
大豆G位点近等基因系叶片类囊体蛋白质组比较分析
2020-12-31 05:58赵宇杨邱丽娟
作物杂志 2020年6期
赵宇杨 宋 健 邱丽娟
(1中国农业科学院作物科学研究所/国家农作物基因资源与遗传改良重大科学工程/农业农村部作物种质资源与生物技术重点开放实验室,100081,北京;2长江大学生命科学学院,434025,湖北荆州)
蛋白质组学的概念最早由Wilkins和Williams于1994年提出,蛋白质组是基因组表达的全部蛋白质的总和[1]。根据研究内容,蛋白质组学可分为两类,一类是“差异”蛋白质组学,用于鉴定和筛选不同样本不同条件下的蛋白质组的区别,旨在寻找引起差异的关键蛋白;另一类是“完全”蛋白质组学,是对某一物种中一种组织或器官内全部蛋白质的研究[2]。凝胶电泳和质谱鉴定是蛋白质组研究的两大技术手段。凝胶电泳是分离蛋白质的重要技术,包括SDS-聚丙烯酰胺凝胶电泳和双向凝胶电泳,电泳后将所需要的蛋白条带或蛋白点切割下来进行胶内酶解,然后进行质谱鉴定[3]。质谱技术的不断更新和植物基因组测序的逐步完善推动了植物蛋白质组学研究的发展。
关于大豆蛋白质组的首次报道是利用2-DE法分离大豆种子油体相关蛋白[4],随后从2002年开始逐渐兴起[5],广泛应用于大豆根[6]、茎[7]、叶片[8]、种子[9]等组织器官。其中大豆叶片蛋白质组参考图谱是Xu等[8]首次构建的,他们利用2-DE、MALDI-TOF MS和LC-MS/MS技术对栽培大豆Clark叶片进行蛋白质组分析,鉴定出260个蛋白点。Ahsan等[10]利用2-DE和MALDI-TOF MS技术对栽培大豆Enrei叶片不同发育时期真叶、第1个三出复叶和成熟叶片进行蛋白质组研究,鉴定出超过500种蛋白。郑维薇[11]利用2-DE、MALDITOF MS技术对大豆地方品种、野生品种和育成品种的幼嫩叶片进行分析和比较,鉴定出727种蛋白。大豆叶片蛋白质组在盐[12]、干旱[13]、臭氧[14]和激素[15]等非生物胁迫和不同处理中的相关研究较多,Tian等[16]对耐寒和低温敏感春大豆品种的幼苗叶片在5℃条件下处理12h和24h的蛋白质组进行比较分析,共鉴定到1 320种蛋白,其中有57种差异蛋白,参与13个代谢途径和细胞过程。Gupta等[15]利用HPLC和LC-MS/MS技术对脱落酸(ABA)和乙烯(ET)处理后的大豆叶片进行蛋白质组分析,鉴定到4 192种蛋白。随着研究水平的不断提高,大豆蛋白质组学已从组织转向亚细胞结构。Arai等[17]利用2-DE和MALDI-TOF MS技术对黄化大豆子叶过氧化物酶体进行分析,共鉴定到173种蛋白。Kamal等[18]利用无凝胶和MS/MS技术分析大豆根部线粒体蛋白质组,鉴定到98种蛋白。然而,有关大豆叶片类囊体蛋白的研究尚鲜见报道。
