
木竹的花器官形态与解剖结构研究
2021-07-28 03:52崔允姬王曙光
南京林业大学学报(自然科学版) 2021年4期
杨 南,崔允姬,王 茜,王曙光*
(1.西南林业大学,云南省高校丛生竹生物学重点实验室,云南 昆明 650224;2.西双版纳国家级自然保护区管护局,云南 景洪 666100;3.山东省临沂市城市管理综合服务中心,山东 临沂 276000)
竹子极少开花,且开花周期特别长,繁殖方式多以无性繁殖为主,在自然条件下大部分竹种需经历120 a或更长的时间才能开花[1]。多数竹子开花后,往往会大面积干枯与死亡,结实率低,在经济和生态上都会造成巨大的损失[2-4]。竹类植物这种开花生理特性,不仅给竹类有性生殖和胚胎发育研究工作带来困难,也极大地限制了竹类植物花器官的形态、分类与遗传学等方面的研究[5],还使得许多竹子标本缺乏花果部分,并且一些竹子种类在发表时也仅仅根据营养体器官而缺乏繁殖器官描述,导致争议很大,给竹子分类学研究及修订造成了困难[1,6]。故竹子的花部特征在分类学研究中具有重要意义。
近30年来,我国发现的竹子开花现象相对较多,对竹子花形态结构方面的研究报道也日益增多,自1984年乔士义等[7]通过对毛竹(Phyllostachysedulis)进行胚胎学观察,初步揭示了其胚胎学特征以来,到目前为止,已较详细报道的竹子花形态与解剖结构的竹种有,爬竹(Drepanostachyumscandeus)、雷竹(Phyllostachyspraecox)、巨龙竹(Dendrocalamussinicus)、月月竹(Chimonobambusasichuanensis)、鹅毛竹(Shibataeachinensis)、异叶苦竹(Arundinariasimoniif.heterophylla)、孝顺竹(Bambusamultiplex)、青丝黄竹(Bambusaeutuldoides)和绵竹(Bambusaintermedia)[8-16]等多个竹种,且在大部分竹种中均发现了雄性败育的现象。2000年,杜凡等[17]对 15 年来在云南观察到的 61 种23 属的竹子进行开花结实现象的报道,但仍然有大量竹种缺乏花器官的形态与解剖结构描述。对竹子花的形态与解剖结构进行解剖与比较研究,在生产与理论上均具有重要科学意义,也为竹子的系统分类研究提供理论参考信息。
木竹(Bambusarutila)又名扁担竹,属箣竹属(Bambusa)竹种,壁厚而坚实,广泛分布于福建、广东、广西、四川、江西等地[18]。竹子花器官由于营养生长周期长、极少开花等生理学特性,难以收集,目前国内外尚未有对木竹花器官进行形态与解剖结构研究的详细报道。笔者通过对木竹不同发育程度的小穗和小花进行解剖观察,分析其胚胎发育特点和规律,探究木竹结实率低的原因,为木竹胚胎学研究及竹类植物的有性育种与分类工作提供参考。
1 材料与方法
1.1 材料的采集与固定
