚有进一步发育迹象,处理三进入鱼雷形胚时期(见图1b),其他处理种胚依然处于心形胚后期。
层积45 d时,各处理种胚形态均已发生明显变化,处理一和处理二种胚从心形胚后期开始向鱼雷形胚时期发育;此时处理三在4℃低温下已持续35 d,种胚发育进程较快,已达到鱼雷形胚后期(见图1c),子叶伸长较明显;而处理四种胚发育较为迟缓,未进入鱼雷形胚时期。处理一、处理二、处理三和处理四前期20℃暖温层积时间分别是20、15、10和25 d,随后转入4℃低温层积分别持续25、30、35和20 d,其中处理一、处理二和处理三均在45 d时使种胚发育成鱼雷形胚,而处理四种胚发育较慢,说明暖温层积影响种胚形态后熟,但时间不宜过长,要及时转入低温层积阶段。
处理一和处理二层积60 d时种胚进入鱼雷形胚后期,处理三种胚两片子叶继续伸长生长,而处理四种胚方发育至鱼雷形胚时期。
层积75 d时,处理一和处理二种胚形态和体积与60 d时相比变化小,与经历10 d-20℃冷冻处理有关,冷冻抑制种胚代谢活动,需一定时间才可恢复发育进程;此时可观察到处理三种胚进入子叶期胚阶段(见图1d),两片子叶开始变宽变长,但子叶脉序较模糊;处理四种胚发育至鱼雷形胚后期,较处理一和处理二晚15 d,原因可能与该处理前期20℃暖温层积时间过长(25 d)有关。
层积90 d时,处理一、处理二和处理四种胚子叶吸收胚乳营养物质继续生长,处理三种胚子叶继续变大,子叶脉序已清晰可见(见图1e)。
处理一、处理二和处理四层积105 d时,种胚两片子叶分化,进入子叶期胚阶段,但子叶上脉序较模糊,比处理三晚近30 d;此时处理三种胚子叶较大、脉序较清晰(见图1f)。
层积结束时四种变温层积处理4℃低温下分别处理60、70、90和70 d,表明4℃低温层积阶段持续时间对种胚发育进程形态影响较大,时间越长越有利于种胚发育。比较四种变温层积处理,层积方法处理三模式更利于老山芹种胚形态后熟完成,是一种经济有效的种子处理方法。
2.2 不同变温层积处理条件下老山芹种胚生长动态
2.2.1 胚长
随层积时间延长,各处理种胚长度均不断增长,但每个处理胚长动态变化过程因温度和时间控制不同,表现出较大差异(见图2)。四种变温层积过程中种胚长度与层积时间关系分别用回归方程 y=0.0034x+0.5408、 y=0.0036x+0.5453、 y=0.0053x+0.5133和y=0.0034x+0.54表示。
层积15 d时,各处理胚长从层积前0.51 mm增长至0.58~0.61 mm,增长13.7%~19.6%,各处理种胚长度相差小,未达显著水平,其中处理三胚长较其他处理略低(0.58 mm),可能与处理三为10 d 20℃暖温层积加5 d 4℃低温层积,而其他处理均为15 d 20℃暖温层积有关。
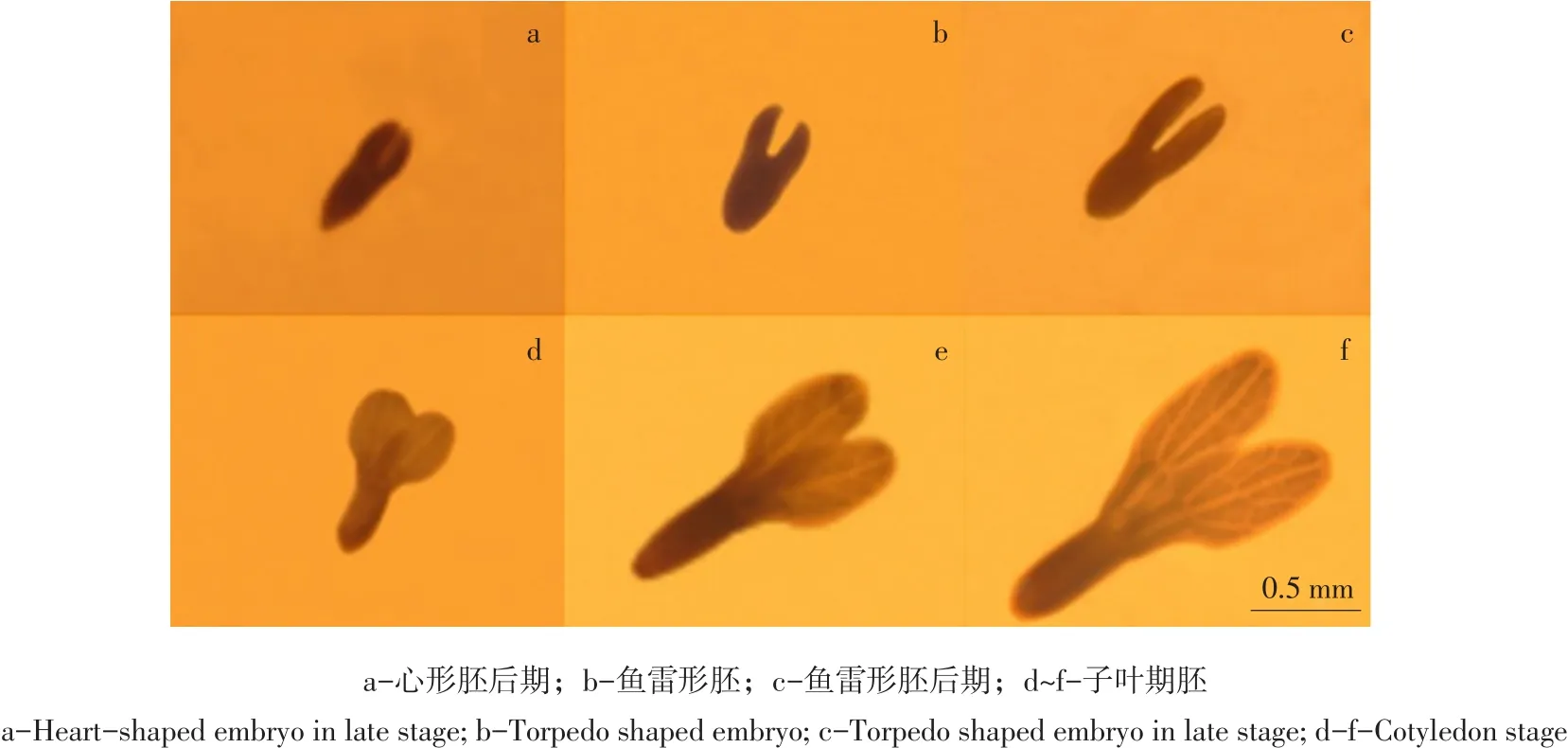
图1 变温层积过程中老山芹种胚发育进程Fig.1 Development of embryo in Heracleum moellendorffii Hance seeds during the process of variable temperature stratification

图2 不同变温层积条件下胚长动态变化Fig.2 Dynamic changes of embryo length under different variable temperature stratification conditions
层积30 d时,各处理胚长分别为0.65、0.68、0.69和0.65 mm,差异不显著;与层积15 d相比,各处理种胚长度分别增长0.05、0.08、0.11和0.04 mm,增长8.3%、13.3%、19.0%和6.6%,处理三种胚增长速率变快。
层积45 d时,各处理种胚长度比层积30 d时分别增加0.04、0.05、0.08和0.06 mm,处理三种胚仍保持较快生长速度,其种胚长度显著高于其他处理(P<0.05),而其他处理间种胚长度无显著差异。层积50 d后将处理一和处理二分别转入-20℃条件放置10 d,而处理三和处理四保持4℃低温层积条件。
