
醋栗番茄LA2093渐渗系的构建及花序相关性状QTL分析
2019-06-26 12:24周艳朝蒋芳玲孙敏涛文军琴李思琦薛灵姿
江西农业学报 2019年6期
周艳朝,蒋芳玲,孙敏涛,刘 帅,文军琴,李思琦,薛灵姿,吴 震
(南京农业大学 园艺学院/农业农村部 华东地区园艺作物生物学与种质创新重点实验室,江苏 南京 210095)
番茄(SolanumlycopersicumL.或LycopersiconesculentumMill.)是茄科(Solanaceae)番茄属(Lycopersicom)草本植物,是一种重要的世界性蔬菜。因其具有风味独特、营养丰富等特点,在世界各地被广泛栽培和食用,在鲜食和加工蔬菜供应中占有重要地位。栽培番茄在许多果实特性如形状和大小等方面表现出广泛的变异性[1]。但总体来看,栽培番茄遗传背景狭窄,仅显示有限的基因型和变异表型。栽培番茄的祖先樱桃番茄从起源地南美向中美洲和欧洲传播过程中,其遗传多样性大量丧失。长期驯化和现代育种的“瓶颈”效应使得栽培番茄的遗传背景日益狭窄[2],甚至缺乏必要的育种资源。据统计,番茄栽培种仅占番茄基因库总遗传变异性的5%[3]。相反,番茄野生种因其在农业和生物学的重要特性上具有丰富的遗传多样性,成为改善番茄栽培性状的有益变异来源[4]。早在20世纪40年代,与栽培番茄相近的野生种就被用于抗病基因的挖掘,进行番茄抗病育种[5]。番茄野生种是番茄进行遗传改良的优异基因库。醋栗番茄(S.pimpinellifolium),又称细叶番茄,是与栽培番茄最近的一个野生近缘种,与栽培番茄杂交亲和,容易通过有性杂交实现遗传物质转移,且具备栽培番茄所缺乏的优异性状,已被广泛用于果实品质[6-7]以及抗病性(番茄黄花曲叶病[8]、疫霉病[9]等)的改良。已鉴定出在醋栗番茄LA2093的第7和12染色体上两个控制番茄红素的主要QTL位点(lyc7.1和lyc12.1)[10]。醋栗番茄种已经被鉴定为非生物胁迫基因的潜在来源[11-12]。
随着作图群体和分子标记辅助选择(marker-assisted selection, MAS)的快速发展,挖掘野生资源基因库中的有益基因成为现代作物育种的重要途径,这将突破传统育种遗传背景狭窄的瓶颈限制。渐渗系(Introgression lines, ILs)是一组含有相同遗传背景,仅在个别染色体片段上存在差异的株系。构建渐渗系是将重要农艺性状从野生近缘种转移到栽培番茄种的优良途径,其能够解决栽培番茄遗传背景狭窄、必要农艺性状匮乏的问题。理想的渐渗系是单片段渐渗系,除了一个渗入片段来自供体亲本外,基因组的其他部分全部与受体亲本相同,并且通常背景回复率超过95%,从而避免一些可能的不育性以及栽培种和野生种杂交中所遇的不良连锁阻力效应等问题。理想的渐渗系群体,渐渗系和受体亲本间的任何表现型差异均可认为由此渗入片段引起,因此可被看作单因素的孟德尔因子,是进行QTL定位的优良材料[13]。Wehrhahn和Allard(1965)首先构建了小麦的渐渗系,并利用含有较短供体亲本渗入片段的回交自交系有效地进行单个QTL效应的估计[14]。随后的几十年中,在水稻、番茄、油菜、甘蓝、玉米、甜瓜等作物上都相继构建了渐渗系[15]。
