
酿酒酵母高级醇合成路径及关键基因
2018-09-07 02:58刘灿珍秦伟帅孙玉霞赵新节
中国酿造 2018年8期
刘灿珍,秦伟帅,孙玉霞,赵新节*
(1.齐鲁工业大学 山东省微生物工程重点实验室,山东 济南 250353;2.山东省农业科学院农产品研究所,山东 济南 250100)
酿酒酵母(Saccharomyces cerevisiae)作为二倍体微生物,在酒类发酵方面的应用较为广泛,是酒类发酵的主导菌株。其代谢中产生的挥发性化合物包括高级醇(杏仁和花的气味)、乙酸酯、乙基酯(水果和花香味)、中长链的挥发性酸(干酪和汗的气味)、醛类(奶油、水果和坚果气味)等[1],对酒类风味有重要影响。高级醇是指≥3个碳原子的一元醇类(杂醇油),是酵母在酒精发酵过程的代谢副产物,其含量对酒类的香气和品质有很大影响,是评价酒质量的重要指标之一[3]。适量的高级醇会增加酒体的香气复杂性,是酒体香气的重要贡献者,但过量可能会导致脑性瘫痪,影响人体健康[5],因此在不影响酒风味的前提下,降低高级醇的含量具有重要意义。该文综述了高级醇的主要代谢路径及部分关键基因,以期为构建应用于工业生产的低产高级醇菌株提供理论基础。
1 高级醇代谢的相关途径
高级醇的代谢包括埃里希途径(Ehrlich)和合成代谢途径(Harris)。高级醇形成的前体物质α-酮酸由氨基酸的转氨作用形成,之后经线粒体和胞质支链氨基酸转氨酶(branched-chain amino acid transaminase,BCAATases)催化[4],由α-酮酸经脱羧形成醛,并转化为相应的杂醇,称为埃里希途径[5]。若α-酮酸来源于碳水化合物代谢,则称为高级醇的合成代谢途径[4]。
1.1 埃里希途径
GIUDICI P等[6]用酿酒酵母研究了氨基酸形成高级醇的形成机制,指出形成高级醇的埃里希途径为:氨基酸在转氨、脱羧、还原作用下经酮酸、醛最终形成相应的高级醇(比与原氨基酸少1个碳)。之后对该途径在酶及基因调控方面进行了丰富,有研究发现[7],支链氨基酸通过支链氨基酸通透酶(由BAP2基因编码),转运至细胞;然后由BAT1编码的线粒体转氨酶作用催化氨基酸为α-酮酸,基因BAT2参与胞质支链氨基酸转氨酶的作用,催化α-酮酸合成氨基酸的路径;最后经2-酮基酸脱羧酶(2-keto-acid decarboxylase,KDCs)脱羧,脱氢酶(dehydrogenase,ADHs)还原为高级醇。苯丙氨酸、亮氨酸、异亮氨酸、缬氨酸、蛋氨酸也通过该途径生成相应高级醇。
1.2 合成途径
合成途径(如图1所示)是由葡萄糖代谢的中间产物α-酮酸,经2-酮基酸脱羧酶(KDCs)脱羧、脱氢酶(ADHs)脱氢还原成高级醇[3]。葡萄糖经糖代谢生成丙酮酸,再转化为α-酮丁酸、α-异酮戊酸、α-酮异己酸,最后分别生成活性戊醇、异丁醇、异戊醇。其中活性戊醇途径为:α-酮丁酸分别在乙酰乳酸合酶(ILV1基因编码)、还原酶(ILV5基因编码)、脱水酶(ILV3基因编码)作用下催化为α-酮-3-甲基戊酸,再经脱羧、还原作用形成;异丁醇途径为:由α-酮异戊酸经脱羧作用形成异丁醛,再经还原作用形成异丁醇;异戊醇途径:α-酮基异己酸形成异戊醛,再还原为异戊醇,其中α-酮丁酸在ILVs基因编码的相关酶催化下转化为α-酮基异戊酸,在LEU4、LEU1基因编码的相关酶的催化下,可转化为α-酮异己酸[7]。