层积60 d时,各处理种胚长度较层积45 d时分别增长0.02、0.02、0.04和0.03 mm,达0.71、0.75、0.81和0.73 mm。处理三种胚长度极显著高于处理一(P<0.01),显著高于处理四,而与处理二无显著差异。-20℃冷冻10 d处理一和处理二种胚增长速率变慢。
层积75 d时,各处理种胚长度分别为0.79、0.81、0.89和0.79 mm,与层积60 d相比分别增长0.08、0.06、0.08和0.06 mm,分别增长11.3%、8.0%、9.9%和8.2%,此时处理一和处理二离开-20℃冷冻条件,恢复代谢种胚增长速度加快,从层积第10天起一直处于4℃低温,处理三种胚长度极显著高于处理一和处理四,显著高于处理二。
层积90 d时各处理种胚长度比层积75 d时分别增 加 0.07、0.07、0.05 和 0.04 mm,达 0.86、0.88、0.94和0.83 mm,分别增长8.9%、8.6%、5.6%和5.1%,处理一和处理二种胚增长速率较快,处理三种胚长度极显著高于处理四,显著高于处理一和处理二;而处理一、处理二和处理四之间均差异不显著。
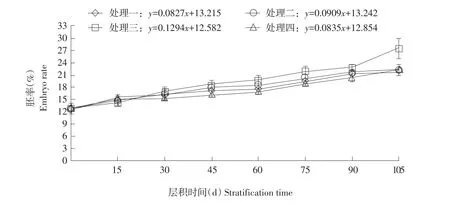
图3 不同变温层积条件下胚率动态变化Fig.3 Dynamic changes of embryo rate under different variable temperature stratification conditions
层积105 d时,各处理种胚长度较90 d时增长0.03、0.02、0.08和0.09 mm,分别增长3.5%、2.3%、8.5%和10.8%;处理一和处理二种胚增长速率变慢,可能与两个处理分别提前10 d和5 d转入20℃暖温层积条件有关。
经105 d变温层积处理,各处理种胚长度由层积前0.51 mm分别增加0.38、0.40、0.61和0.41 mm,达0.89、0.91、1.12和0.92 mm,分别增长74.5%、78.4%、119.6%和80.4%。四种处理相比较,处理三层积模式极显著优于其他处理,种胚生长最快。
2.2.2 胚率
由图3可知,层积15 d后种子胚率开始逐渐增大,各处理胚率分别增加2.9%、2.7%、1.5%和2.0%,胚率值分别达15.4%、14.7%、14.2%和15.0%;四种处理相比较,处理三20℃暖温条件时间最短,胚率值最低,但此时各处理间胚率差异未达显著水平。
随种胚不断伸长,胚率亦逐渐增大,层积至30 d时,胚率分别增加0.7%、1.6%、2.7%和0.3%后达16.1%、16.3%、16.9%和15.3%,此时各处理胚率差异仍未达显著水平。与层积15 d相比,处理三胚率增长最快,其他处理胚率增长速率均下降,可能与三个处理20℃暖温层积条件下时间过长有关。
层积45 d时,与层积30 d相比处理三胚率增长最快,处理二次之,而处理一和处理四胚率增长较慢,各处理胚率分别增加0.8%、1.6%、2.0%和0.8%,胚率值分别达16.9%、17.9%、18.9%和16.1%。处理三胚率值极显著高于处理四,而与处理一和处理二差异不显著。
层积60 d时,与层积45 d相比各处理种子胚率分别增长0.5%、0.5%、1.0%和0.8%后达17.4%、18.4%、19.9%和16.6%。处理三胚率增长速率变慢,其胚率值极显著高于处理一和处理四,而与处理二无显著差异;此期间处理一和处理二经-20℃冷冻10 d层积条件,代谢活动受阻,胚率增长速率减慢,增长率仅为处理三一半。
层积75 d时,各处理种子胚率分别增长2.0%、2.0%、2.0%和1.9%,胚率值分别为19.4%、20.3%、21.9%和18.8%,处理三胚率值极显著高于处理一和处理四,显著高于处理二。处理一和处理二此阶段胚率增长加快,与两个处理冷冻后再次转入4℃低温条件、使种子恢复正常代谢活动有关。此阶段各处理胚率均开始加快增长,说明此阶段是种胚发育重要时间节点,层积过程中要加强管理,促进种胚生长。