光合作用的光反应发生在叶绿体的类囊体膜上,类囊体膜上主要包括光合系统Ⅰ(PSⅠ)、光合系统Ⅱ(PSⅡ)、细胞色素b6f和ATP复合酶4种蛋白复合物。这些膜蛋白的组成和稳定性对类囊体膜的结构和功能起着至关重要的作用,如基粒的数目、基粒的堆叠程度和光合作用效率等[19-20]。研究这些蛋白质的结构和功能,对于了解类囊体膜光合作用效率和稳定性的分子机制及提高农作物的光合产量来达到高产具有重要意义。
本文以近等基因系NIL-G(绿色种皮)和NIL-Y(黄色种皮)为材料,采用比较蛋白质组学的方法,系统研究叶片类囊体蛋白复合物,从蛋白质组水平了解1对近等基因系叶片的类囊体蛋白之间的差异,为大豆叶片类囊体蛋白的研究提供思路。
1 材料与方法
1.1 材料
前期利用黄色种皮大豆品系ZP95-5383为母本、深褐色种皮野生大豆NY279为父本,于2008年在北京顺义配制杂交组合,2009年在海南种植F1,2010年在北京顺义种植F2,利用F2群体进行定位分析。2010年在海南加代繁殖F3,2011-2013年分别在海南和北京顺义以单粒传方式衍生出重组自交系(RIL)群体。利用开发的dCAPS标记850-dCAPs-MseI检测GmSG基因型,同时利用42对SSR标记鉴定遗传背景,从残余杂合系中选择出NIL-G和NIL-Y,二者遗传背景相似性为97.6%,NIL-G携带GmSG基因型,表现为绿色种皮,NIL-Y携带Gmsg基因型,表现为黄色种皮[21]。Wang等[22]利用GWAS关联到1个与种皮颜色相关的SNP,与G位点是同一个基因,并证明其与种子休眠有关。这对G位点近等基因系用于本试验。将其种植于培养箱中,光周期16h(光)/8h(暗),温度为28℃(日)/26℃(夜),生长至第3个三出复叶完全展开后用于类囊体蛋白试验。
1.2 方法
1.2.1 大豆叶片类囊体膜蛋白检测 大豆叶片类囊体膜蛋白样品的制备:参照Zhou等[23]的方法提取叶片类囊体膜蛋白,稍有调整。分别取1g叶片,加入约5mL预冷的提取缓冲液(0.4mol/L蔗糖,0.01mol/L NaCl,0.002mol/L MgCl2·6H2O,0.05mol/L Hepes,pH=7.8),在冰浴条件下研磨至匀浆,用4层擦镜纸过滤,5 000g 4℃离心10min,类囊体膜用洗涤缓冲液(0.33mol/L Sorbitol,0.05mol/L Bis-Tris,pH=7.0)洗涤数次至无杂质,然后用适量的悬浮缓冲液(0.025mol/L Bis-Tris,20%甘油,pH=7.0)悬浮沉淀,样品于–80℃保存备用。