迄今为止,番茄渐渗系的构建已有报道,如利用野生番茄S.pennelliiLA716[16]、S.habrochaitesLA1777[17]、S.lycopersicoidesLA2951[18]、S.habrochaitesLYC4[19]、S.pennelliiLA716[15]和S.pimpinellifoliumTO-937[20]等分别构建了比较完整的渐渗系群体。但国内番茄渐渗系群体研究起步较晚,而且多利用潘那利番茄渐渗系进行研究,以醋栗番茄为供体构建渐渗系的研究鲜见报道。遗传图谱是构建渐渗系的基础,但在渐渗系构建过程中,多数研究者仅参考公共数据库中已有的高密度遗传图谱进行分子标记辅助选择,从而筛选单片段渐渗系,虽省时省工,但部分分子标记顺序的差异会影响一些片段数目及大小的准确检测[13]。有研究者证明,先构建遗传连锁图谱再筛选单片段渐渗系,结果更可靠[21-22]。花序性状是番茄产量的重要构成因素,利用高代回交群体进行花序相关性状的QTL研究也较少。鉴于此,本研究以野生醋栗番茄LA2093(S.pimpinellifolium)为父本,以栽培番茄‘Jina’(S.lycopersicum)为母本,利用InDel、SSR 标记在BC1代先构建遗传连锁图谱,在此基础上,通过回交结合分子标记辅助选择,建立了一套以‘Jina’为遗传背景、含LA2093基因的渐渗系群体,并利用BC3F1群体对花序相关性状进行了QTL初步定位。醋栗番茄LA2093渐渗系的构建可为挖掘野生番茄的优异基因提供新的遗传材料,为番茄重要农艺性状的遗传机理剖析提供基础群体,对指导番茄分子育种具有重要意义。
1 材料与方法
1.1 试验材料与群体构建
1.1.1 试验材料 本研究所用材料为野生醋栗番茄LA2093(S.pimpinellifolium)和栽培番茄‘Jina’(S.lycopersicum),均由南京农业大学蔬菜生理生态实验室保存。
供体亲本:醋栗番茄LA2093,从美国加州大学番茄遗传资源中心(Tomato Genetics Resource Center, TGRC)引进。其为无限生长型,株型为蔓性,茎叶茸毛短稀。花柱长于雄蕊,花序类型为单式花序,始花节位为3~5节位,单花序花数可达20~30朵;果实呈圆形,成熟果红色,单果重1.02 g左右,可溶性固形物含量在7.2%左右,早熟,耐裂果、耐热性强。
受体亲本:栽培番茄‘Jina’,从荷兰引进并经6代以上自交筛选的纯合优良品系。其为无限生长型,株型为半蔓性,茎叶茸毛短密。花序类型为多歧花序,始花节位为7~8节位,单花序数可达35~45朵;果实呈长圆形,成熟果红色,单果重15.32 g左右,可溶性固形物含量在9.8%左右。
1.1.2 群体构建过程 在2016年3月,以醋栗番茄LA2093为父本,以栽培番茄为母本配制杂交组合并收取F1代种子;在2017年3月,以‘Jina’为母本,以其F1为父本进行回交,得到BC1代种子,2017年8月播种BC1代。从BC1代起利用InDel、SSR分子标记进行全基因组检测。经过GGT软件分析后,在2018年3月,从BC1代84个株系中挑选出18个株系,将其播种后组成378株的BC2代群体。重复上述方法,在2018年8月,从BC2代378个株系中挑选出20个株系,播种组成420株BC3代群体,筛选留种株系,自交得到BC3S1代种子。
1.2 性状调查与统计
