
7个烤烟产量相关性状的QTL定位分析
2018-09-05 06:27童治军焦芳婵陈学军吴兴富方敦煌肖炳光
西北植物学报 2018年7期
童治军,焦芳婵,陈学军,吴兴富,方敦煌,肖炳光
(云南省烟草农业科学研究院,烟草行业烟草生物技术育种重点实验室,国家烟草基因工程研究中心,昆明 650021)
栽培烟草(NicotianatabacumL.)是烟草属内一年生的异源四倍体草本植物,同时也是一种广泛种植于温带农业区域的重要经济作物[1-5]。作为重要经济作物的烟草,烟叶生产是烟草行业的基础,而烟草品种的培育又是烟叶生产的基础[6-7]。因此,培育具有较高烟叶产量的新品种,是当前烟草育种中最重要的目标之一[8-9]。与其他大田作物一样,烟叶的产量也是属于微效多基因控制且极易受环境影响的数量性状[10-11]。据报道,烟叶产量是与株高(节距)、茎围、叶片数和叶片形状等农艺性状直接相关[12-14]。故此,改善与烟叶产量相关的农艺性状,就成为提高烟叶产量的首选途径[15]。
局限于烟草栽培品种间狭窄的遗传基础和极低的遗传多样性[16-20],使得QTL定位研究相较于其他作物显得比较落后,远未达到育种应用的程度。目前,在烟草的QTL定位研究方面虽然也有一些报道[21-34],但这些研究全部是基于低质量的烟草遗传图谱上进行,因此,极有可能会因错过许多图谱未涵盖基因组区域而导致获得的QTLs数量偏少。此外,这些已报道的研究主要集中在对烟草抗病性的QTL定位分析上,如烟草根黑腐病[21]、马铃薯Y病毒病(PVY)[21]、黑胫病[22-23]、青枯病[21,24]、赤星病[25]、白粉病[26]、番茄斑萎病(TSWV)[27]和黄瓜花叶病(CMV)[28]等的QTL分析,也有少部分是关于烟草化学成分[29-32]及叶片烘烤特性[33]的QTL定位研究。迄今,关于烟草(烤烟)产量性状相关QTL定位分析的研究仅有1篇,即,Tong等[34]利用一个由207个株系构成的DH群体和基于该群体构建获得的SSR标记遗传图谱,对包含株高、节距、叶数等6个与烤烟产量相关农艺性状的QTL定位分析。
上述烟草QTL定位研究主要是基于非特异性标记(如RAPD、ISSR、AFLP、SCAR等)[21,24,27,29,31]构建的局部烟草遗传图谱[21,23-24,26-29,31-33]进行的,因此,在很大程度限制了其在烟草育种中的进一步应用。鉴于此,本研究利用烤烟品种Y3和K326为亲本构建的F6代RILs群体和已构建获得的高质量烟草遗传连锁图谱,并结合连续2年(2016~2017)的田间表型数据,在全基因组范围内对与烤烟产量相关的7个农艺性状进行QTL分析,以期为产量相关性状基因的精细定位、克隆及利用分子标记辅助选择(MAS)方法培育高产烤烟新品种提供重要的理论依据。
1 材料和方法
1.1 实验材料