而形成α-酮丁酸的途径有3条,丙酮酸途径:丙酮酸经LEU1基因编码的异丙基苹果酸合成酶催化为β-苹果酸甲酯,β-苹果酸甲酯再转化为α-酮丁酸,其中α-酮异戊酸到α-酮异己酸(亮氨酸合成途径的前体物质)的转化也需要LEU1基因参与,因此,该基因的缺失可能对高级醇及亮氨酸的合成造成一定影响;天冬氨酸途径:天冬氨酸转化为高丝氨酸,高丝氨酸在THR4基因编码的苏氨酸合成酶催化下合成苏氨酸,苏氨酸经ILV1基因编码的苏氨酸脱氢酶作用,转化为α-酮丁酸[8];高丝氨酸合成途径[7]:可能是高丝氨酸转化为中间物质O-乙酰高丝氨酸、胱硫醚,最后形成最终产物α-酮丁酸,该途径未见详细报道(如图2所示)。
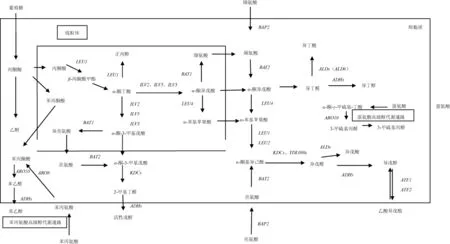
图1 高级醇代谢通路Fig.1 Metabolic pathways of higher alcohol
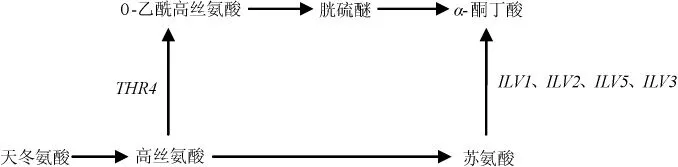
图2 天冬氨酸合成α-酮丁酸途径Fig.2 Synthesis pathway of α-keto butyric acid using aspartic acid
2 高级醇调控关键基因及功能
2.1 埃里希途径中关键基因及功能
2.1.1 BAP2基因
酿酒酵母BAP2基因编码支链氨基酸通透酶,该酶参与3种支链氨基酸包括亮氨酸,异亮氨酸和缬氨酸的转运,底物特异性较广泛[9],上述3种氨基酸可通过埃里希途径形成高级醇[10]。GRAUSLUND M等[9]研究发现,BAP2基因的缺失导致亮氨酸,异亮氨酸和缬氨酸利用量减少20%~50%。也有研究发现BAP2基因的组成型表达导致异戊醇生产增加,因此BAP2基因表达和高级醇的形成之间存在一定的相关性[11]。
2.1.2 BAT1、BAT2基因
Bat1p和Bat2p酶分别位于线粒体和细胞溶质,分别催化支链氨基酸和α-酮酸间氨基的转运,而α-酮酸是合成高级醇的前体,因此会间接影响酵母发酵产生的口感和香气[12]。YOSHIMOTO H 等[13]研究发现,BAT1和BAT2基因分别编码Bat1p和Bat2p两个支链氨基酸转氨酶(BCAATases)。BAT2基因参与支链醇、乙酸异戊酯的生产,BAT2基因缺失突变体KY1058与野生型相比,异戊醇、异丁醇的量分别减少40%、72%[13],另一方面过表达BAT2基因,异戊醇、异丁醇量分别增加几倍[14]。此外,双敲除BAT1和BAT2基因的单倍体酿酒酵母,仍能产生异戊醇[15],表明其他酶能够补偿支链氨基酸转氨酶缺失带来的影响。此外,有研究表明,Bat1p优先参与支链氨基酸的合成,Bat2p参与缬氨酸的分解代谢,表明了支链氨基酸转氨酶的功能多样化[16]。MA L等[17]敲除CAR1、BAT2并过表达BAT1基因构建了一株低产氨基甲酸乙酯同时低产高级醇的酵母菌株。
2.1.3 ARO基因