在BC3F1群体中,对415个单株的首花序节位(node below the first inflorescence, Nfi)、花序间隔节位(node of interval inflorescence, Nii)、单花序花数(number of flowers per inflorescence, Nfpi)进行调查与统计,调查方法参照《番茄种质资源描述规范和数据标准》。
1.3 分子标记筛选
引物序列信息选自前人[23-25]开发的200对SSR引物、597对InDel引物。利用父母本及F1筛选均匀分布于12条染色体的多态性标记。
采用改良的CTAB法提取DNA。10 μL PCR扩增反应体系含基因组DNA(10 ng/μL) 2 μL、2×TSINGKE Master Mix 5 μL、正向和反向引物(50 ng/μL)各0.4 μL,以ddH2O补足至10 μL。PCR扩增程序:94 ℃预变性4 min;94 ℃变性15 s,55 ℃退火15 s,72 ℃延伸30 s,共循环34次;72 ℃ 5 min;16 ℃保存。聚丙烯酰胺凝胶电泳后,进行银染检测,显色后在荧光灯下统计引物多态性数据。
1.4 数据统计与分析
比对父母本的带型,若PCR扩增产物带型与‘Jina’一致则记为‘a’;若PCR扩增产物带型与LA2093一致则记为‘b’;将杂合带型记为‘h’,将缺失的记录为‘-’。利用选择的亲本间多态性SSR、InDel分子标记,对BC1群体的141个单株的基因型数据进行收集,应用Joinmap 4.0构建遗传连锁图谱。
根据Young和Tanksley(1989)的方法计算代换片段的长度[26]。不考虑两个相邻标记区间发生的双交换事件,当染色体上相邻标记的基因型相同时,认为这两个标记之间的区段由相同的标记基因型组成;当相邻标记的基因型不同时,就认为这两个标记基因型分别组成这个区间的一半。
基因型作图采用 Berloo(2008)开发的 GGT(2.0)作图软件[27],按照软件的要求将EXCEL原始数据转换成GGT数据格式,然后导入到GGT(2.0)版基因型作图软件中,分析染色体内野生渗入片段大小及其所占基因组的比例,并绘出全部单株基因型图示,从而筛选合适的单株材料。
选择标准:(1)入选的单株含有供体亲本LA2093的渗入片段尽可能少,最好只含有1个片段,但片段长度尽可能较长;(2)所有当选单株含有的野生番茄片段尽可能覆盖整个基因组。入选单株授粉后留种下一代播种;(3)对于出现的双交换片段的单株,给予淘汰。
采用Microsoft Excel 2016软件进行花序相关性状的数据统计分析。用WinQTLcart 2.5软件进行QTL定位,采用复合区间作图法以LOD2.5作为QTL位点阈值,对花序相关性状进行分析研究。
2 结果与分析
2.1 番茄遗传图谱的构建
利用受体亲本(栽培番茄‘Jina’)和供体亲本(醋栗番茄LA2093)进行多态性引物的筛选,从200对SSR引物与597对InDel引物中,筛选出了189对多态性引物,亲本间多态性为23.71%。图1为部分标记在亲本间的检测结果。采用189对亲本多态性引物对BC1F1群体内141个单株基因分型的数据进行统计与分析;采用作图软件Joinmap 4.0构建了1张总长1243.80 cM、含158对多态性InDel与SSR分子标记的遗传图谱(其中InDel标记137对,SSR标记21对,删除了31对在群体扩增条带差异性不明显或图谱构建中未能连锁的标记),该图谱包含12个连锁群,对应番茄的12条染色体,平均遗传图距为7.87 cM(表1)。