以木竹不同发育程度小穗和小花为实验材料,木竹小穗于2017年7月采自西南林业大学竹种园(102°10′~103°40′E,24°23′~26°22′N)。该竹丛于1991年西南林业大学竹园建园开始时由薛纪如先生等引入竹种园,具体引种地已不可考。该竹丛于2016年秋季开始开花,开花持续两年后死亡。将采集的木竹不同发育程度小穗,用50%(体积分数)FAA固定液固定,抽真空备用。
1.2 小穗、小花形态结构观察
选取30枚不同发育程度的小穗,用直尺测量小穗长度,统计每小穗小花个数,而后使用体视解剖镜(Olympus HO11)进行解剖并拍照。随后将竹花的各部分分离,并分别进行形态观察、测量、拍照,最后进行系统的形态学描述。
1.4 子房、花药石蜡切片观察
将解剖后发育到不同程度的子房和花药进行浸蜡、包埋、石蜡切片,切片厚度7 μm左右,番红-固绿双重染色,中性树脂封片,待切片干固后,置于光学显微镜下观察并拍照。
2 结果与分析
2.1 木竹小穗和小花形态结构观察
木竹的花序比较复杂,其花序轴是由不同的节和营养枝生长发育而来,小穗顶生或腋生。在木竹花期的初期,其花枝的顶端及节部往往只生1枚小穗,穗柄短(图1A),小穗基部由1枚先出叶包裹(图1D)。中后期由于小穗柄基部不断发育出新的小穗,表现为多个小穗呈簇状长在花枝节上和顶端(图1B)。木竹小穗基部的小花先开放,开花顺序由形态学的下端向上端依次开放(图1D),顶端的小花通常败育,因此小穗具有总状花序的特征,但小穗基部具有1个或数个苞片(图1B),有些苞片内具有明显的潜伏芽(图1E),可产生新的小穗,因此又具有有限花序的特征,可知,木竹小穗属于一种混合花序。此外,木竹开花的小穗可见较多的昆虫(图1C),未能明确为害虫还是授粉昆虫,需要进一步开展相关研究。

A. 花期初期early stage of blooming period;B. 花期中期middle stage of blooming period;C. 小穗上的昆虫(箭头所示) insect on spikelets (arrow);D.正在开花的小穗,显示小穗基部的先出叶(箭头所示)blooming spikelet, showing the prophyll at the base lawow (arrow);E. 基部的苞片,可见潜伏芽(箭头所示) floret showing bracts and latent buds lawow (arrow);F. 开放的小花 blossoming flower;G. 小穗轴 rachillas of spikelet;H. 小花的解剖构成 anatomical structure of floret;I.外稃 lemma;J. 内稃palea;K. 浆片 lodicule;L. 雄蕊 stamens;M. 褐化小花,显示未成熟的种子browning floret, showing immature seed;N. 雌蕊 pistil;O. 种子 seeds。