以烤烟品种K326和Y3为父母本,配制组合并产生F1,从F2世代始通过单粒传法(SSD)获得一个包含262个株系的遗传性状稳定(F6)的重组自交系(RILs)。其中,K326是由美国引进的具有优良品质和广泛种植面积的烤烟品种,而Y3则是由津巴布韦引进的具有优良农艺性状且丰产性好的烤烟种质资源。
RILs群体于2016~2017年种植于云南省烟草农业科学研究院研和试验基地,采用完全随机区组设计,设置3个重复,每个株系种植一个小区,每个小区20株,株行距为0.50 m ×1.0 m,并按当地优质烟生产技术措施进行栽培管理。农艺性状调查按照烟草行业标准YC/T 369-2-10进行,其中包括株高(自然株高)、叶片数(自然叶片数)、节距、茎围、茎叶角度、腰叶长和腰叶宽,具体测量方法如下:
(1)株高:采用杆尺,测量自地表茎基处至第一青果柄基部间的长度。
(2)叶片数:自主茎基部至中心花下第五花枝处着生叶片数。
(3)节距:沿着烟株的主茎从下往上占整个株高的1/3的位置称腰部,在烟株腰部的上下各测量2个相邻叶片的基部间距离,然后取平均值。
(4)茎围:在烟株的腰部测量主茎的周长。
(5)茎叶角度:在主茎腰部,测量叶片与主茎间的夹角。
(6)腰叶长:选择腰部最大叶片,测量茎叶连接处至叶尖的直线长度。
(7)腰叶宽:选择腰部最大叶片,测量与主脉垂直的叶面最宽处长度。
1.2 数据统计与分析
应用 Excel 2013和 DPS v14.10软件,对262个RILs材料在2个年份的7个农艺性状的平均值、标准差、偏度、峰度和相关系数等指标进行整理和分析。
1.3 QTL定位分析
基于已构建获得的高质量烤烟遗传连锁图谱[35],并结合7个目标性状的田间表型数据,利用MapQTL®v 6软件[36]提供的Restricted Multiple-QTL model Mapping (rMQM)方法进行全基因组QTL扫描定位,相关的参数设置如下:算法选用回归算法,测试统计量选择LOD,选择Fn(RILs)为合适的模型,定位时的步长为1.0,相邻标记的最大数目为30,最大迭代次数和排列数均为10 000。按照McCouch等[37]规则进行QTLs命名。
2 结果与分析
2.1 7个性状的表型分析
通过对7个目标性状的田间表型数据进行统计分析,其结果(表1)显示:1)7个与烤烟产量相关的农艺性在双亲间均存在明显的差异,其中,株高(PH)、叶片数(LN)、节距(IL)和腰叶长(LWL)4个性状在双亲间呈极显著差异,且在性状表型值上母本Y3均较父本K326高;2)全部的7个性状在RILs群体内各株系间存在较大范围的连续变异且具有显著的双向超亲分离,同时,各性状的平均值也很接近中亲值;3)各性状的偏度(skewness)和峰度(kurtosis)绝对值均小于1(<1),此表明本研究中所涉及的7个性状在RILs群体中呈正态分布,符合典型的数量性状特征。7个农艺性状的平均广义遗传率为73.33%,其中,株高和节距的广义遗传率在80%以上;茎围和茎叶角度的广义遗传率则低于60%,表明其易受环境影响。上述结果表明,7个与烤烟产量相关的农艺性状是既受微效多基因控制又受环境条件影响的数量性状。