SDS-PAGE电泳:采用Bio-Rad小型垂直凝胶电泳,浓缩胶浓度5%,电压80V,分离胶浓度12%,电压120V,凝胶厚度为1.5mm。当指示剂溴酚蓝跑到凝胶底部时停止电泳。将完整的凝胶从玻璃板上剥离,放入染色液考马斯亮蓝R-250(冰醋酸60mL,乙醇440mL及考马斯亮蓝R-250 2.25g用超纯水定容至1L)染色1h,取出放入脱色液(冰醋酸50mL和甲醇200mL,用超纯水定容至1L)中直至凝胶背景变透明。
1.2.2 蛋白酶切 首先用灭菌的手术刀将胶切成1mm的小块,置于1.5mL Eppendorf离心管中;用200μL的双蒸水洗涤2次,每次10min;加入考染脱色液[50mmol/L NH4HCO3与ACN(1∶1)]脱色15min,再用双蒸水洗,重复3次,直至胶块完全脱色;加入100μL ACN进行脱水直至胶粒变白,真空抽干 10min;加入 200μL 10mmol/L DTT(25mmol/L NH4HCO3溶解),37℃水浴 1h,加入 100μL ACN进行脱水直至胶粒变白;加入200μL 55mmol/L 生长素(IAA)(25mmol/L NH4HCO3溶解),黑暗处理30min,加入100μL ACN脱水直至胶粒变白;胶粒分别用ddH2O、ACN、ddH2O、ACN进行混悬清洗 ,每 次 10min;将 100μL 的 0.01μg/μL 胰 蛋 白酶工作液(酶液用25mmol/L NH4HCO3稀释)加入胶块中,让胶粒与酶液充分接触,于4℃放置30min至胶粒把酶液完全吸收后,再加入100μL的25mmol/L NH4HCO3,37℃过夜;第2天离心收集酶解上清液,置于一新离心管中;剩余胶粒用抽提缓冲液(5% TFA、95% ddH2O)1h,收集酶解上清液,置上一离心管中合并;剩余胶粒再用缓冲液(2.5%TFA、50% ACN、47.5% ddH2O)抽提1h,收集酶解上清液,置于上一离心管中合并;真空冻干,–20℃保存。
1.2.3 蛋白质谱与数据检索 质谱鉴定由北京青莲百奥生物科技有限公司完成。利用Thermo Q-Exactive型质谱仪完成Label-free的质谱分析,采用MaxQuant软件处理质谱原始文件。参数设置:固定修饰为半胱氨酸碘乙酰胺化(Carbamidomethyl),可变修饰为甲硫氨酸(M)氧化的自发脱氨基,母离子搜索误差±15mg/L,碎片离子质量偏差±0.5Da,最多允许2个胰蛋白酶漏切位点。