成熟小穗平均长4.756 cm,宽4.0~5.0 mm,成熟小穗苞片及外稃尖端呈紫红色,少数呈嫩绿色(图1D、1F)。小穗轴分节,轴节间极短,长约2.9 mm,小穗轴中空,外被绒毛(图1G)。每个小穗上具5~12朵小花,花开放时花药悬垂于小穗外(图1B—1D,1F)。成熟小花由外向内依次为1枚外稃、1枚内稃、6枚雄蕊、3枚浆片、1枚雌蕊组成(图1H)。外稃呈长圆状披针形,质地较硬(图1I);内稃具两脊,两脊上部被纤毛,两脊间一般4脉(图1J)。成熟的小花(位于小穗下部)外稃稍短于内稃,未成熟的小花(位于小穗中部和上部)外稃长于内稃,且越向上内外稃长度差异越大。木竹小花浆片呈卵圆形,半透明状,上部边缘具流苏状纤毛,偏紫红色,不同浆片大小接近(图1H、1K)。
木竹雄蕊幼时白色,花药底部着药,基部具二尖,顶部也具二尖(图1H、1L)。成熟的花药长6.0~6.5 mm。呈淡黄色,尖部略带紫色(图1D、1F),散粉后花药呈褐色且逐渐脱落(图1M)。木竹小花为6枚雄蕊包围1枚雌蕊,子房长2.5~3.0 mm,具棱,且上宽下窄,上部具绒毛,花柱短,柱头长,羽毛状3分枝柱头,呈紫红色(图1D、1H和1N)。开花时,一般由浆片吸水膨胀而撑开外稃,此时雄蕊露出小花,因此属于开放型花,子房与花药同时伸出,花药则由于花丝伸长而垂悬于小花外,因此木竹的小花属于雌雄异位、雌雄同熟型(图1D和1F)。木竹结实率极低,在本研究所有材料中也只发现1粒果实,颖果,深黄色,种子上部被纤毛(图1O),该纤毛应为子房上部纤毛的残留。
2.2 木竹的雄蕊发育
由小花的横切面可以观察到,雄蕊6枚花丝的所有薄壁细胞均具有浓厚的细胞质和细胞核,花丝内的维管束细胞与其他薄壁细胞结构差异并不显著(图2A)。由于观察到的木竹开花数量较少,未能采集到所有发育阶段的小穗,因此从孢原细胞到初生造孢细胞的各个阶段没有观察到。通过对采集到的材料进行观察,发现同一朵小花的6枚花药在进入减数分裂期前的阶段为同步发育。各花药室的细胞均处于次生造孢细胞时期(图2B、2C)。该时期,造孢细胞体积较大,排列紧密,多为不规则的多面体形,不同药室内的造孢细胞的数量不完全相同(图2C)。此时,不同花药的4层花药壁均已分化完成,由外向内依次为表皮、药室内壁、中层、绒毡层(图2C、2D)。表皮细胞具有浓厚的细胞质和显著的细胞核。药室内壁紧邻表皮细胞,木竹的药室内壁为1层细胞,木竹中层也仅有1层细胞,均具有浓厚的细胞质和细胞核,且形状均较为扁平。次生造胞细胞时的绒毡层细胞质浓厚,部分具两核(图2D)。由于随机取材观察到此时期次数较多,说明次生造孢时期在花药发育过程中所持续的时间较长。
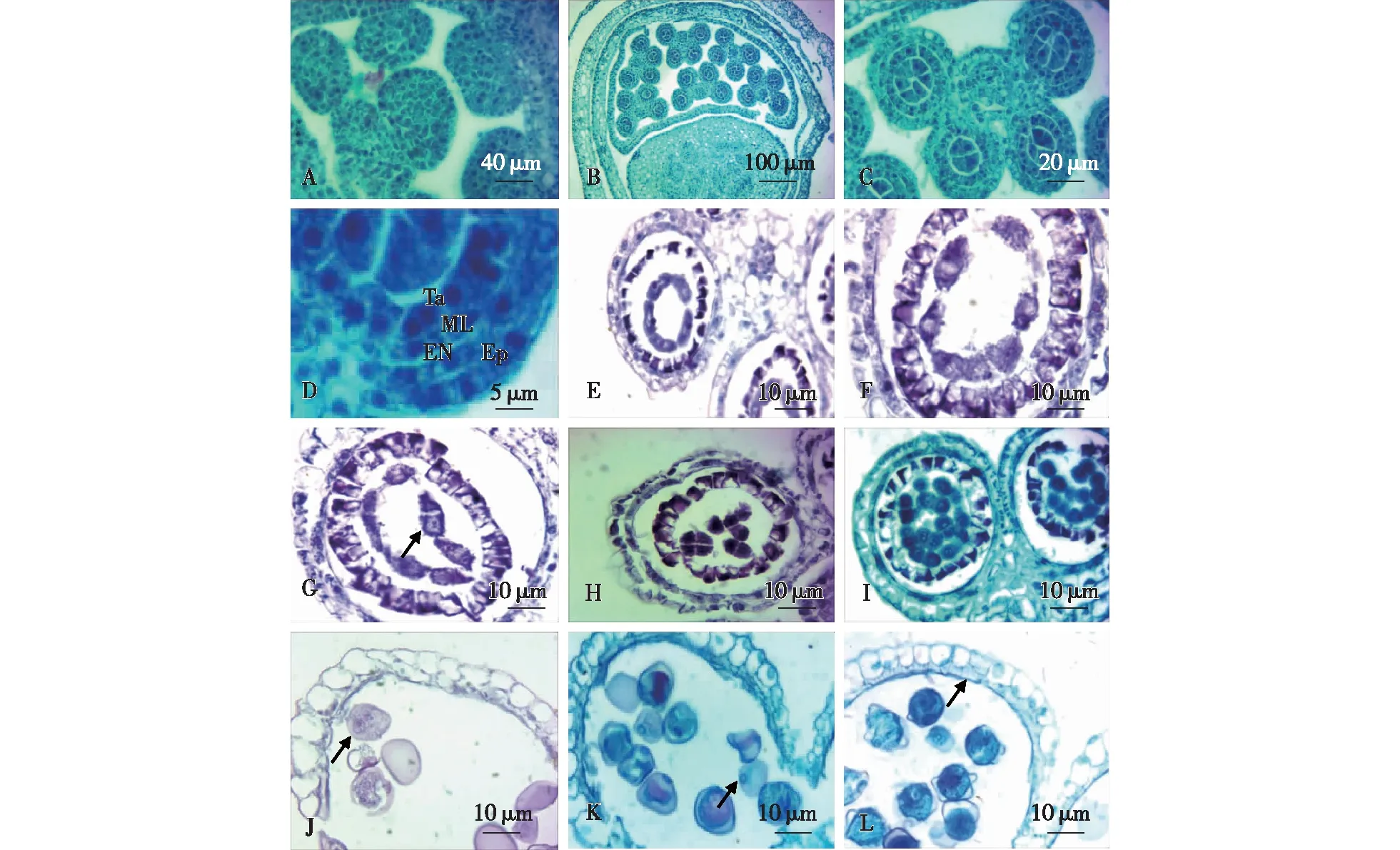
A.6枚花丝横切面 the cross sections of six filaments;B.6枚花药(造孢细胞时期)。The six anthers were all in the stage of secondary sporogenous cell;C. 次生造孢细胞secondary sporogenous cells period;D.次生造孢细胞时期的花药壁 the anther wall during secondary sporogenous cells(Ep.