对7个性状田间表型数据的相关性分析表明(表2):每个性状均与其他的2个或2个以上性状存在显著的相关性。如,株高(PH)与叶片数(LN)、节距(IL)和腰叶长(LWL)存在显著的相关性;茎围(SG)与叶片数(LN)、节距(IL)、腰叶长(LWL)和腰叶宽(WWL)存在显著的正相关。除叶片数(LN)与节距(IL)呈显著的负相关外,余下的所有性状间均呈现显著的正相关。7个性状间相关系数达到极显著水平(P<0.01)的有4对,其中,株高(PH)与节距(IL)间的相关系数最大,为0.913,该结果表明本研究中的两亲本及其衍生RILs群体单株的主茎节距越大,则其株高也就越高;腰叶长(LWL)与腰叶宽(WWL)间的相关性也非常高(达0.835),这表明,叶片形状(以LWL/WWL的比例表示)在RILs群体内并没有较大的变化;此外,节距(IL)与叶片数(LN)之间则呈现出极显著的负相关(-0.803),表明在株高一定的前提下,节距越大则植株主茎上着生的叶片数就越少。
注:遗传率为广义遗传率;*和**分别指双亲间差异达0.05和0.01显著水平
Note: Heritability is the heritability in broad sense.*and**mean significant difference between parents at 0.05 and 0.01 levels, respectively
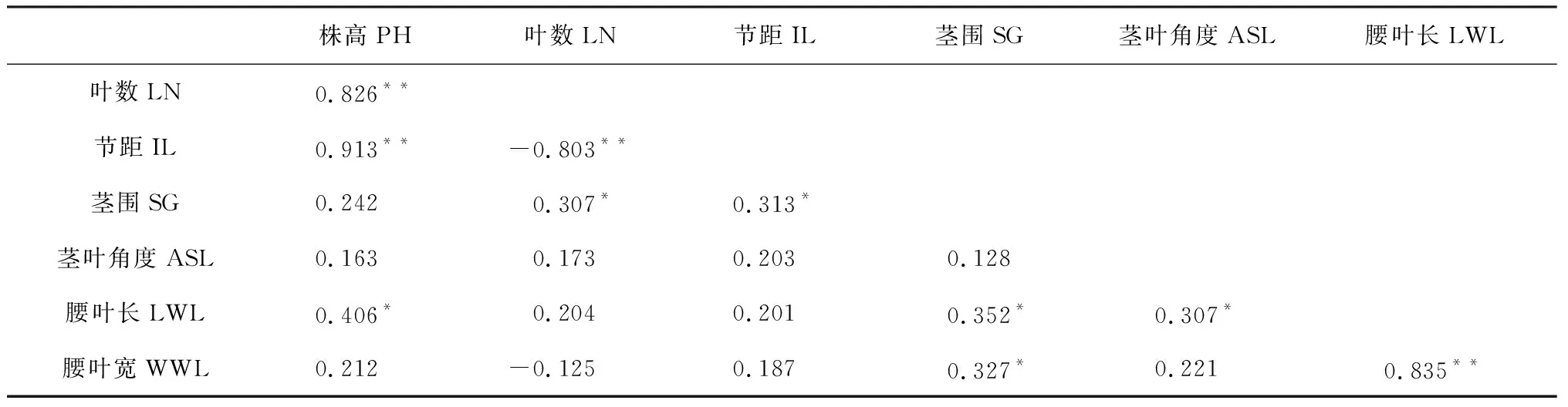
表2 7个产量相关性状在RILs群体内的相关性分析
注:*和**分别表示相关性达到0.05和0.01显著水平
Note:*and**indicate significant correlation at 0.05 and 0.01 level, respectively
2.2 目标性状的QTL定位分析
本研究针对7个与烤烟产量相关的农艺性状进行全基因组QTL扫描,共检测获得30个QTLs,其分别分布在第1(LG01)、3(LG03)、4(LG04)、7(LG07)、13(LG13)、14(LG14)、17(LG17)、22(LG22)和24(LG24)共9个连锁群上(图1;表3)。在检测获得的30个QTLs中,2016年和2017年分别检测获得28个和23个QTLs,21个QTLs在连续2年中均能检测到。检测到与株高相关的QTLs最多,为6个;其次是与叶片数和节距相关的QTLs,各有5个;与茎围和腰叶宽相关的QTLs分别为4个;而与茎叶角度和腰叶长相关的QTLs数最少,为4个(图1;表3)。连锁群LG17上检测到的QTL数目最多,为6个,其次是连锁群LG04(5个QTLs),而连锁群LG13上检测到的QTL最少,仅有1个QTL。其中,LG01、LG03、LG07、LG17和LG22共5个连锁群上的QTLs均是在连续2年中能够检测到。