1.2.4 数据分析 按照细胞组分、分子功能和生物学过程,利用在线分析工具DAVID比较分析已鉴定的蛋白质质谱数据。利用在线网站KEGG分析蛋白通路。
2 结果与分析
2.1 大豆叶片类囊体蛋白电泳分析
提取叶片类囊体蛋白,经SDS-PAGE梯度凝胶电泳检测,结果(图1)显示近等基因系NIL-G和NIL-Y之间的平行度较好,主条带较清晰,表明蛋白提取效果理想,样品纯度可用于下一步质谱鉴定。为了尽可能鉴定到更多的蛋白,对凝胶条带按照分子量大小分3段(小于25、25~45和大于45kDa)进行质谱鉴定。
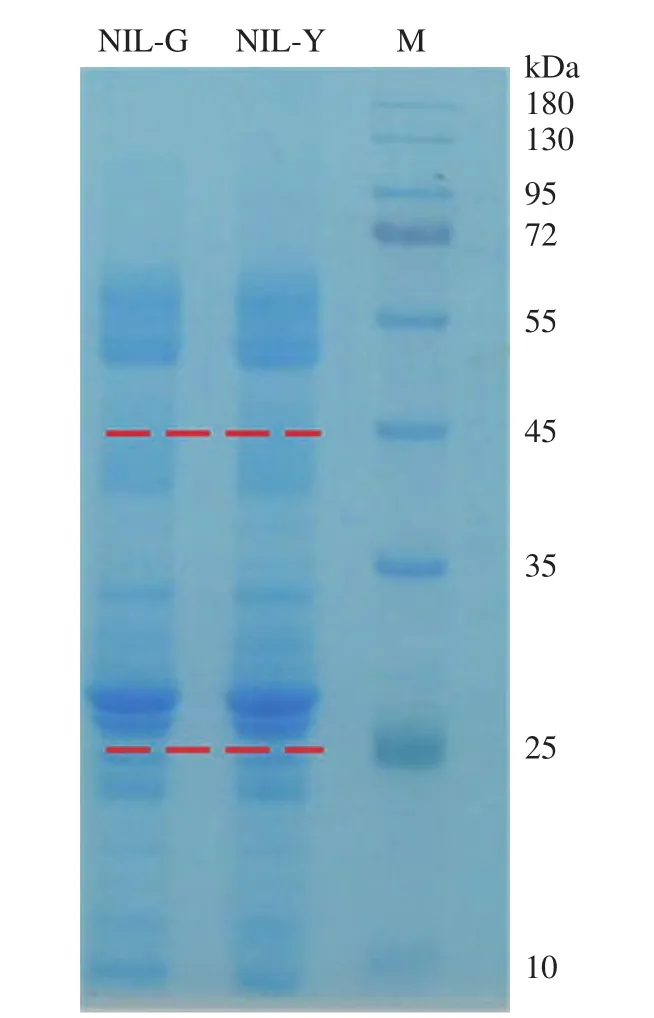
图1 近等基因系叶片类囊体蛋白SDS-PAGE检测Fig.1 Detection of thylakoid proteins in leaves of near-isogenic lines by SDS-PAGE
2.2 质谱鉴定结果
为了提高分析结果的准确性,降低假阳性率,选取报告置信度在95%以上的蛋白并且排除分子量不在选定区域的蛋白,如表1所示,经鉴定,NIL-G中共检测到2 170种蛋白,其中分子量小于25kDa的有485种,25~45kDa的有656种,大于45kDa的有1 029种;NIL-Y中共检测到1 730种蛋白,其中分子量小于25kDa的有373种,25~45kDa的有458种,大于45kDa的有899种。NIL-G叶片类囊体中特异性表达的蛋白1 140种,NIL-Y叶片类囊体中特异性表达的蛋白700种。共检测到相同蛋白1 030种,其中表达存在差异且差异大于3倍的蛋白有68种,NIL-G叶片类囊体中含量高的有23种,NIL-Y叶片类囊体中含量高的有45种。

表1 分段质谱鉴定近等基因系NIL-G和NIL-Y大豆叶片类囊体蛋白数目Table 1 Identification of thylakoid proteins number in soybean leaves of near-isogenic lines NIL-G and NIL-Y by segmented mass spectrometry
2.3 大豆近等基因系叶片类囊体特异蛋白GO注释分析
利用DAVID软件分别对近等基因系大豆叶片类囊体特异蛋白进行GO注释分析,主要分为生物学过程(biological process,BP)、细胞组分(cellular component,CC)和分子功能(molecular function,MF)3类。NIL-G叶片类囊体蛋白参与生物学过程的占55.2%,构成细胞组分的占60.5%,参与分子功能的占68.8%;而NIL-Y叶片类囊体蛋白参与生物学过程的占55.6%,构成细胞组分的占61.2%,参与分子功能的占63.5%。