层积90 d时,各处理胚率依然保持较高增长速率,分别较前阶段增加1.7%、1.2%、1.1%和1.6%,胚率值分别达21.1%、21.6%、23.0%和20.3%。其中处理三胚率值极显著高于处理四,显著高于处理一,而与处理二差异未达显著水平。
层积105 d时,胚率分别较前期增加0.7%、0.7%、4.4%和2.2%,其中处理三胚率值增加最多,比处理一和处理二高近六倍,比处理四高近两倍;处理一和处理二胚率值增加明显减慢,可能与两个处理分别提前15 d和10 d转入20℃暖温层积条件有关。此时各处理胚率值分别为21.8%、22.3%、27.5%和22.5%,处理三胚率值极显著高于其他处理,而处理一、处理二和处理四之间均差异不显著。
四种不同变温层积处理后种子胚率均有不同程度增加,四种变温层积过程中胚率与层积时间关系分别以回归方程y=0.0827x+13.215、y=0.0909x+13.242、y=0.1294x+12.582和 y=0.0835x+12.854表示。经105 d变温层积处理,各处理胚率值由层积前12.6%分别增加9.1%、9.6%、14.8%和9.9%,达21.8%、22.3%、27.5%和22.5%。四种处理相比较,处理三胚率增加最多,极显著高于其他处理,层积效果最好。
2.3 变温层积过程中老山芹种子生理生化指标动态变化
2.3.1 可溶性糖
由表1可知,与未层积时相比,各处理层积15 d后可溶性糖含量均呈下降趋势,但下降幅度小,各处理间差异不显著。层积30 d时,各处理可溶性糖含量差异提高,呈显著差异,其中处理二极显著高于其他处理;处理一极显著高于处理三和处理四;处理三显著高于处理四。层积过程中胚乳贮藏物质由于代谢活动而分解,与层积15 d时相比较,处理一和处理二可溶性糖含量分别增加3.65和20.11 mg· g-1,升至167.78和185.63 mg· g-1,两个处理胚发育较慢,物质消耗量少,可溶性糖含量小幅升高。而处理三和处理四则分别减少34.51和45.77 mg· g-1,降至119.64和106.39 mg·g-1,此时正是处理三种胚迅速分化发育时期,可溶性糖被大量消耗;而处理四可溶性糖含量下降是20℃暖温层积条件时间过长,种子呼吸消耗物质过多所致。
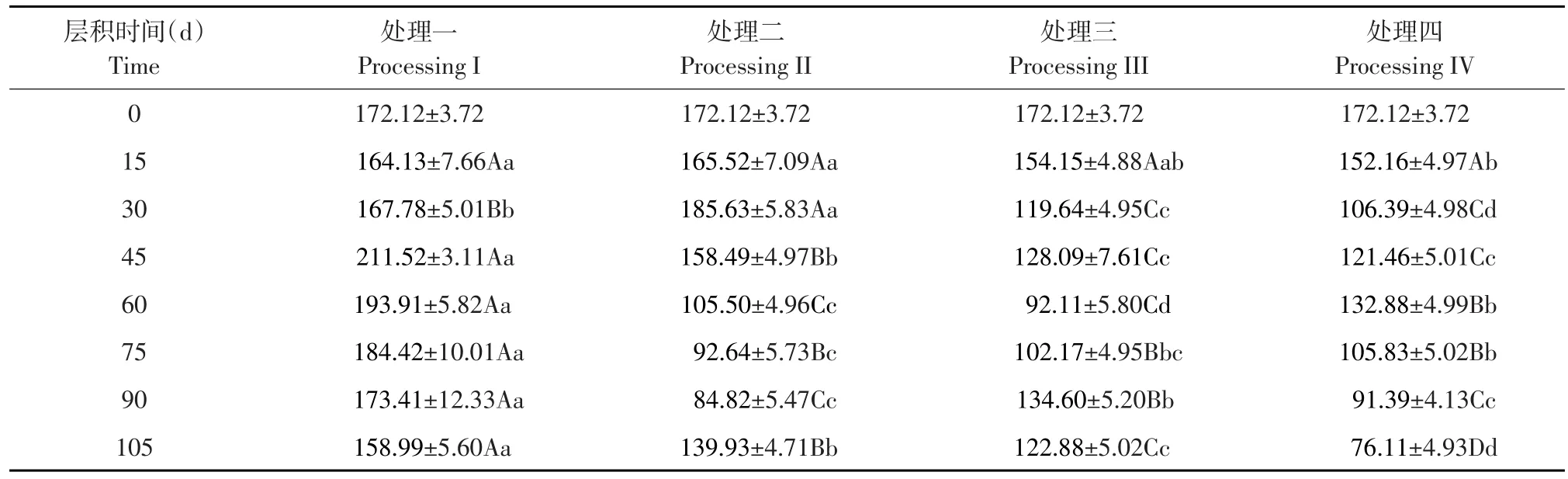
表1 四种变温层积处理过程中可溶性糖含量变化Table 1 Changes of soluble sugar content in four variable temperature stratification processes (mg·g-1)
层积45 d时,四种处理可溶性糖含量分别是211.52、158.49、128.09和121.46 mg· g-1,其中处理一极显著高于其他处理;处理二极显著高于处理三和处理四;而处理三和处理四之间差异不显著。处理一可溶性糖含量较层积30 d时增加43.74 mg·g-1,达到层积期间峰值211.52 mg·g-1;处理二可溶性糖含量减少27.14 mg·g-1;处理三和处理四可溶性糖含量略有上升,分别增加8.45和15.07 mg·g-1。
层积60 d时,各处理可溶性糖含量分别为193.91、105.50、92.11、132.88 mg· g-1,其中处理一极显著高于其他处理;处理四极显著高于处理二和处理三;处理二显著高于处理三;处理一、处理二和处理三此时均呈下降趋势,而处理四则呈上升趋势。处理一和处理二可溶性糖含量下降可能与种子转至-20℃冷冻条件使代谢活动减弱有关;而处理三此时种胚发育较快、消耗营养物质过多,故可溶性糖含量低。
层积75 d时,除处理四在-20℃冷冻5 d后又转至4℃低温继续层积外,其他处理均是4℃低温条件下层积。与层积60 d相比较,处理一、处理二和处理四可溶性糖含量分别下降9.49、12.86和27.05 mg·g-1,而处理三上升10.06 mg·g-1。四个处理相比,其中处理一极显著高于其他处理,其他处理差异小。