7个性状的加性效应值绝大部分是正值,即增加性状的表型值的等位基因均是来自高值亲本-Y3,此外,也存在少量来自亲本K326的具有负效应值的等位基因(表3)。此结果恰好与两亲本间在7个目标性状上具有较显著的差异(Y3具有比K326高的表型值)及所有性状在RILs群体中超亲分离的结果是一致的。在连续2年检测获得的30个QTLs中,大部分的效应值很小,平均每个QTL仅解释7.22%的表型变异值(R2),其中,2016年和2017年每个QTL分别平均解释表型变异值为7.10%和7.34%。
在株高和节距2个性状上2年均检测到了2个具有较大效应值(R2>20%)的QTLs,即,qPH17(R2=23.18%, 2016;21.37%, 2017)和qIL17(25.75%;20.09%)。此外,叶片数、腰叶长和腰叶宽3个性状在连续2年中也检测到3个效应值大于10%的QTLs。上述5个性状在2年检测出的5个效应值较大的QTLs均位于第17号连锁群(LG17)上,其中,qPH17、qLN17和qIL17定位于LG17近中部的PT53015和PT53756标记之间约1.62 cM(34.522~32.906)区域内;而qLWL17和qWWL17则被定位在LG17的近端处,位于标记TM66721和TM62196之间约3.54 cM(48.881~45.339)范围内,与此同时,具有较小效应值且与茎围相关的QTLs(qSGS17;R2=6.8%,2016和7.17%,2017)也在该位置被检测到(表3;图1)。上述6个QTLs的加性效应值均是正值,即,来自高值亲本Y3的等位基因对6个不同性状在连续2年检测获得的6个QTLs贡献是正方向的。
同样,本研究也发现在烟草基因组中存在一些较小区域(<5.0 cM),每个区域包含2个或2个以上紧密连锁的不同性状QTLs。如qPH3、qIL3和qLN3连续2年检测到3个QTLs均位于LG03的标记TM61400和TM57320之间,qLWL4、qWWL4和qSG4 3个性状2年检测获得的3个QTLs位于LG04上一个跨度约3.85 cM的小区域内,qPH7、qIL7和qLN7 位于LG07上,及qLWL22、qWWL22和qLN22、qIL22位于LG22上(图1;表3)。此结果与本研究中7个目标性状间存在显著的相关性的结果吻合(表2)。
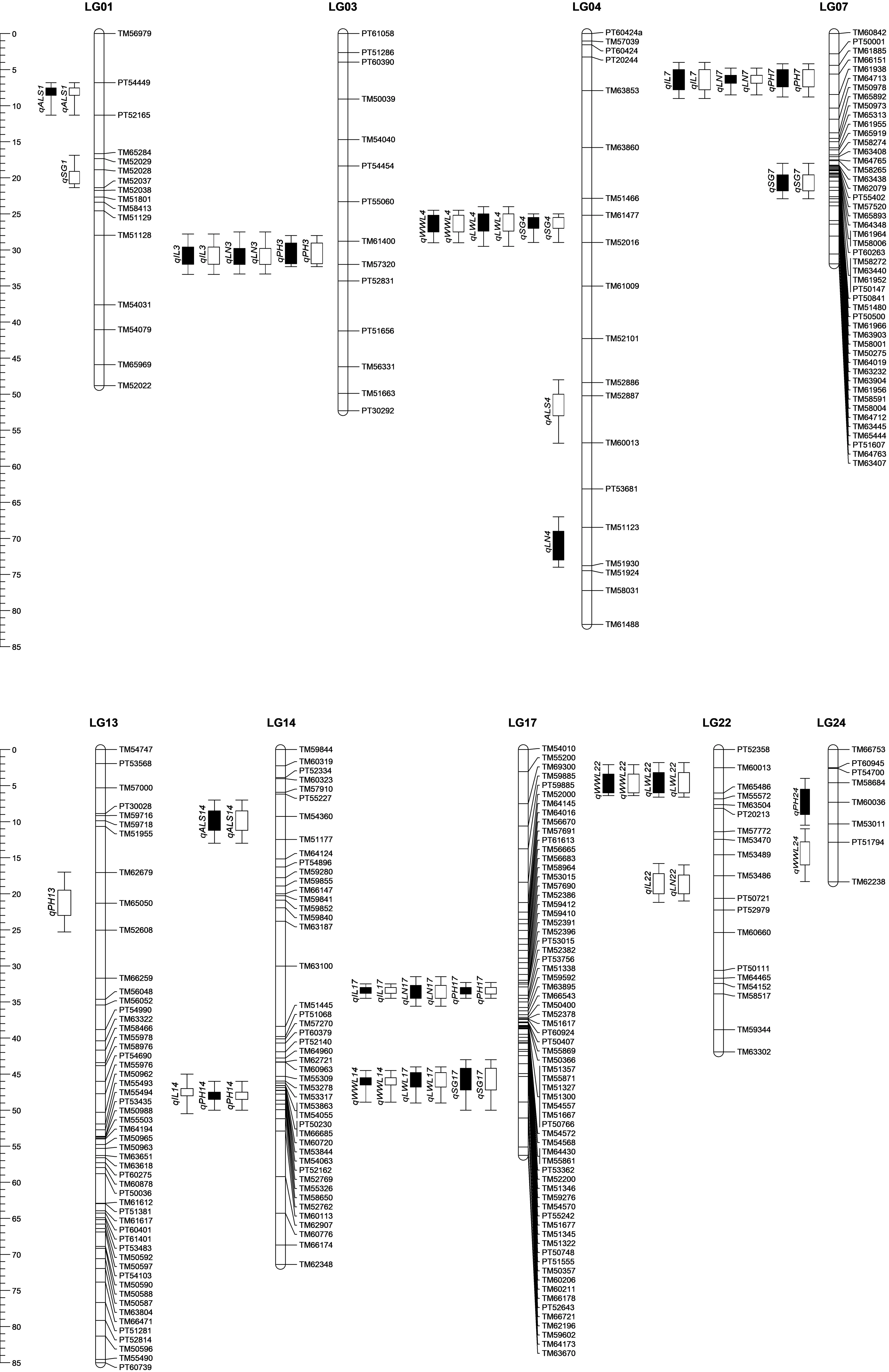
2016和2017年的QTLs分别用黑色空心条和黑色实心条表示图1 利用严格复合区间作图法(rMQM)检测到的7个产量相关性状QTLs在烤烟遗传图谱上的位置信息2016 and 2017 indicated by black hollow squares and black solid squares, respectivelyFig.1 Genetic map of flue-cured tobacco showing the positions of QTLs of seven yield-related traits estimated by composite interval mapping