如图2所示,NIL-G和NIL-Y叶片类囊体特异蛋白均参与大量的翻译、蛋白折叠、信号传导等途径,同时它们都在光合作用过程中发挥功能,但NIL-G叶片类囊体蛋白中特异的是与PSⅠ捕光色素、电子传递及叶绿素生物合成相关的蛋白,NIL-Y叶片类囊体中特异的是与PSⅡ稳定性相关的蛋白。在两者共有的生物学途径中NIL-G鉴定到的蛋白数目均高于NIL-Y。
膜组分、大分子复合物及各系统亚基所占比例最大,说明2种材料中膜组分仍是不可或缺的重要组成成分,它们同时参与PSⅠ和PSⅡ的组成,NIL-G叶片类囊体蛋白中特异蛋白的特异细胞组分为PSⅠ反应中心,而NIL-Y叶片类囊体蛋白中叶绿体基质和内质网膜是其特异组分(图3)。
NIL-G和NIL-Y叶片类囊体特异蛋白中参与核糖体结构组成功能占比最大,核糖体负责转录翻译,这一结果与生物学过程中参与翻译途径为主相吻合。同时特异蛋白的分子功能还集中体现在结合GTP、NAD蛋白及叶绿素。其中NIL-G叶片类囊体特异蛋白中特有电子载体活性、结合铁离子等分子功能,NIL-Y叶片类囊体蛋白中特有结合色素和FMN的分子功能(图4)。
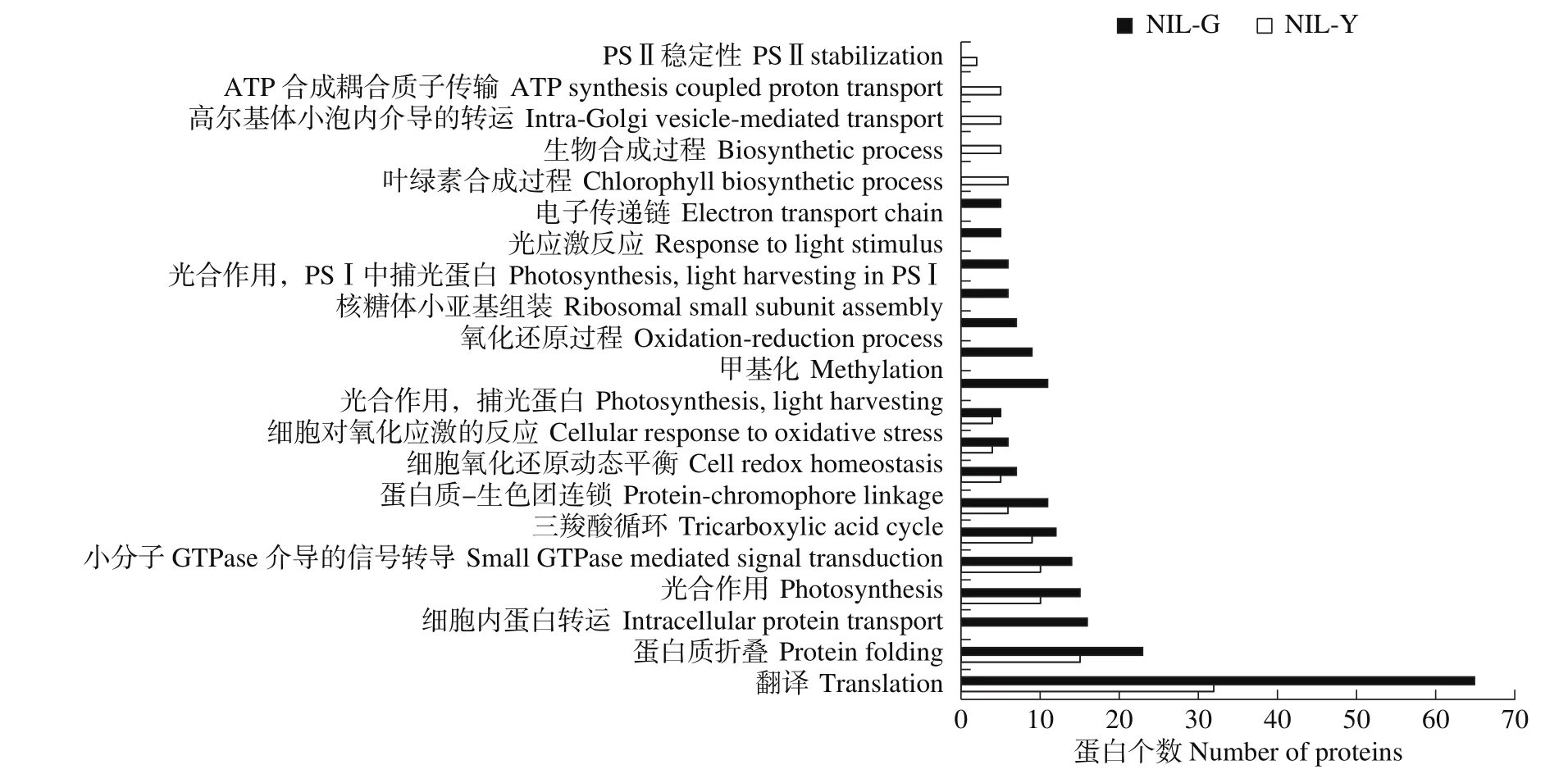
图2 近等基因系NIL-G和NIL-Y特异蛋白的生物学途径水平GO注释Fig.2 GO annotation of specific proteins of near-isogenic lines NIL-G and NIL-Y at biological pathway level
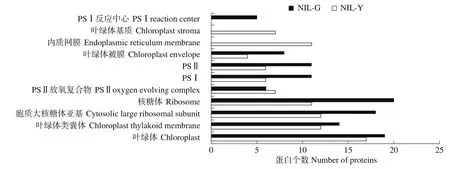
图3 近等基因系NIL-G和NIL-Y特异蛋白的细胞组分水平GO注释Fig.3 GO annotation of specific proteins of near-isogenic lines NIL-G and NIL-Y at cellular component level
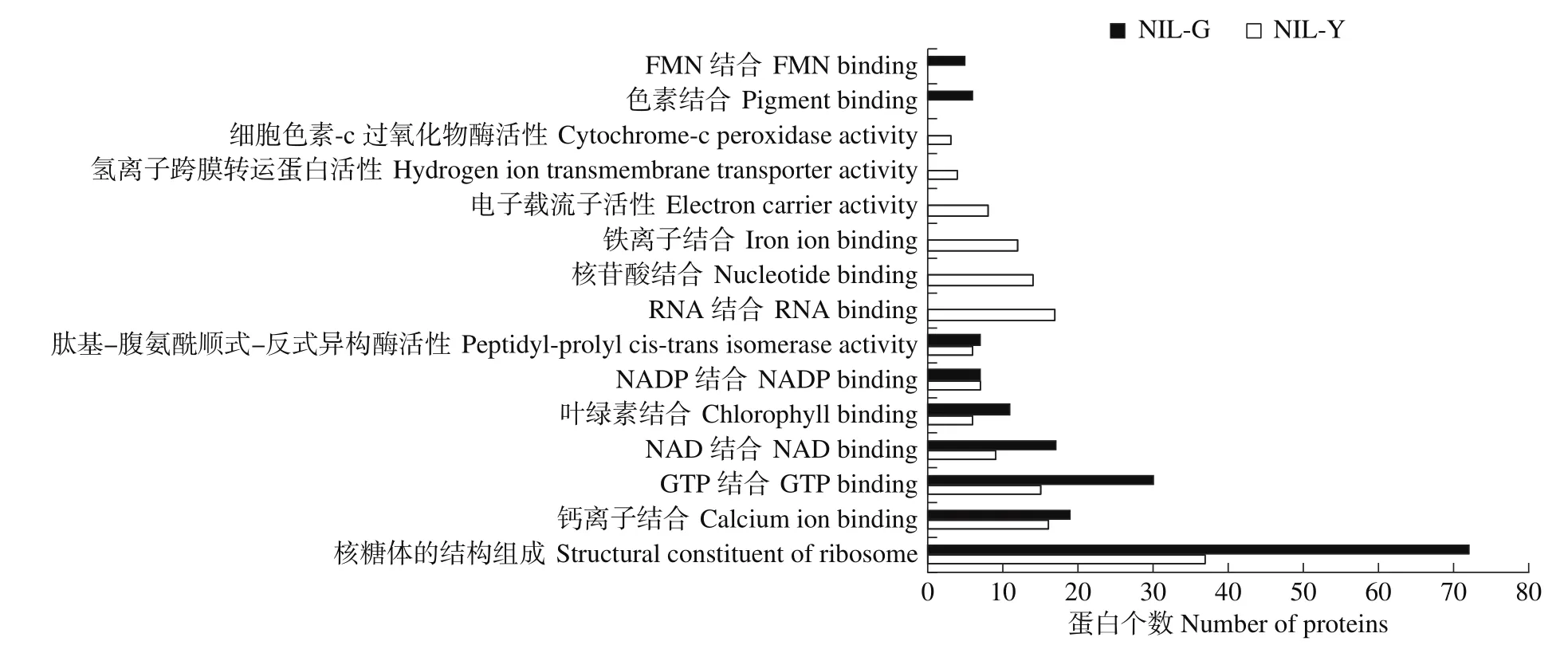
图4 近等基因系NIL-G和NIL-Y特异蛋白的分子功能水平GO注释Fig.4 GO annotation of specific proteins of near-isogenic lines NIL-G and NIL-Y at molecular functional level
2.4 大豆近等基因系叶片类囊体特异蛋白KEGG通路分析