表皮细胞,EN.药室内壁endothecium;ML.中层middle layer;Ta.绒毡层tapetum secondary sporogenous cell);E.小孢子母细胞 microspore mother cell;F和G.小孢子母细胞减数分裂期 the meiosis stage of microspore mother cells;H.二分体和四分体共存 the stage of dyad and tetrad;I.单核小孢子时期 monokaryotic microspore;J. 二细胞花粉粒 two-celled pollen;K. 萌发孔(箭头所示) germinal aperture (arrow);L. 纤维层(箭头所示) fiber layer (arrow)。
次生造孢细胞发生游离,进入小孢子母细胞时期(图2E)。小孢子母细胞体积增大近椭圆形,细胞核大,细胞质浓,无明显液泡,此时花药壁的中层已消失不见(图2E)。此阶段从花药室的横切面上看,花药室的空间较大,可能是花药继续增大和药壁中层分解消失的结果。随后小孢子母细胞进入减数分裂期,形成二分体和四分体(图2F—2H),在木竹花药中观察到二分体和四分体共存于同一个花粉囊(图2H),说明在同一个花粉囊中存在小孢子母细胞减数分裂不同步的情况。木竹四分体4细胞排列在同一平面上故其分裂方式为连续型,其排列方式为左右对称型。通常四分体时期比较短暂,4个小孢子彼此分开释放到药室中去(图2I)。
小孢子是雄配子体的第一个细胞,小孢子从四分体释放后即可称之为花粉粒。此时小孢子仅具有1个细胞核,即单核花粉粒时期(图2I)。由四分体游离出来的小孢子有浓厚的细胞质和位于中央的细胞核,此时绒毡层细胞已明显变小(图2I)。木竹成熟的花粉粒为二细胞花粉粒(图2J),即为成熟花粉粒。花粉粒成熟后具有凸透镜状萌发孔(图2K)。此时药室内壁发生了显著的纤维化,形成纤维层(图2L)。绒毡层彻底退化,仅遗留一薄层状结构贴附于药室内壁。因此,木竹的绒毡层为分泌型绒毡层,成熟花药为两层药壁。
在木竹雄蕊的发育成熟过程中,发现大量花药不能正常发育(图3A),可将其归纳为不同败育类型。第1种类型为花药壁完全发育异常,且花药壁与造孢细胞完全皱缩(图3B)。第2种类型为花药的维管束、药壁和药隔皆正常发育,少数花粉粒空壳,不具细胞质和细胞核(图3C)。此外,还可观察到有的花药室中大量花粉粒皱缩变形,但花药壁和维管束的发育正常(图3D、3E)。
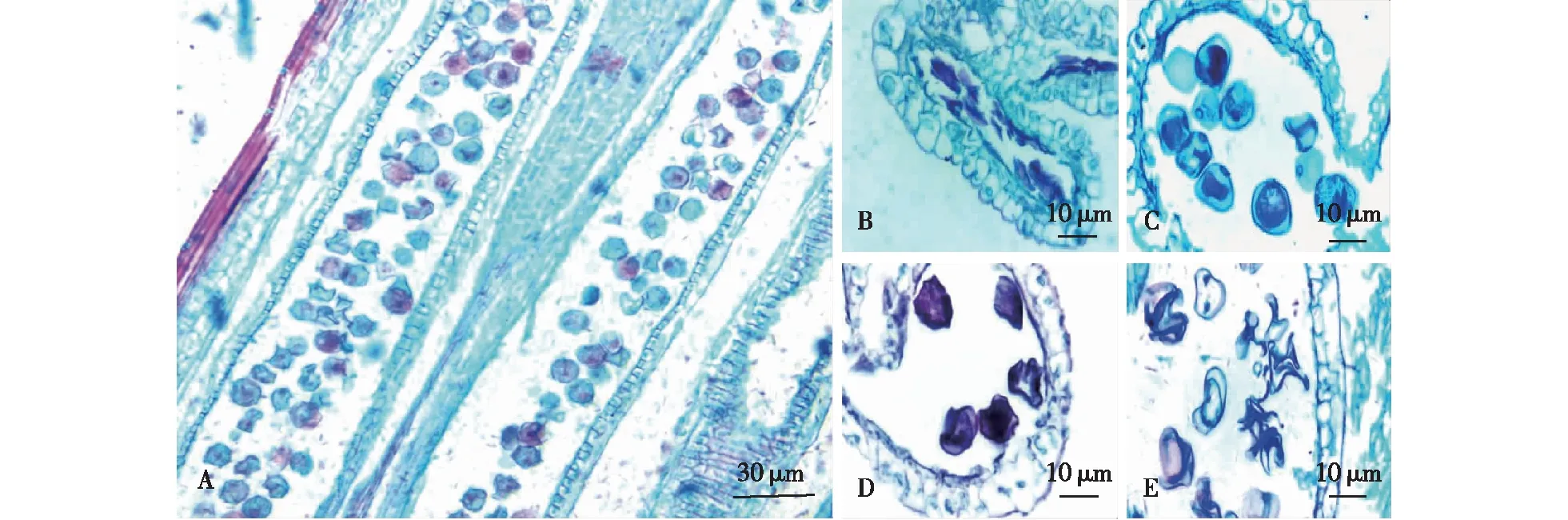
A.纵切的花药室 longitudinal section of pollen sac;B. 小孢子与花药壁均发生收缩 deformed microspore and anther;C.花粉粒无原生质体(箭头所示) pollen grains without protoplast (arrow);D和E.小孢子发生异常收缩 abnormal shrinkage of microspore。