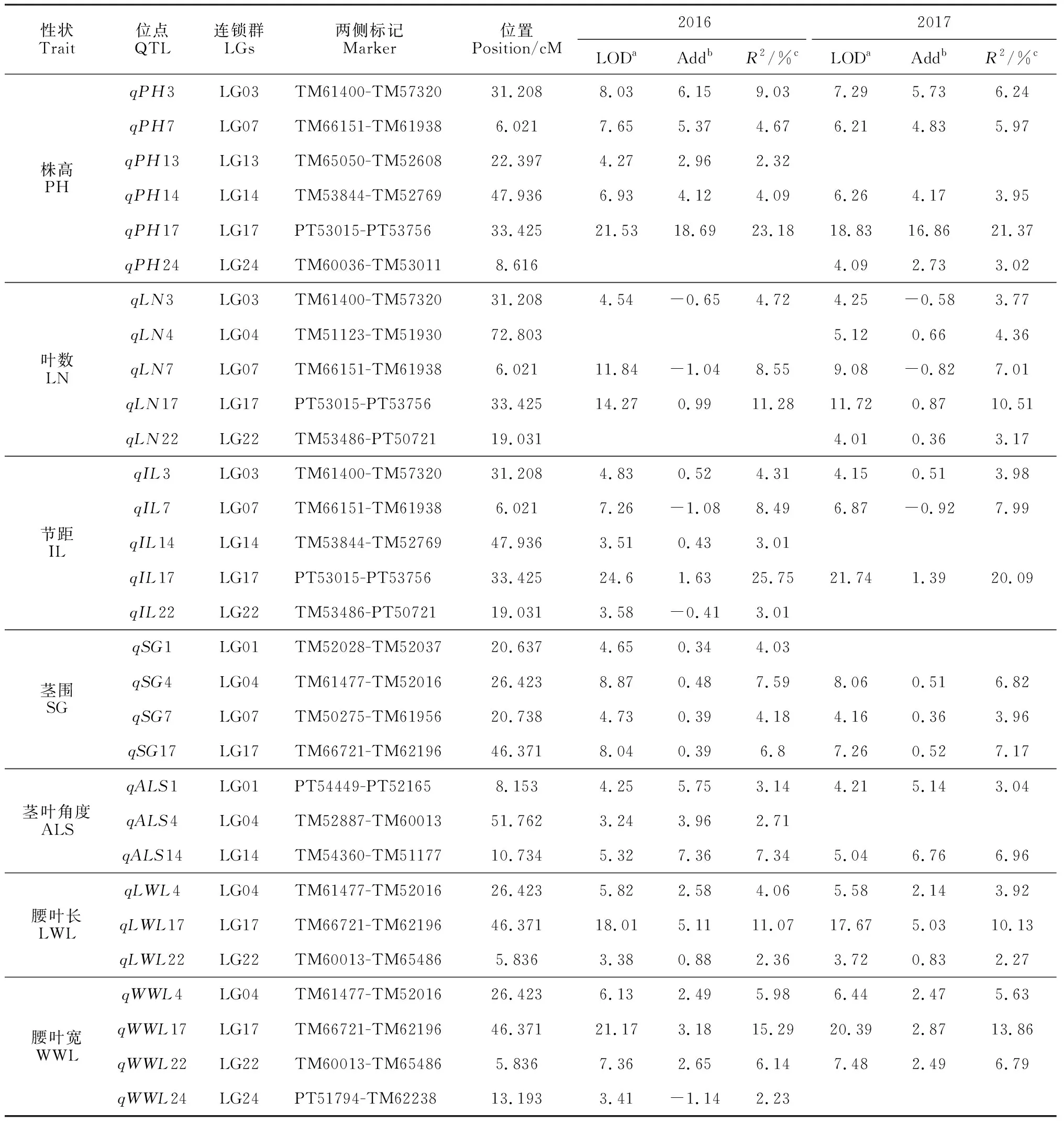
性状Trait位点QTL连锁群LGs两侧标记Marker位置Position/cM20162017LODaAddbR2/%cLODaAddbR2/%c株高PHqPH3LG03TM61400-TM5732031.2088.036.159.037.295.736.24qPH7LG07TM66151-TM619386.0217.655.374.676.214.835.97qPH13LG13TM65050-TM5260822.3974.272.962.32qPH14LG14TM53844-TM5276947.9366.934.124.096.264.173.95qPH17LG17PT53015-PT5375633.42521.5318.6923.1818.8316.8621.37qPH24LG24TM60036-TM530118.6164.092.733.02叶数LNqLN3LG03TM61400-TM5732031.2084.54-0.654.724.25-0.583.77qLN4LG04TM51123-TM5193072.8035.120.664.36qLN7LG07TM66151-TM619386.02111.84-1.048.559.08-0.827.01qLN17LG17PT53015-PT5375633.42514.270.9911.2811.720.8710.51qLN22LG22TM53486-PT5072119.0314.010.363.17节距ILqIL3LG03TM61400-TM5732031.2084.830.524.314.150.513.98qIL7LG07TM66151-TM619386.0217.26-1.088.496.87-0.927.99qIL14LG14TM53844-TM5276947.9363.510.433.01qIL17LG17PT53015-PT5375633.42524.61.6325.7521.741.3920.09qIL22LG22TM53486-PT5072119.0313.58-0.413.01茎围SGqSG1LG01TM52028-TM5203720.6374.650.344.03qSG4LG04TM61477-TM5201626.4238.870.487.598.060.516.82qSG7LG07TM50275-TM6195620.7384.730.394.184.160.363.96qSG17LG17TM66721-TM6219646.3718.040.396.87.260.527.17茎叶角度ALSqALS1LG01PT54449-PT521658.1534.255.753.144.215.143.04qALS4LG04TM52887-TM6001351.