层积90 d时,各处理可溶性糖含量分别为173.41、84.82、134.60和91.39 mg· g-1,其中处理三比层积75 d时增加32.43 mg·g-1,其他处理均呈下降趋势,分别减少11.01、7.82和14.44 mg·g-1,可能与此时种胚生长加速,消耗营养物质过多有关。
105 d层积结束时,各处理可溶性糖含量分别为158.99、139.93、122.88和76.11mg· g-1,各处理间均表现极显著差异。处理一、处理三和处理四可溶性糖含量分别减少14.42、11.72和15.28 mg·g-1,而处理二可溶性糖含量增加55.11 mg·g-1。此时处理一和处理四种胚生长加速,消耗营养物质较多,处理三种胚已发育完全。
四种变温层积处理过程中老山芹种子可溶性糖含量总体上呈先降后升再降趋势,但较未处理种子相比均有不同程度下降。各处理出现第一个峰值时间分别为层积后第45、30、45和60天;处理二和处理三第二个峰值则出现在105 d和90 d,而处理一和处理四无第二峰值。除处理三外,其他处理均有一定时间-20℃冷冻条件,且处理三在20℃暖温层积条件下经历时间较短,说明20℃暖温层积条件和-20℃冷冻条件均对可溶性糖含量产生影响,影响种胚发育。
2.3.2 淀粉
由表2可知,与未层积时相比,层积15 d时四种处理淀粉含量略有下降,分别减少9.5%、9.3%、11.0%和9.8%,此时各处理间淀粉含量差异均不显著;其中处理三下降较多,这可能与处理三前15 d中有5 d处于4°C低温使种胚发育较其他处理略快有关。
层积30 d时,淀粉含量依然呈下降趋势,各处理较层积15 d时分别降低9.6%、11.6%、18.5%和6.4%。处理三淀粉含量最低,与其他各处理呈极显著或显著差异。
随4℃低温处理时间延长,淀粉分解速率加快,层积45 d时,各处理淀粉含量较层积30 d时分别下降20.0%、15.2%、21.1%和12.3%。处理四显著高于处理二、极显著高于处理一和处理三,处理一和处理二之间差异不显著,但两个处理极显著高于处理三。
层积60 d时,各处理淀粉含量较层积45 d时分别下降3.6%、4.0%、7.0%和5.6%。处理四淀粉含量极显著高于处理一和处理三,显著高于处理二;处理二和处理三之间差异极显著;处理一和处理三之间差异显著。
层积75 d时,各处理淀粉含量仍呈下降趋势,较层积60 d时分别下降19.1%、21.3%、12.5%和10.26%。处理四淀粉含量极显著高于其他处理,而其他处理间均表现为差异不显著。此时处理一和处理二淀粉含量下降较快,可能与其从-20℃冷冻阶段转入4℃低温阶段,种子代谢活动恢复有关。处理四淀粉含量下降较慢,与此时处理四经历5 d-20℃冷冻阶段有关。
层积90 d时,各处理淀粉含量较层积75 d时分别下降7.7%、10.5%、33.6%和14.8%。处理三极显著低于其他处理;处理四显著高于处理一和处理二;处理一和处理二差异不显著。此时各处理淀粉含量分别减少0.74、1.01、2.98和1.75 mg·g-1,其中处理三下降最快,与此时处理三可溶性糖含量对应。
105 d处理结束时,处理三极显著低于处理一和处理四、显著低于处理二;其他处理间均表现为差异不显著。各处理淀粉含量较层积90 d时分别下降15.6%、18.3%、2.5%和21.6%,其中处理四下降最多、处理三下降最少。说明种胚发育快的处理此时淀粉含量下降少,发育慢淀粉含量下降多,即种胚发育需分解淀粉为可溶性糖作为能量和原料来源。
四种变温层积处理均使老山芹种子淀粉含量呈下降趋势,四种变温层积过程中种子淀粉含量与层积时间关系分别以回归方程y=-0.11x+18.197、y=-0.1119x+18.47、y=-0.1279x+18.083和y=-0.0982x+18.705表示。各处理不同时期淀粉含量减少程度相比,处理三在层积90 d下降最快,此时处理三种胚发育较快,子叶上脉序已清晰可见,消耗淀粉量较大。