2.2 追踪标记在染色体组上的分布情况
从所构建的番茄遗传连锁图谱中,每10~30 cM选取1个标记,共选取96个多态性标记(InDel标记82个,SSR标记14个)用于群体构建过程中追踪染色体的变化,这些标记在染色体上的具体位置见图2。每条染色体上标记数为5~11个,平均8个;两个标记间平均图距在9.22~16.86 cM;标记间平均遗传距离为12.96 cM,共覆盖番茄染色体组1243.80 cM。两个标记间最小间距为2.30 cM,间距大于30 cM的有8处,其中最长间距在3号染色体C03M0078与C03M03542之间,长度为47.80 cM。

第1孔道为MARKER;随后每3个孔道为1对引物。

图2 96个标记在染色体上的具体位置与遗传距离
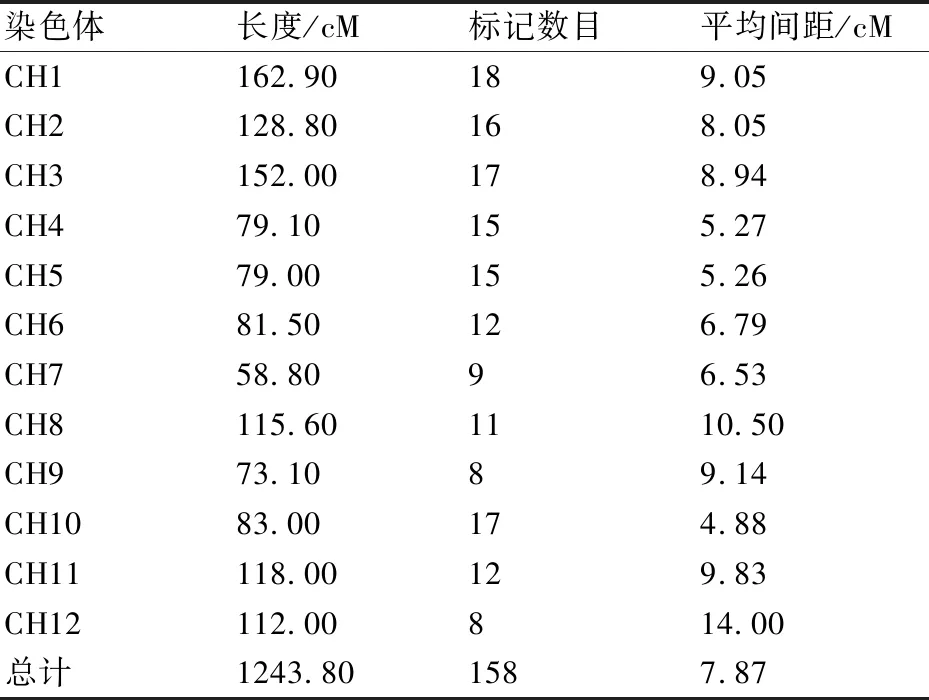
表1 标记在遗传图谱上的分布
2.3 BC3代渗入片段在染色体组的分布情况
依据单株选择标准,经过3代回交并结合分子标记辅助选择,将BC3代415个单株基因型数据导入GGT(2.0)分析后,留选了51份互相重叠、完全覆盖12条染色体的渐渗系材料,其中单片段材料为14份,双片段材料27份,三片段材料10份。渗入片段总长度为3497.80 cM,渗入片段数为98个,平均单个渗入片段长度为35.69 cM,完全覆盖12条染色体。51份渐渗系的渗入片段在染色体组的具体位置详见图3。
从图3和表2可看出,51份渐渗系材料的渗入片段数在12条染色体上分布不均匀,平均每条染色体上渗入8.17个片段。其中1号染色体上的渗入片段数最多,为14个,渗入片段总长度为517.10 cM;5号染色体上的片段最少,为2 个,渗入片段总长度为116.30 cM。98个渗入片段长度在3.35~110.00 cM,其中最长片段位于3号染色体上,长度为110.00 cM;最短片段位于2号染色体末端,长度为3.35 cM。
在51份渐渗系材料中,野生染色体片段平均占受体基因组大小的5.6%,每份材料平均渗入片段总长度为69.65 cM。第48号材料基因组含有的野生片段比例最高,为14.2%;第36号材料基因组含有的野生片段比例最低,仅为0.3%。
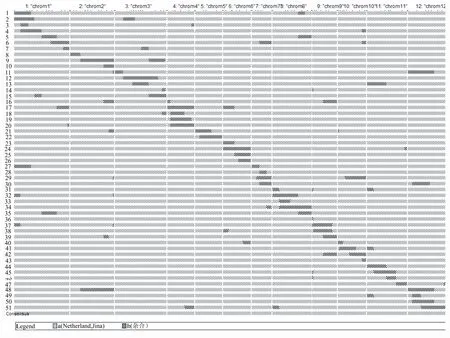
图3 51份渐渗系的渗入片段在染色体组上的具体位置
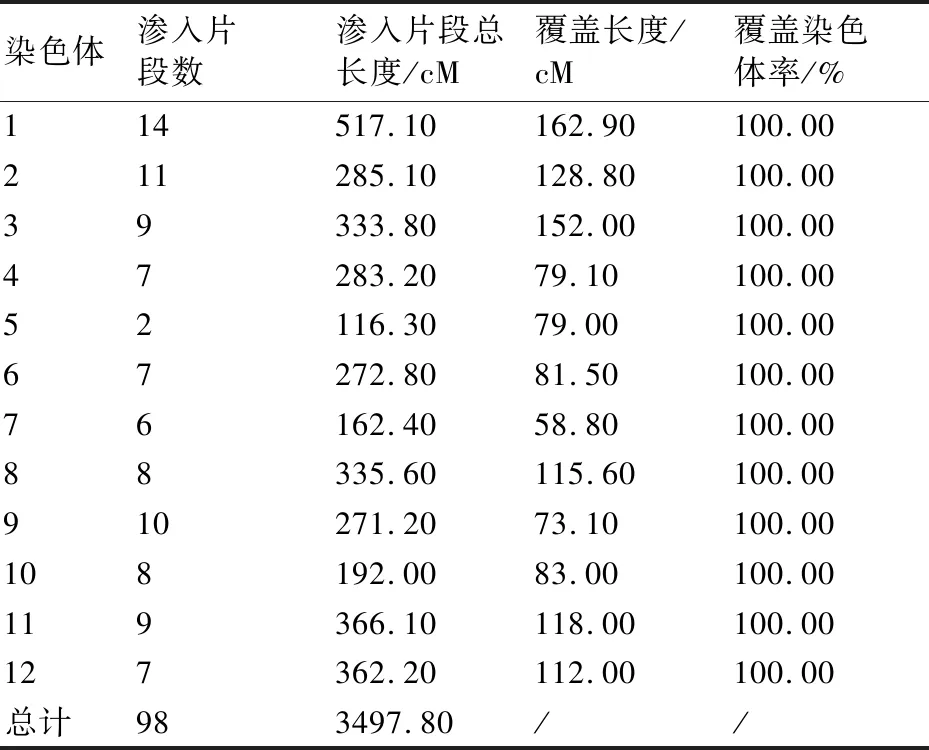
表2 渗入片段在染色体上的分布及覆盖情况
2.4 QTL分析结果
利用复合区间作图法(CIM)对BC3F1群体的首花序节位、花序间隔节位、单花序花数进行QTL定位,共检测到6个QTL,其中控制首花序节位的有2个,控制花序间隔节位的有3个,控制单花序花数的有1个,这些QTL分布于第1、2、4、6、9、12染色体上(表3)。
检测到2个与首花序节位相关的QTL位点:Nfi2.1和Nfi9.1,分别分布于第2、9染色体上,贡献率分别为7.16%和5.51%;与花序间隔节位相关的QTL位点有3个,分布于第1、4、6染色体上,其中,Nii1.1与Nii4.1的贡献率均大于40%,Nii6.1的贡献率为4.33%;与单花序花数相关的QTL位点Nfpi12.1位于第12染色体上,其贡献率为8.09%。其中1个与首花序节位相关的QTL位点Nfi9.1的加性效应为0.64,说明来源于醋栗番茄LA2093首花序节位的等位基因在该位点起增效作用;其他5个QTL位点的加性效应值均为负值,说明源于醋栗番茄LA2093的等位基因起减效作用。