芳香族氨基酸苯丙氨酸经苯丙氨酸转氨酶(ARO8基因编码)、脱羧酶(ARO10基因编码)催化形成β-苯乙醇,脱羧酶是β-苯乙醇生物合成途径的关键酶,过表达ARO10基因β-苯乙醇的产量提高[18]。ARO80、CAT8、MIG1是β-苯乙醇合成重要转录因子,过表达CAT8基因或敲除MIG1基因均影响基因ARO9与ARO10的转录,使苯乙醇的产量增加[19]。而脱羧酶对转氨产物α-氨酮基-γ-甲硫基-丁酸(蛋氨酸转氨物)的脱羧作用尤为突出[20],脱羧酶也是3-甲硫基丙醇生物合成中的关键酶,ARO10基因过表达使3-甲硫基丙醇的产量提高[21]。
2.1.4 HOM2基因
生物合成苏氨酸、蛋氨酸的天冬氨酸β-半醛脱氢酶由HOM2基因编码,因此HOM2基因表达可能会影响2种氨基酸的产量,从而导致高级醇含量的变化。ROBICHONSZULMAJSTER H等[22]提出,天冬氨酸β-半醛脱氢酶是埃里希途径的关键酶。缺失HOM2基因的菌株,异丁醇和异戊醇的生成量显著降低[23]。齐亚楠[24]研究了HOM2基因对菌株发酵能力与高级醇的影响,发现敲除HOM2的一个等位基因,对酵母菌的发酵性能没有明显影响,但异戊醇的生成量降低。敲除HOM2的2个等位基因则严重影响酵母菌生长,同时高级醇含量明显降低。这可能是由于天冬氨酸β-半醛脱氨酶影响了埃里希代谢途径,也可能是菌体自身生长减弱造成的。
2.2 合成途径关键基因及功能
2.2.1 LEU基因
异丙基苹果酸合成酶由LEU1基因编码,是形成α-酮丁酸和α-酮异戊酸到α-酮异己酸的转化过程的重要编码基因。因此该基因缺失会对丙酮酸到α-酮丁酸的形成途径有阻断作用,对正丙醇的产量产生影响;同时该基因缺失影响α-酮酸(α-酮异戊酸-α-酮异己酸)间的转化,导致异戊醇产量降低;而α-酮异戊酸的积累也会导致异丁醇的生成量增加[25]。有研究通过敲除LEU1基因发现正丁醇、异戊醇的含量提高,同时发现LEU1基因的敲除,使ILV1基因(编码苏氨酸脱氢酶)表达上调[25]。LEU1是合成亮氨酸的关键基因,通过单敲除及双敲除LEU1的等位基因发现异丙基苹果酸脱氢酶的酶活降低,异戊醇、总高级醇的产量均降低[26]。LEU2基因编码β-丙基苹果酸脱氢酶,可催化β-丙基苹果酸合成α-酮异己酸(异戊醇的前体)[6],因此该基因缺失会减少α-酮异己酸的量,从而使异戊醇的产量下降,有研究敲除LEU2基因,异戊醇含量较出发菌株降低了11.82%,高级醇总量降低9.97%[6]。
2.2.2 ILV基因
与支链氨基酸合成相关的酶有4种,由ILV基因编码。其中异亮氨酸的合成由苏氨酸脱氨酶(ILV1基因编码)、乙酰乳酸和酶(ILV2基因编码)、乙酰羟酸还原酶(ILV5基因编码)、脱水酶(ILV3基因编码)、转氨酶(BATs基因编码)共同催化。同时在上述后4种酶的作用下丙酮酸催化为缬氨酸,缬氨酸是合成高级醇的前体物,因此ILV基因的表达会影响高级醇的合成[27]。在酿酒酵母中分别过量表达编码4种酶的基因,异丁醇的产率均升高[28]。BOLLON A P[29]证明,酿酒酵母苏氨酸脱氨酶参与催化异亮氨酸生物合成的第一步反应,而且也参与异亮氨酸-缬氨酸的生物合成的调控途径。苏氨酸脱氨酶催化苏氨酸转化为α-酮丁酸,有研究证明[30]ILV1基因的缺失对乙酰羟酸还原异构酶和支链氨基酸转氨酶的调节没影响,但导致异丁醇生产率提高到3.5倍。同时EDEN A等[31]指出,ILV2基因缺失导致中间物α-酮丁酸积累,使正丙醇的生成量增加。