NIL-G叶片类囊体总蛋白中有583种蛋白在KEGG数据库中有功能注释,特异蛋白中有407种蛋白有功能注释,共参与19条代谢通路。参与代谢途径的蛋白最多,共132种,其余蛋白主要参与次生代谢产物生物合成、碳代谢和氨基酸生物合成等,涉及光合作用和卟啉与叶绿素代谢的蛋白有29种(图5),可以看出,处于同一时期的大豆叶片,NIL-G叶片类囊体中参与光合作用和叶绿素代谢途径的蛋白更多。
为了进一步确定总蛋白和特异蛋白参与的通路,在GO注释的基础上,利用KEGG对总蛋白和特异蛋白参与的通路进行分析。结果表明,NIL-Y叶片类囊体总蛋白中有496种蛋白在KEGG数据库中有功能注释,特异蛋白中有276种蛋白有功能注释。从图6可以看出,276种特异蛋白共参与18条代谢通路,参与代谢途径的蛋白最多,有85种,其余分别参与次生代谢产物生物合成、碳代谢和氨基酸生物合成等,参与光合作用和卟啉与叶绿素代谢途径的蛋白较少,共12种。

图6 近等基因系NIL-Y中特异蛋白的KEGG通路分析Fig.6 Analysis of KEGG pathway of specific proteins in near-isogenic line NIL-Y
从表2可以看出,NIL-G叶片类囊体特异蛋白中涉及光合作用和天线蛋白的共有20种,其中未定义蛋白5种,其余15种蛋白分别参与PSⅠ的PsaD,反应中心亚基Ⅱ和Ⅲ;PSⅡ的PsbP和PsbO;光合电子传递链的FNR和Fd;ATPase的gamma链;叶绿素a/b结合蛋白CP24、CP29和P4,LHCII型叶绿素a/b结合蛋白,PSⅡ型叶绿素a/b结合蛋白。NIL-Y叶片类囊体特异蛋白参与光合作用和天线蛋白的共7种,其中未定义蛋白2种,其余5种蛋白分别为PSⅡ的PsbR和Psb27、ATPase的b链、叶绿素a/b结合蛋白6A和CP26。NIL-Y叶片类囊体中的特异蛋白主要与PSⅡ和ATPase相关;而NIL-G叶片类囊体中的特异蛋白不仅参与PSⅡ和ATPase,还与PSⅠ和光合电子传递链密切相关。
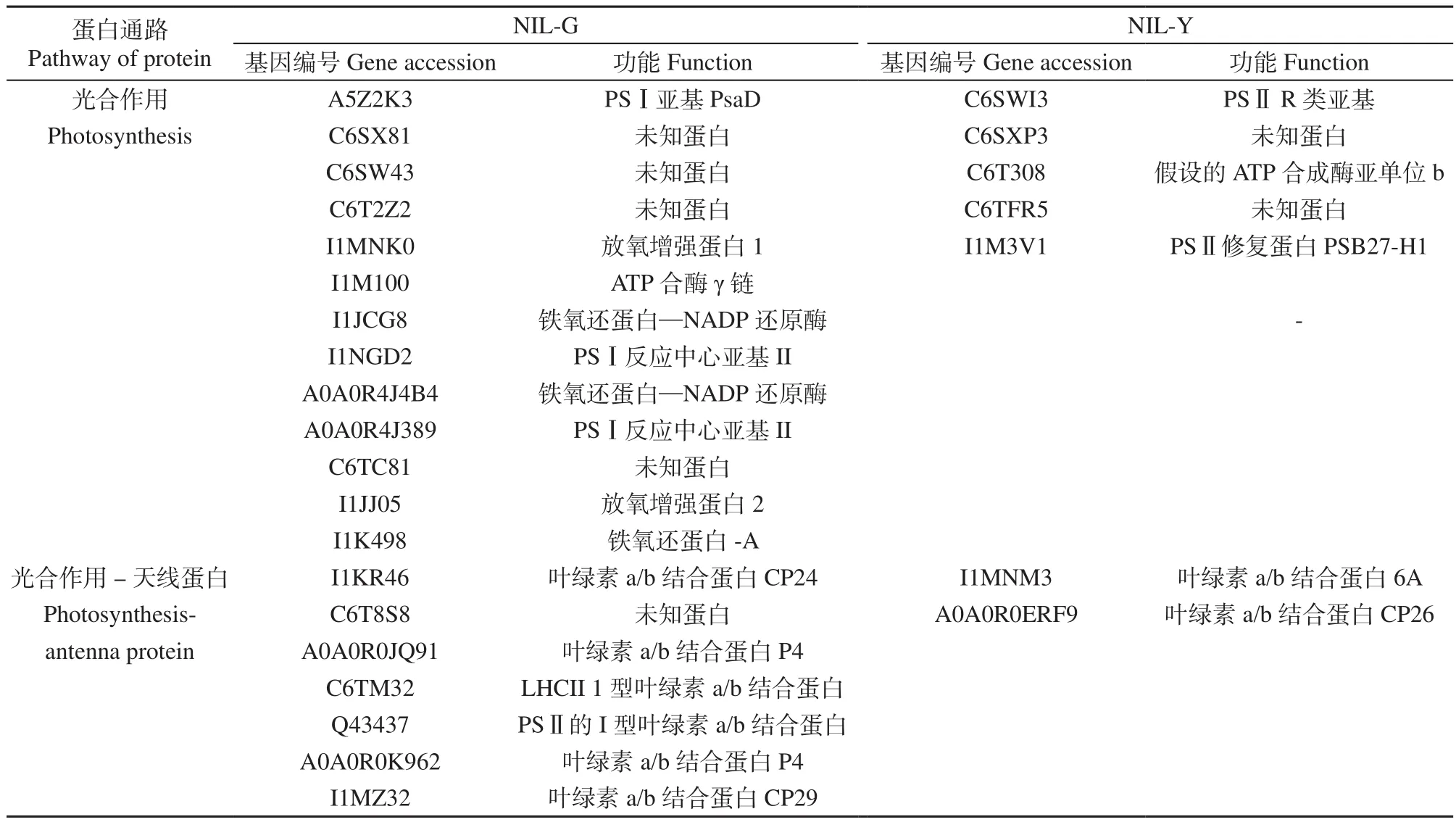
表2 近等基因系NIL-G和NIL-Y特异蛋白中与光合作用和天线蛋白相关蛋白Table 2 Specific proteins related to photosynthesis and antenna in near-isogenic lines NIL-G and NIL-Y
2.5 大豆近等基因系叶片类囊体表达存在差异的蛋白质分析