2.3.3 可溶性蛋白质
由表3可知,与未层积时相比,各处理层积15 d后可溶性蛋白质含量上升,分别较未层积时增加209.96、251.54、179.53和257.83 mg· g-1,其中处理三可溶性蛋白质含量涨幅略小于其他处理,显著低于处理二和处理四。此时层积前期20℃暖温阶段种子代谢活跃,多种蛋白质及酶大量合成,可溶性蛋白质含量上升。
层积30 d时,各处理可溶性蛋白质含量较层积15 d时均出现大幅下降,分别下降18.8%、31.4%、44.0%和30.0%。处理一极显著高于其他处理,处理二和处理四极显著高于处理三,而处理二和处理四差异不显著。其中处理三下降幅度最大,是处理一两倍,与此阶段处理三种胚生长加速、营养物质消耗过多有关。
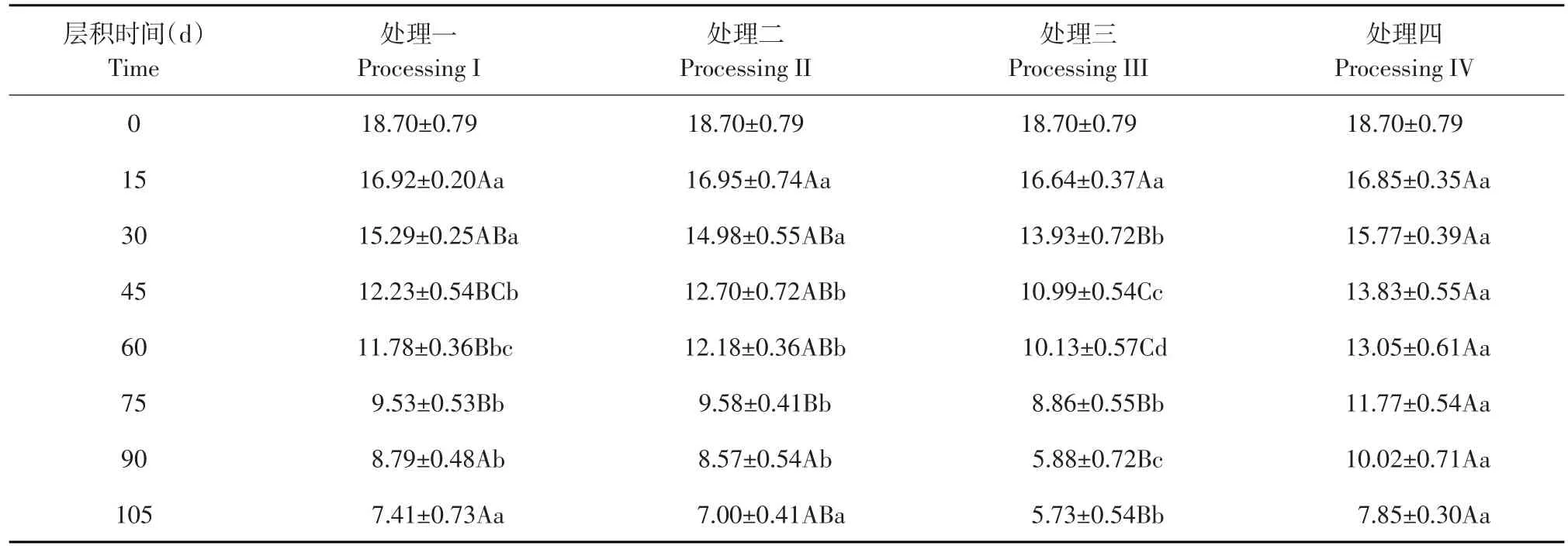
表2 四种变温层积处理过程中淀粉含量变化Table 2 Changes of starch content in four variable temperature stratification processes (mg·g-1)
层积45 d时,各处理可溶性蛋白质含量比层积30 d时分别下降4.1%、2.5%、4.0%和7.6%。此时处理一可溶性蛋白质含量极显著高于其他处理,处理二和处理四极显著高于处理三,而处理二和处理四差异不显著。此阶段较层积30 d时下降幅度明显趋缓。
层积60 d时,各处理间可溶性蛋白质含量表现不一,有升有降,各处理间均表现为极显著差异。此时处理三和处理四可溶性蛋白质含量继续下降,分别减少107.80和12.68 mg·g-1,处理三可溶性蛋白质含量大幅下降可能与种胚此时生长加速、消耗蛋白质过多有关。而处理一和处理二可溶性蛋白质含量上升,分别增加123.52和79.94 mg·g-1,可能与两个处理转至-20℃冷冻阶段使代谢活动减少有关。
层积75 d时,各处理可溶性蛋白质含量分别为538.35、371.63、378.67和601.24 mg· g-1,处理四极显著高于其他处理,处理一极显著高于处理二和处理三,而处理二和处理三间差异不显著。处理一和处理二经15 d 4℃低温阶段后可溶性蛋白质含量分别减少281.86和316.68 mg·g-1,可能与两个处理从-20℃冷冻阶段转至4℃低温阶段使种子恢复代谢、种胚生长加速有关。
层积90 d时,各处理可溶性蛋白质含量分别为667.57、438.88、458.70和246.08 mg· g-1。与层积75 d相比,处理一、处理二和处理三均呈上升趋势,分别升高24.0%、18.0%和21.1%;处理四则呈下降趋势,下降59%。此时处理一极显著高于其他处理;处理二和处理三极显著高于处理四;处理二和处理三之间差异不显著。此期间处理四种胚生长较快,消耗蛋白质过多。
层积105 d时,处理一、处理二和处理三可溶性蛋白质含量较层积90 d时分别下降30.1%、9.7%和19.1%,其中处理一下降幅度最大;而处理四可溶性蛋白质含量则上升25.5%。此时处理一极显著高于其他处理;处理二和处理三极显著高于处理四;处理二和处理三之间差异不显著。
整个层积过程中,各处理可溶性蛋白质含量变化较大,呈先升再降趋势,由未层积时658.67 mg·g-1分别降至层积结束时465.99、395.91、370.78和308.92 mg·g-1。其中处理三有90 d处于4℃低温阶段,其可溶性蛋白质含量在多数取样期较其他处理低,且种胚生长较快,说明种胚生长需消耗较多蛋白质。而处理一和处理二45~60 d时经历一定时间-20℃冷冻阶段,此期间可溶性蛋白质含量大幅上升,与-20℃冷冻阶段蛋白质消耗减少、抗寒性蛋白质大量合成有关。各处理可溶性蛋白质含量大幅下降时间分别为层积后75、75、60和90 d,处理三大幅下降时间最早、种胚发育最快,处理四大幅下降时间最早最迟,种胚发育最慢。说明可溶性蛋白质含量变化与种胚发育进程有关。
2.3.4 粗脂肪