7623.243.962.71qALS14LG14TM54360-TM5117710.7345.327.367.345.046.766.96腰叶长LWLqLWL4LG04TM61477-TM5201626.4235.822.584.065.582.143.92qLWL17LG17TM66721-TM6219646.37118.015.1111.0717.675.0310.13qLWL22LG22TM60013-TM654865.8363.380.882.363.720.832.27腰叶宽WWLqWWL4LG04TM61477-TM5201626.4236.132.495.986.442.475.63qWWL17LG17TM66721-TM6219646.37121.173.1815.2920.392.8713.86qWWL22LG22TM60013-TM654865.8367.362.656.147.482.496.79qWWL24LG24PT51794-TM6223813.1933.41-1.142.23
注:a在a=0.05水平下,利用交换测验方法(permutation times = 10 000)在全基因组范围内估计LOD显著阈值:PH和IL的LOD= 3.5,ALS的LOD=3.2,LN的LOD=3.4,SG、WWL和LWL的LOD= 3.3。bAdd指加性效应,其中的正/负号表示来自于亲本Y3/K326的等位基因的作用方向。cR2指QTL解释表型总变异的比例
Note:aLOD thresholds at the genome-wise significance level of 0.05 estimated by 10 000 permutation tests were: 3.5 for PH and IL, 3.2 for ALS, 3.4 for LN, 3.2 for SG, WWL and LWL.b Add is additive effect, of which the positive/negative sign indicates the action direction of the allele from parent Y3/K326.cR2is the proportion of phenotypic variance explained by the QTL
3 讨 论
大量的研究表明,与烟草产量相关的农艺性状属于易受环境影响的数量性状[32,38-39]。蔡长春等[32]利用烤烟DH群体对烟草开花期和叶数2个性状进行了遗传分析,结果表明,2性状存在一定程度的双向超亲现象,且均符合正态分布特征,说明该2个性状属于多基因控制的数量性状。许明辉等[38]利用‘红花大金元’、‘青梗’和‘中烟14号’3份烤烟材料配制的2个组合,经6个世代,对7个农艺性状和4个品质性状进行了遗传分析,结果表明:烟草的株高、叶数、腰叶长、腰叶宽、节距和茎围(茎粗)等6个性状是受微效多基因控制数量性状。李华丽等[39]基于F2:3群体,对6个烟草重要性状进行了遗传分析,结果显示:所观察的6个性状呈连续分布,且各性状均存在不同程度的双向超亲现象,表现出数量性状的特点。本研究通过对烤烟RILs群体连续2年的田间表型数据分析,其结果表明与烟草产量相关的7个农艺性状属于由多基因控制的数量性状,与已报道的研究结果一致[13,29,34]。