2.3 木竹雌蕊发育
木竹子房是在雌蕊下端的膨大部分,外面被有较硬的纤毛,且有棱边。子房1室,两层珠被,外珠被和内珠被。胚珠与子房侧壁合生,珠孔朝向子房基部,因此属于倒生胚珠(图4A、4B)。木竹子房的柱头为3分枝柱头,暗示其子房为3心皮合生,因此木竹花为侧膜胎座,而非边缘胎座。由于仅有1丛木竹开花,受采集材料的数量限制,仅观察到孢原细胞(图4C)和大孢子母细胞(图4D)以及成熟胚囊阶段(图4E)。成熟胚囊显示了反足细胞、中央细胞及卵器等结构。此外也观察到了正在发育的胚(图4F)。因此,木竹雌蕊的发育是正常的。
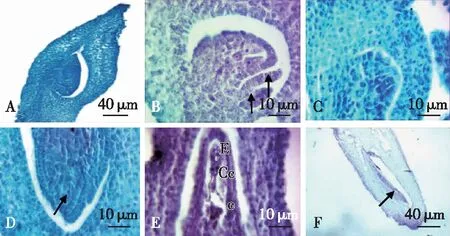
A.子房1室,侧膜胎座。The ovary is unilocular,parietal placenta and the ovule was anatropous;B.双层珠被(箭头所示) dual-integument (arrow);C和D. 孢原细胞(箭头所示) archesporial cells (arrow);E. 成熟胚囊mature embryo sac(Ea. 卵器 egg apparatus;Cc.中央细胞central cells;Ac.反足细胞antipodal cells);F. 发育的胚(箭头所示) developing embryo corrow (arrow)。
3 讨 论
竹类植物的花器官主要为小穗,小穗是构成其花序的基本单位[19]。不同竹类植物小穗形态差异巨大,经笔者观察分析、查阅相关文献[20]以及与同行交流发现,在合轴丛生这一类型,如箣竹属、牡竹属等近源属的竹种中,不同类型竹种的小穗长度差异巨大。小穗可大致总结为两种类型,一种小穗较长,小穗轴节间长,外观明显可见,小花交互排列在小穗轴的两侧,如箣竹属的车筒竹(B.sinospinosa)[21]、孝顺竹[22]、青丝黄竹[15]、绵竹[16]、硬头黄竹(B.rigida)[23]等竹种,笔者观察到的木竹小穗也属于这一类型。另外一种类型的小穗外观形态短扁,小穗轴节间极度短缩,小花紧密排在小穗轴的两侧[18]。绿竹(B.oldhamii)[20]、慈竹(B.emeiensis)[21]以及牡竹属的竹种如巨龙竹[10]、麻竹(D.latiflorus)[24]等竹种,多属于这一类。
不同类型竹种在同一小穗上的开花先后是不一致的。林树燕等[20]根据目前的相关研究报道,将其总结为3种类型:①小穗基部小花先开放;②小穗顶部小花先开放;③小穗中部小花先开放。笔者观察到的木竹即属于小穗基部小花先开放类型。林树燕等[20]认为竹子的小穗各小花的发育是由基部向顶部发育的,属于总状花序,但小穗的基部具有潜伏芽,又属于有限花序的特征,因此认为竹类植物花序基本单位为小穗,花序应统一为混合花序。木竹小穗基部的小花先开放,小穗基部具有潜伏芽,支持林树燕等[20]的观点,属于混合花序。