由表4可知,与未层积时相比较,层积15 d后各处理粗脂肪含量变化较小,分别减少0.17%、0.20%、0.35%和0.25%,各处理间粗脂肪含量均无显著差异。
层积30 d时,各处理粗脂肪含量较层积15 d时分别减少0.51%、0.68%、0.86%和0.29%。处理一和处理四显著高于处理三,而处理二和处理三之间差异不显著。此时粗脂肪含量下降速度加快,其中处理三下降幅度最大,其次为处理二和处理一,处理四下降幅度最小。
层积45 d时,种子粗脂肪含量呈下降趋势,各处理较层积30 d时分别减少0.55%、0.41%、0.73%和0.42%。处理一、处理二和处理四之间差异不显著;处理一和处理四极显著高于处理三;处理二显著高于处理三。
层积60 d时,各处理较层积45 d时粗脂肪含量分别减少0.12%、0.14%、0.61%和0.45%。此时处理三粗脂肪含量极显著低于其他处理,其他处理间均表现为差异不显著。处理一和处理二粗脂肪含量下降速率显著低于处理三,原因是两个处理有10 d处于-20℃冷冻条件,种子代谢活动减慢;而处理三和处理四粗脂肪含量下降速率较层积45 d时无明显变化。
层积75 d时,仍表现为处理三粗脂肪含量极显著低于其他处理,其他处理间均表现为差异不显著。各处理粗脂肪含量降低幅度较大,分别降低0.87%、0.87%、0.60%和0.45%。其中处理一和处理二粗脂肪含量下降速率加快,此时已从-20℃冷冻条件转换至4℃低温条件;处理三下降速率保持不变;而处理四下降速率较慢,经历5 d-20°C冷冻条件。

表4 四种变温层积处理过程中粗脂肪含量变化Table 4 Changes of crude fat content in four variable temperature stratification processes (%)
层积90 d时,各处理粗脂肪含量分别减少0.52%、0.79%、0.60%和0.33%。处理三极显著低于其他处理,处理四极显著高于处理二和处理三,而与处理一差异不显著。
105 d层积结束时各处理粗脂肪含量分别较层积90 d时减少0.69%、0.96%、0.83%和1.14%。处理三极显著低于其他处理;处理一和处理四差异不显著;处理一与处理二差异显著。
经变温层积处理后,各处理粗脂肪含量从未层积时8.27%分别下降3.43%、4.05%、4.58%和3.62%,降至4.84%、4.22%、3.69%和4.65%,粗脂肪含量总体呈下降趋势。四种变温层积过程中粗脂肪含量与层积时间关系分别以回归方程:y=-0.0328x+8.5126、y=-0.0379x+8.6176、y=-0.0441x+8.4037、y=-0.0326x+8.5489表示。四种处理相比较,处理三粗脂肪含量下降最多,处理二和处理四次之,处理一下降最少。
3 讨论与结论
3.1 老山芹种子休眠类型
种子休眠是指有生命种子在适宜环境条件下仍不萌发现象[18],是种子为度过外界不良环境而形成的一种适应性生物学特性,对植物个体生存和进化具有重要意义。自然条件下播种老山芹种子,需越冬后第二年春天萌发,部分种子则可能需更长时间,出芽率低、出苗不齐和育苗周期长等问题限制老山芹产业发展。研究老山芹种子休眠机理具有重要意义。
种子休眠类型多且复杂,Baskin等将种子休眠分为物理休眠、形态休眠、生理休眠、形态生理休眠和复合休眠五种类型[19]。种子具有坚硬厚实外壳使透气透水性变差,引起休眠为物理休眠[20]。刘增兵对老山芹种子作吸水试验,结果表明老山芹种被对种子吸水影响较小,不会构成种子萌发障碍[21]。说明老山芹种子不存在物理休眠,是种子内部因素引起休眠。种子脱离母体后胚未分化或胚已分化但未发育完全引起休眠称为形态休眠。本研究结果表明,老山芹种子采收时种胚较小,胚率仅为12.6%,种胚发育程度不一致,但大多数种胚处于心形胚后期阶段,层积过程中不断分化发育,胚率逐渐增大,最后发育至作为成熟胚标志子叶期胚阶段,具备发芽能力,说明老山芹种子存在形态休眠。