3 讨论
在渐渗系构建过程中,亲本材料的选择决定了渐渗系群体的质量。本研究中选择的供体亲本材料醋栗番茄LA2093,具有优良的抗病性、非生物胁迫抗性以及较高的番茄红素含量等[10,28-30],受体亲本‘Jina’经多年自交选育后综合性状表现优良。构建的渐渗系成功将醋栗番茄LA2093的片段渗入到栽培番茄‘Jina’中,可解决栽培番茄遗传背景狭窄的问题并可深入挖掘更多的优良农艺性状。并且这些渐渗系材料覆盖了整个野生番茄基因组,是在栽培番茄‘Jina’遗传背景上构建的野生番茄基因组文库,可通过聚合育种加快多个优良基因聚合,培育出具有更多优良性状的新品种,缩短育种周期,实现育种目标。

表3 BC3F1群体中花序相关性状QTL定位分析结果
遗传图谱是构建渐渗系的基础。本研究的构建方法是先筛选多态性引物,利用BC1群体构建遗传图谱,并采用分子标记在回交世代同时进行前景和背景选择,使野生番茄片段在BC3世代时占受体基因组的相应比例由理论值12.5%降为5.6%。李学峰利用分子标记在回交世代同时进行前景和背景选择,也将野生黄瓜片段在BC3世代时基因组中相应比例降为7.8%[31]。相比于参考已有高密度遗传图谱再进行渐渗系筛选的方法,该方法保证了标记顺序的准确性。另外在构建渐渗系过程中,若只检测目标片段,则可能会导致部分片段残留[32]。本研究在各回交世代除对候选渐渗系的目标片段进行前景检测外,还对染色体其他部位的遗传背景进行检测,防止了入选单株中存在漏检的残留片段。
渐渗系作为永久性的遗传群体在当前的植物育种中被广泛应用。单片段渐渗系消除了遗传噪音与QTL互作之间的干扰,稳定且不会分离,可进行多年多点的重复实验,保证了QTL的准确性与可靠性;可辨别是否存在有利等位基因,为挖掘作物野生资源的有利等位基因、研究基因功能、进行遗传改良提供了新的工具与材料[33]。张桂权等[34]构建了1529个染色体单片段代换系(CSSL),通过对代换片段的基因鉴定和定位,筛选获得了包括植株形态、稻米品质、产量、抗病性、生育期等重要性状共2000多个优良基因(QTL)。本研究留选了以栽培番茄‘Jina’为遗传背景的51个含有1~3个供体片段的番茄渐渗系。从渗入片段分布情况来看,含有单片段的渐渗系为14份,这些材料可自交以获得纯合株系;而其余材料需要继续回交,以获得更多的单片段材料。此外,某些渗入片段长度较大,究其原因,主要是筛选的多态性标记较少,使遗传图谱的密度不够,也就是在BC1世代构建的图谱不够完善,这也说明虽然本研究的渗入片段在染色体组的覆盖率为100%,但仍有可能有一定的片段检测不到。而Arbelaez J D等在水稻的BC3F1DH代利用GBS技术基因分型提供的标记密度超过原始SSR标记的600倍[22],大大提高了检测的准确性与精确性,省时省工。综上,接下来可通过两种方法进行改善:(1)继续筛选标记,将使标记间平均距离加密到5 cM以内,进一步保证检测的准确性;(2)利用GBS(genotyping-by-sequencing)简化基因组测序技术对群体进行基因分型,以提高标记密度,检测前期因InDel、SSR标记密度稀疏而未检测到的供体渗入片段,并准确定义目标片段的渗入位置与大小。这样可以获得一整套理想的覆盖番茄全基因组的渐渗系,为复杂性状QTL分析及有效转育供体亲本LA2093中优异性状的QTL/基因到其他番茄品种提供理想材料。