在NIL-G和NIL-Y叶片类囊体中共有蛋白为1 030种,其中表达存在差异的蛋白(差异>3倍)共68种,包括NIL-Y叶片类囊体中上调的蛋白质45种,其中有功能注释的22种;在NIL-G叶片类囊体中上调的蛋白质23种,有功能注释的17种。将有功能注释的39种蛋白进行层次聚类分析(图7)。这些表达存在差异的蛋白分为两类,第一类以碳代谢及核糖体途径为主,共26种,其中NIL-Y叶片类囊体蛋白占65.4%;第二类主要为光合作用途径,共计13种,NIL-G叶片类囊体蛋白占61.5%。光合作用途径的差异蛋白主要为叶绿素a/b结合蛋白,它们作为光受体在捕获和传递光能过程中发挥重要作用。
3 讨论
3.1 不同物种近等基因系在蛋白质组学研究中的应用
近等基因系是1988年由Young等[24]和Muehlbauer等[25]提出,可通过多代回交、重组自交和分离突变体等途径获得。由于近等基因系仅在目标性状基因存在差异且遗传背景相同,因此是分子水平上遗传研究的理想材料。近等基因系材料广泛应用于基因定位,近年来在小麦[26]、水稻[27]和玉米[28]等作物的籽粒[26]、根[27]和叶[28]等差异蛋白组学研究中的应用也不断加强,筛选出了大量的差异蛋白。例如Lesage等[26]对面包小麦胚乳硬度近等基因系籽粒发育4个阶段蛋白质组分析,鉴定出87种差异表达蛋白,其中54种蛋白是功能已知的蛋白。Torabi等[27]以水稻亲本Nipponbare与其携带主要磷摄取QTL(Pup1)的近等基因系为材料,比较了1μmol/L和100μmol/L磷胁迫下根系蛋白质组的变化,在2-DE凝胶上重复检测出669种蛋白,有32种蛋白在2种基因型上有显著变化,其中低磷条件下存在17种差异蛋白。Wang等[28]利用双向电泳和质谱技术在玉米自交系沈137及其近等基因系第5、10、19片新展开叶主脉中鉴定到24种差异蛋白,其中第10叶期沈137及其近等基因系中存在19个差异蛋白。与其他作物相比,大豆近等基因系的差异蛋白组研究报道极少,仅在大豆闭锁花[29]和蚜虫[30]方面有所应用。Khan等[29]以大豆闭锁花近等基因系为材料,对花芽进行蛋白质组分析,鉴定出640种蛋白质,其中表达差异蛋白19种。本研究首次利用比较蛋白质组分析G位点近等基因系对幼嫩叶片进行鉴定,不仅分别鉴定出NIL-G和NIL-Y特有蛋白1 140种和700种,还鉴定出表达量存在差异的蛋白68种,为阐明大豆叶片类囊体蛋白质组解析提供了重要参考。

图7 近等基因系NIL-G和NIL-Y表达存在差异的蛋白质层次聚类分析Fig.7 Hierarchical clustering analysis of proteins with different expression of near-isogenic lines NIL-G and NIL-Y