竹子的小花一般由外稃、内稃、浆片、雌蕊和雄蕊5部分组成,它们在数量、形态和颜色上会因竹种的不同而略有变化。在本研究中,观察到木竹的小花也是由这5部分组成,即外稃1枚,内稃1枚,浆片3枚,雌蕊1枚,雄蕊6枚。浆片的主要生理功能通常为开花时,浆片强烈吸水膨胀,将内外稃撑开,帮助雄蕊伸出小穗外[15-16]。因此根据内外稃在开花时是否显著张开,将竹子的小花分为开放型和闭合型两类。开放型花在箣竹属中常见,开花时,浆片膨胀,内外稃张开。闭合型则以牡竹属为代表,其小花不具有浆片,开花时稃片不张开。慈竹[21]、绿竹[20]等竹种虽然具有浆片但并不属于开放型,反而属于闭合型。闭合型小花,如慈竹[21]、麻竹[24]、巨龙竹[10]等竹种,开花时,通常雌蕊先伸出小穗外完成授粉,为典型的雌雄异熟或雌蕊先熟。木竹属于箣竹属竹种,为典型的开放型小花,开放时,内外稃张开,雌雄同熟,雄蕊由于极度伸长的花丝而悬垂于小穗外,远低于雌蕊柱头的位置。从进化的角度看,这种雌雄异位也是一种进化,并有利于异花授粉的繁育机制,但在只有极少数个体开花的情况下,则对繁育下一代极其不利,这可能是竹子结实率低的重要原因[15]。
自然条件下,竹类植物不常开花,且开花后结实率低,造成这一现象的原因是多方面的。除了雌雄异位、开花个体少,容易导致授粉率、结实率低以外,雄配子体发育异常也是导致竹类植物结实率低的重要原因。竹类植物的有性生殖过程中,雄配子体发育异常的现象常见,雌配子体大都发育正常,在木竹花药中发现大量发育异常的花粉粒,包括药囊与其中的小孢子严重收缩变形,花粉粒无原生质体以及药囊壁发育正常但小孢子异常收缩等现象。类似的败育现象在爬竹[8]、青丝黄竹[15]、鹅毛竹[12]和龙竹(D.giganteus)[21]、车筒竹[21]和绵竹[16]、新小竹(Neomicrocalamusprainii)[25]、‘霞早’绿竹[26]中也观察到。本研究在所有采集到的木竹小穗中仅获得1枚未成熟的种子,因此雌雄异位、异花授粉、开花个体少,部分雄配子体发育异常是导致木竹结实率低的主要原因。
木竹小穗基本结构发育正常,为混合花序,小穗基部具潜伏芽,每个小穗5~12朵小花,下部小花先开放,成熟小花呈紫红色或嫩绿色。完整小花由外稃1枚、内稃1枚、浆片3枚,雄蕊6枚和雌蕊1枚构成,羽毛状柱头3分支,开放型小花,雌雄同熟且异位,异花授粉。花药具有4药室。次生造孢细胞时,花药壁由外到内依次为表皮、药室内壁、中层和绒毡层,绒毡层腺质型。减数分裂期时,不同造孢细胞分裂不同步。花药成熟后纵裂散粉,花药壁仅有表皮和纤维层,绒毡层退化。成熟花粉粒为2细胞型,且具有1个萌发孔。子房外型具棱,且上宽下窄,上部具绒毛,1室,上位,侧膜胎座,倒生胚珠,胚囊发育正常,能产生少量种子。花药出现皱缩、小孢子变形及空洞等现象及部分雄配子体发育异常等多个因素导致木竹花药出现大量败育现象。
参考文献(reference):
[1]易同培,史军义,马丽莎.中国竹类图志[M].北京:科学出版社,2008.YI T P,SHI J Y,MA L S.Iconographia bambusoidearum sinicarum[M].Beijing:Science Press,2008.