相较于前人有关烟草农艺性状QTL定位分析的报道,本研究进行QTL定位分析所用的遗传图谱具有更高的质量和饱和度,因此,更利于在烟草全基因组范围内进行QTL扫描分析。Julio等[29]基于构建的局部烟草遗传图谱(仅含有18个连锁群)对59个烟草性状(农艺性状、化学成分及品质等)进行了QTL分析,最终共检测到了75个QTLs,平均每个性状仅能检测到1.27个QTLs。蔡长春等[32]基于160个标记构建的仅含有23个连锁群的不完整烟草图谱,对2个性状进行QTL分析并获得3个QTLs,每个性状可检测获得的QTL数平均为1.5个。同样,李华丽等[39]基于构建获得的部分烟草连锁图谱(190个非特异性标记构建获得包含26个连锁群的烟草遗传图谱)对6个性状进行QTL分析,最终获得11个QTLs,每个性状平均可检测到QTLs数为1.83个。而本研究基于一张包含626个SSR标记和24条连锁群的高质量烟草图谱,对7个目标性状在全基因组范围上进行QTL扫描分析并最终获得30个QTLs,平均每个性状可检测到多达4.3个QTLs。造成上述QTL检测能力(检测获得QTL数量多少)的因素有多种,如,构成群体的亲本间遗传关系、群体的种类、大小及性状的遗传力等,但用于进行全基因组QTL扫描分析的遗传图谱的质量(包含标记的种类、数量、密度、图谱的基因组覆盖度等)可能是关键因素之一。不难发现,上述用于QTL定位分析的遗传图谱质量不高(如,标记的稳定性较差、标记数量较少、图谱的完整度较低等),因此,就极大地降低了对图谱尚未覆盖的众多与目标性状相关的QTLs检测能力。同样是基于SSR标记构建的完整烤烟遗传图谱对6个烟草农艺性状(株高、节距、茎围、叶数、最大腰叶长和宽)进行QTL定位分析,Tong等[34]对目标性状在207个DH株系一年2重复的表型数据进行QTL分析,结果获得多达69个QTLs,其中,有4个QTLs具有较大效应值,可解释约15%~20%的表型变异。与之相比,本研究在检测QTLs数量上要少些,但连续2年均能检测到具有较大效应值的QTLs为5个,其中,与株高和节距相关的2个QTLs效应值均在20%以上。这一结果刚好与株高和节距2个性状在RILs中具有较高的广义遗传率(株高和节距的广义遗传率分别为83.15%和81.33%;表1)相吻合。