生理休眠是指种子脱离母体后虽然种胚已发育完全,但还不具备萌发能力,需经一段时间完成生理后熟积累种子萌发所需营养物质后才有萌发可能。形态生理休眠是指植物种子同时具备形态休眠和生理休眠生物学特性[22];复合休眠是指植物种子同时具备物理和生理休眠生物学特性[23]。本研究发现,层积处理结束后虽然老山芹种胚已发育到子叶期胚阶段,种胚已发育完全,但催芽过程中种子萌发率仍较低。刘增兵研究表明老山芹种子内存在大量抑制物质[21]。因此,可初步认为老山芹种子是形态生理休眠类型。但其生理休眠具体机制尚不清楚,有待深入研究。
3.2 形态生理休眠类型种子打破休眠措施
张恩和等采用变温(15~25℃/2~5℃)层积处理野生羌活种子,暖温层积促进种胚分化发育完成形态后熟,低温层积促进种子生理后熟完成[24];何利平对刺楸气干种子的研究表明,5~15℃沙藏110 d种子完成形态后熟后,需继续沙藏,种子完成生理后熟萌发[25];黄玮等研究结果表明,经180 d变温层积处理,滇重楼种子虽已完成胚形态后熟,但种子仍不萌发,需再继续低温处理,待种子完成生理后熟才具备发芽能力[26];葛超等对短梗五加种子研究表明,种子萌发不受种皮阻碍影响,需先暖温层积条件下使种胚分化生长,再转入低温层积处理完成生理后熟促进种子萌发[6]。本研究设计四种变温层积处理探讨打破老山芹种子休眠方法,变温层积过程中4℃低温持续时间对种胚发育进程影响最大,时间越长越有利于种胚发育;四种变温层积处理相比较,处理三模式(20℃,10 d→4℃,90 d→20℃,5 d)更利于老山芹种胚形态后熟完成,最早进入子叶期胚,胚长和胚率增长最快,是一种经济有效的种子处理方法。未来需深入研究,高效打破种子休眠。
3.3 老山芹种子胚形态后熟过程与营养物质变化关系
种子后熟是代谢过程,需提供原料和能量,伴随复杂的代谢活动[27]。可溶性糖为种子代谢活动提供能量基础物质,用于消耗;淀粉、蛋白质和粗脂肪是种子中主要贮藏物质,种子后熟和萌发过程中会被分解为小分子物质,为种胚发育提供原料和能量。具有形态生理休眠类型的北五味子[11]、南方红豆杉[28]和秤锤树[29]等种子,层积处理过程中种胚分化发育与伸长生长同时伴随营养物质分解、转化及利用。
种子层积期间淀粉及粗脂肪被分解,继而转化成可溶性糖及其他小分子物质为种胚生长发育利用,可溶性蛋白质会降解成氨基酸,形成不含氮化合物和游离氨,其中不含氮化合物会转化成糖类为种子呼吸代谢提供能量,游离氨会在可溶性糖积累达到一定阈值时,重新形成氨基酸,合成蛋白质,蛋白质与种胚形态分化发育的结构蛋白及某些酶类有关[29]。本研究结果表明,层积处理前期,老山芹种子可溶性糖、淀粉及粗脂肪含量下降,可溶性蛋白质含量上升,与东北红豆杉[10]、北五味子[11]和三七[12]在层积前期种子生理生化变化研究结果一致。随层积时间延长,淀粉和脂肪被不断分解,可溶性蛋白质分解速率加快、含量下降,其中处理一、处理二和处理四种胚生长较慢,可溶性糖消耗较少,含量增加出现峰值,之后种胚生长速率加快,可溶性糖出现下降趋势。处理三种子种胚生长速率较其他处理快,整个层积过程种子可溶性糖消耗速率较高,因此可溶性糖一直呈下降趋势。当处理一、处理二和处理四进入-20℃冷冻阶段时,种子呼吸代谢活动变慢,种胚生长速率减小,因此淀粉和粗脂肪含量下降缓慢,可溶性蛋白质含量略有上升。冷冻解除后,代谢活动和种胚生长逐渐恢复正常,但由于水解酶被-20℃钝化,需逐步提高活性,可溶性糖、淀粉、可溶性蛋白和粗脂肪含量均呈下降趋势。层积后期处理三种胚发育至子叶期胚阶段,形态后熟已基本完成,物质消耗减少,可溶性蛋白质和可溶性糖含量升高。
本研究表明,处理三(20℃,10d→4℃,90d→20℃,5 d)较其他处理种胚分化发育速度快,且可溶性蛋白质、淀粉及粗脂肪转化和利用均早于其他处理。说明-20℃冷冻阶段并非必需,采用暖温-低温-暖温变温层积方法有利于老山芹种子后熟,但暖温层积时间不宜过长。
[1] 中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,2004.