3.2 植物类囊体蛋白的研究进展
类囊体是光合作用和电子传递离不开的载体,是叶绿体的核心组分。高等植物叶绿体中的类囊体膜主要包含4种高丰度的多亚基膜蛋白,分别为PSⅠ、PSⅡ、细胞色素b6f复合物(Cytb6f)和ATP合酶(ATPase)。电泳技术和质谱技术的结合促进了植物类囊体蛋白质组学的发展[31-34]。Kashino等[31]对蓝藻类囊体PSⅡ中鉴定到的31个蛋白进行分析,发现了5种新的功能未知蛋白。Steven等[32]利用LC-MS/MS技术鉴定出525种拟南芥类囊体蛋白,这些蛋白在光合作用能量传递和转化过程中发挥重要作用。Peltier等[33]对豌豆类囊体蛋白进行分析,鉴定到200~230种可溶和外周蛋白。Shao等[34]利用BN/SDS-PAGE双向电泳结合质谱的方法对水稻叶片类囊体进行分析,共鉴定出58个蛋白点。本研究采用一维凝胶电泳的方法,有效避免了双向电泳试验过程复杂、操作步骤多、重复性差和疏水性膜蛋白检测效率低的问题[35-37]。同时,本研究利用分级质谱在NIL-Y叶片类囊体中鉴定出多达1 730种蛋白,其中在KEGG中有功能注释的496种,未知功能蛋白1 234种;在NIL-G叶片类囊体中鉴定到2 170种蛋白,有功能注释的583种,未知功能蛋白1 587种。与前人相比,鉴定到类囊体蛋白数目最多,且利用近等基因系研究植物类囊体尚未见报道,为大豆类囊体蛋白的研究提供了理论基础。
4 结论
利用质谱技术鉴定1对近等基因系大豆叶片类囊体蛋白,NIL-G叶片类囊体特异蛋白1 140种,NIL-Y叶片类囊体特异蛋白700种。其中,NIL-G叶片类囊体蛋白涉及光合作用和天线蛋白的共有20种,参与PSⅡ、ATPase、PSⅠ和光合电子传递链;NIL-Y叶片类囊体蛋白中参与光合作用和天线蛋白的共有7种,仅涉及PSⅡ和ATPase。
猜你喜欢
今日农业(2022年16期)2022-11-09
科学(2022年4期)2022-10-25
中国化肥信息(2022年5期)2022-08-30
今日农业(2021年20期)2021-11-26
今日农业(2021年14期)2021-10-14
食品安全导刊(2021年20期)2021-08-30
Coco薇(2016年5期)2016-06-03
当代化工研究(2016年5期)2016-03-20
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