2.2.3 PDC基因
脱羧酶由PDC1、PDC5、PDC6、YDL080c或YDR380w基因编码,使酵母能利用α-酮-β-甲基戊酸[32]。过表达酿酒酵母的ILV2、KDC、ADH基因,并且敲除PDC1基因(编码丙酮酸脱羧酶),发现异丁醇的产率是出发菌株的13倍[33],原因可能是丙酮酸合成乙醇的途径受到抑制,从而导致过多的碳源流向埃里希途径,进而增加了异丁醇产量。也有研究发现[33],敲除编码丙酮酸脱氢酶复合体(负责将丙酮酸催化为乙酰辅酶A(coenzyme A,CoA))的基因:PDA1、PDB1、LAT1或LPD1基因,导致异丁醇生产更多。敲除编码丙酮酸脱羧酶的同工酶基因PDC1基因,导致异丁醇产量增加[34]。类酮酸脱羧酶(YDL080c基因编码),是α-酮异己酸生成异戊醇的关键酶,研究发现[34],敲除该基因不能合成类丙酮酸脱羧酶,导致异戊醇的产量显著减少,可能是由于糖代谢合成高级醇的途径受阻后,氨基酸代谢为高级醇的分解途径活跃,而高级醇产量主要来自于糖代谢,因此异戊醇的产量下降。但也有研究发现,YDL080c基因缺失并未造成异戊醇的生成量的降低[35]。
3 展望
高级醇的研究开始于1906年,经过110多年的发展,相关研究已经从生化分析阶段进入到分子生物学阶段。目前对高级醇的研究已取得很大进展,并且不断丰富高级醇的代谢路径,找到了高级醇的部分代谢通路及调控基因。该文综述了部分高级醇形成路径埃里希途径、合成途径及主要的调控基因BAP2、BAT1、BAT2、LEU、ILV、THR4、PDC、ARO、HOM2,由于上述基因是埃里希途径及合成途径的关键基因,因此综述上述基因对于理解高级醇的代谢具有重要意义。目前在高级醇的研究方面由集中于高级醇产量的影响因素和通过何种生产工艺控制高级醇的产量方面,进一步深入研究高级醇代谢机理,以期通过分子生物学的方式将酿酒酵母进行菌种改造,从而达到工业生产的要求。然而通过基因工程手段构建高产酯,低产高级醇的酵母菌株也存在相应问题,如构建的酵母菌株中存在抗生素选择标记,因此在应用于工业生产方面还存在安全问题。同时,对于基因调控、各通路间相关关系方面还存在研究空间。因此在研究代谢通路的同时,也应在工业应用及发酵条件对高级醇调控基因的表达方面继续研究。
猜你喜欢
能源化工(2022年2期)2023-01-15
高分子材料科学与工程(2022年10期)2022-12-13
食品与发酵工业(2022年15期)2022-08-18
当代水产(2022年1期)2022-04-26
当代水产(2022年2期)2022-04-26
农业机械学报(2021年10期)2021-11-09
中国饲料(2021年17期)2021-11-02
当代水产(2021年2期)2021-03-29
汽车文摘(2017年7期)2017-12-08
北京航空航天大学学报(2014年1期)2014-12-19