[2] 刁绍起,朱楠楠,孙广仁.山芹菜营养成分分析与加工特性[J].东北林业大学学报,2010,38(10):48-50.
[3] 卢伟,付强,贾楠,等.短果回芹化学成分的研究[J].吉林医药学院学报,2015,36(6):480-481.
[4] 芮海云.银杏种子发育和后熟生理研究[J].廊坊师范学院学报:自然科学版,2009,9(3):65-68.
[5] 张秋香,武绍波,杨荣萍,等.果树种子休眠原因及解除休眠的方法[J].山西果树,2004(1):31-33.
[6] 葛超,宁伟,张立军,等.短梗五加种子萌发影响因子研究[J].种子,2007,26(11):6-9.
[7] 洑香香,周晓东,刘红娜.山茱萸种子休眠机理与解除方法初探[J].中南林业科技大学学报,2013,33(4):7-12.
[8] 秦公伟,曹小勇,赵烨,等.植物生长物质对西洋参种子与离体种胚的休眠解除作用[J].陕西理工学院学报:自然科学版,2016,32(3):46-52.
[9] 李昭玲,童凯,闫燊,等.变温层积过程中华重楼种胚后熟生理生化的变化[J].中国中药杂志,2015,40(4):629-633.
[10] 廖云娇,李雪,董学会.不同变温层积过程中东北红豆杉种子生理生化特性和胚形态的变化[J].中国农业大学学报,2010,15(1):39-44.
[11] 夏广清,李冬梅,李彩凤,等.北五味子种子萌发中形态学观察及生理生化指标分析[J].植物生理学报,2014,50(4):415-418.
[12] 段承俐,李章田,金玲,等.三七种子的后熟生理特性研究[J].中国中药杂志,2010,35(20):2652-2656.
[13] 刘润妮,周珊,刘长命.大叶白蜡种子休眠机理及预处理方法的研究[J].山西农业科学,2012(2):106-110.
[14] 信小娟,刘成学,李玉成.大兴安岭山野菜老山芹栽培技术[J].防护林科技,2016(8):120-121.
[15] 李富恒,刘增兵,崔巍金琦,等.老山芹生长发育规律及主要性状相关性分析[J].东北农业大学学报,2017,48(1):15-22.
[16] 李富恒,李楠丁,刘增兵,等.不同层积条件下老山芹种子形态后熟的研究[J].东北农业大学学报,2017,48(5):28-34.
[17] 中国科学院上海植物生理研究所,上海市植物生理学会.现代植物生理学实验指南[M].北京:科学出版社,1999.
[18] 付婷婷,程红焱,宋松泉.种子休眠的研究进展[J].植物学报,2009,44(5):629-641.
[19] Baskin J M,Baskin C C.A classification system for seed dormancy[J].Seed Science Research,2004,14(1):1-16.
[20] 颜启传.种子学[M].北京:中国农业出版社,2001.
[21] 刘增兵.老山芹生物学性状及种子休眠特性的研究[D].哈尔滨:东北农业大学,2017.
[22] 黄丹,许岳香,胡海波.植物种子休眠原因与机理的研究进展[J].亚热带植物科学,2010,39(2):78-83.
[23] 张侠,李进,吕海英,等.准噶尔山楂种壳、种皮、种胚特性与种子休眠的关系[J].植物生理学报,2014,50(6):867-874.
[24] 张恩和,陈小莉,方子森,等.野生羌活种子休眠机理及破除休眠技术研究[J].草地学报,2007,15(6):409-414.
[25] 何利平.刺楸种子休眠原因及解除休眠的研究[J].山西林业科技,2003(4):22-24.
[26] 黄玮,孟繁蕴,张文生,等.滇重楼种子休眠机理研究[J].植物生理科学,2008,24(12):242-246.
[27] 杨国会,孙立梅,邢力,等.不同层积处理方法对东北红豆杉种子后熟过程中贮藏物质的影响[J].北方园艺,2013(14):82-85.
[28] 黄儒珠,郭祥泉,方兴添,等.变温层积处理对南方红豆杉种子生理生化特性的影响[J].福建师范大学学报:自然科学版,2006,22(2):95-98.
[29] 贾书果.秤锤树种实发育的生理特性与种子休眠机理的研究[D].南京:南京林业大学,2008.
猜你喜欢
植物资源与环境学报(2022年6期)2022-12-16
含笑花(2021年4期)2021-08-13
防护林科技(2020年11期)2020-12-30
潍坊学院学报(2020年6期)2020-11-22
中国食品学报(2019年3期)2019-01-13
许昌学院学报(2018年8期)2018-09-05
微型小说选刊(2017年4期)2017-12-06
蔬菜(2016年10期)2016-03-27
武夷学院学报(2015年3期)2015-07-18
云南中医学院学报(2014年3期)2014-07-31