花序节位与单花序花数作为番茄的重要农艺性状,是番茄产量的重要构成因子,对指导番茄育种具有重要意义。目前已经发现了许多与番茄花发育相关的基因,包括花分生组织特征基因FA[35]、花序分生组织特征基因SFT[36]以及合轴分生组织特征基因SP[37]等。SFT/SP比率调节开花与分枝,并决定了番茄生殖和营养分生组织中的生长与终止平衡,影响番茄产量[38]。近年来,通过QTL定位挖掘番茄花发育相关基因的研究已有报道。田小琴[39]利用F2群体将控制番茄花序间隔节位的两个QTLs(qN1SU-1-1和qN1SU-1-2)定位在1号染色体上。位江静等[40]利用F2群体检测出控制始花节位的3个QTLs,它们均分布在第1染色体上。李景富等[41]利用F2群体检测出控制番茄始花节位的7个QTLs,分布在第1、2、5和11条染色体上。刘杨等[42]利用146个番茄重组自交系群体在第2、11和12染色体上检测到3个与首花序节位有关的QTLs,在第6、10和12染色体上检测到3个与每花序花数有关的QTLs。本研究利用高代回交BC3F1群体在第1、4和6染色体上检测到与花序间隔节位相关的3个QTLs,在第2、9染色体上检测到与首花序节位相关的2个QTLs,在第12染色体上检测到与单花序花数相关的1个QTL。利用高代回交群体进行农艺性状定位,与利用F2、BC1和RIL群体相比,它使得QTL的检测和有利等位基因的导入相结合,可以消除供体不利基因对回交后代表型的影响[43]。许多研究表明AB-QTL(Advanced backcross QTL analysis)分析法对野生资源中有利基因的发掘是可行的[24,43]。为了保证本研究QTL结果的准确性与稳定性,我们接下来准备继续加密图谱,通过对花序性状数据进行多年多点采集,并通过回交构建这些QTL的近等基因系,验证所定位QTL的真实性,为多个有利等位基因的进一步聚合提供材料。
4 结论
本研究以野生醋栗番茄LA2093(S.pimpinellifolium)和栽培番茄‘Jina’(S.lycopersicum)为材料,在BC1代构建了一张总长1243.80 cM、含158对多态性InDel与SSR分子标记的遗传连锁图谱;以此为基础,在BC3代留选了51份互相重叠、完全覆盖12条染色体的渐渗系材料,其中单片段材料14份,双片段材料27份,三片段材料10份;同时利用BC3F1群体检测到与首花序节位、花序间隔节位、单花序花数3个农艺性状相关的6个QTL位点,单个位点贡献率为4.33%~47.93%。
猜你喜欢
植物研究(2022年4期)2022-08-04
西南农业学报(2022年5期)2022-06-06
中国糖料(2021年3期)2021-07-13
广东农业科学(2021年3期)2021-04-23
塔里木大学学报(2021年1期)2021-03-31
南方农业(2021年1期)2021-02-22
贵州农业科学(2020年8期)2020-09-11
蔬菜(2020年1期)2020-01-17
热带农业科技(2019年1期)2019-01-14
果农之友(2018年5期)2018-10-19