此外,本研究通过对17号连锁群上连续2年(2016~2017)获得的5个高效应值QTLs 两侧的标记进行检测,发现与株高、节距和叶数3个性状相关的QTLs两侧标记(PT53015和PT53756)在262个RILs中有10个株系与低值亲本K326的基因型相同,其性状均值分别为170.39 cm(PH变异范围:160.83~191.57)、5.21 cm(IL变异范围:4.26~6.17)和35.50(LN变异范围:32.00~40.00);13个株系与高值亲本Y3具有相同的基因型,其性状均值分别为236.33 cm(PH变异范围:207.27~303.77)、9.63 cm(IL变异范围:8.03~12.31)和45.33(LN变异范围:40.00~53.00)。同样,检测到与腰叶长和腰叶宽2个性状相关的QTLs两侧标记(TM66721和TM62196)有7个株系与低值亲本K326的基因型一致,其性状均值分别为62.82 cm(LWL变异范围:58.76~70.04)和38.05 cm(WWL变异范围:36.16~41.03);11个株系与高值亲本Y3基因型一致,其性状均值分别为81.04 cm(LWL变异范围:71.49~92.03)和40.95 cm(WWL变异范围:38.66~58.59)。上述2组的株高值差异为65.94(约占两亲本间PH值差异的72%),解释了约59.6%(qPH17的加性效应值 / 全部株高QTLs的加性效应值)的加性效应值;同样,节距、叶数、腰叶长和腰叶宽等4个性状值在两组间的差异分别为4.42(占两亲本间IL值差异的约76%)、9.83(占两亲本间LN值差异的约70%)、18.22(占两亲本间LWL值差异的约80%)和2.9(占两亲本间WWL值差异的约56%),解释了约43.8%、31.2%、61.2%和35.0%的加性效应值。更有意义的是,本研究中位于第17号连锁群上且连续2年均能检测到的效应值大于10%的5个QTLs,也能够同时在Tong等[34]研究中的相同连锁群上检测到,即,与株高、节距、叶数、腰叶长和腰叶宽5个性状相关的QTLs均在第17号连锁群上同时被检测到。以上结果表明:一方面,本研究的结果具有较高的真实性和可信度;另一方面,与烟草产量相关农艺性状中的株高、节距、叶数、腰叶长和腰叶宽5个性状具有较高的遗传率和较强的稳定性。
另外,在本研究的烟草基因组中存在一些较小区域,每个区域包含2个或2个以上紧密连锁的不同性状的QTLs。由于位于同一区域内的部分QTLs间如此密切,它们很可能是一个具有‘一因多效’的单独QTL,也即它们可能是影响多个性状的相同基因(QTL)[34]。故此,可将这些包含有紧密相连QTL的小区域视作为具有多效性的单个遗传单位,在开展分子标记辅助选择(MAS)育种提高烟草产量时,予以充分考虑这些具有多效性的区域。
猜你喜欢
轮胎工业(2022年4期)2022-07-20
西南农业学报(2022年5期)2022-06-06
轮胎工业(2021年1期)2021-07-19
中国糖料(2021年3期)2021-07-13
农村百事通(2019年17期)2019-10-08
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