[2]李睿,章笕,章珠娥.中国竹类植物生物多样性的价值及保护进展[J].竹子研究汇刊,2003,22(4):7-12,17.LI R,ZHANG J,ZHANG Z E.Values of bamboo biodiversity and its protection in China[J].J Bamboo Res,2003,22(4):7-12,17.DOI:10.3969/j.issn.1000-6567.2003.04.002.
[3]李洁.孝顺竹的生殖生物学研究[D].南京:南京林业大学,2013.LI J.Studies on the reproductive biology ofBambusamultiplex[D].Nanjing:Nanjing Forestry University,2013.
[4]林树燕,石文文,缪彬彬,等.竹类植物生殖生物学研究进展[J].世界竹藤通讯,2010,8(2):1-6.LIN S Y,SHI W W,MIU B B,et al.Research advances in reproduction biology of bamboos[J].World Bamboo Rattan,2010,8(2):1-6.DOI:10.3969/j.issn.1672-0431.2010.02.001.
[5]侯丹.毛竹生殖器官发育相关miRNA挖掘与miR159-GAMYB途径对花药发育调控研究[D].北京:中国林业科学研究院,2018.HOU D.Identification of miRNAs involved inPhyllostachysedulisreproductive organs and regulatory role of miR159-GAMYB pathway in anther development[D].Beijing:Chinese Academy of Forestry,2018.
[6]彭晟,杨汉奇,李德铢.竹亚科箭竹属两种植物花序的补充描述[J].云南植物研究,2006,28(3):257-258.PENG S,YANG H Q,LI D Z.Supplementary description of inflorescences for two species ofFargesia(Bambusoideae)[J].Acta Bot Yunnanica,2006,28(3):257-258.DOI:10.3969/j.issn.2095-0845.2006.03.005.
[7]乔士义, 廖光庐. 毛竹的胚胎发育观察[J]. 竹类研究, 1984, 3(1): 15-22. QIAO S Y,LIAO G L.Embryology observation ofPhyllostachysedulis[J]. Bamboo Research,1984,3(1): 15-23.
[8]庞延军,喻富根,胡成华,等.爬竹雄蕊发育异常的初步观察[J].竹子研究汇刊,1994,13(4):42-46.PANG Y J,YU F G,HU C H,et al.Preliminary observation on abnormal development of the stamens ofDrepanostachyummicrophyllum[J].J Bamboo Res,1994,13(4):42-46.
[9]黄坚钦,黄华宏,何福基,等.雷竹的小孢子发生和雄配子体形成[J].竹子研究汇刊,1999:55-58.HUANG J Q,HUANG H H,HE F J,et al.The formation of microspore and the development of male gametophyte ofPhyllostachyspraecox[J].J Bamboo Res,1999:55-58.
[10]王曙光,普晓兰,丁雨龙.巨龙竹生殖器官形态结构及雌、雄配子体的发育[J].植物研究,2006,26(3):270-274.WANG S G,PU X L,DING Y L.The structures of reproductive organs and development of the female and male gametophyte ofDendrocalamussinicus[J].Bull Bot Res,2006,26(3):270-274.DOI:10.3969/j.issn.1673-5102.2006.03.006.
[11]林树燕,郝娟娟,辛华,等.月月竹大、小孢子发生和雌、雄配子体发育研究[J].南京林业大学学报(自然科学版),2009,33(3):9-12.LIN S Y,HAO J J,XIN H,et al.The megasporogenesis,microsporogenesis and the development of their female and male gametophyte inMenstruocalamussichuanensis[J].J Nanjing For Univ (Nat Sci Ed),2009,33(3):9-12.DOI:10.3969/j.issn.1000-2006.2009.03.003.
[12]林树燕,丁雨龙.鹅毛竹大小孢子及雌雄配子体发育[J].西北植物学报,2012,32(5):907-914.LIN S Y,DING Y L.Development of the male and female gametophytes inShibataeachinensis(Bambusoideae)[J].Acta Bot Boreali-Occidentalia Sin,2012,32(5):907-914.DOI:10.3969/j.issn.1000-4025.2012.05.010.
[13]林树燕.鹅毛竹和异叶苦竹的生殖生物学研究[D].南京:南京林业大学,2009.LIN S Y.Studies on the reproductive biology ofShibataeachinensisandArundinariasimoniif.albostriatus[D].Nanjing:Nanjing Forestry University,2009.
[14]林树燕,李洁,赵荣,等.孝顺竹花芽分化及小孢子发生与雄配子体发育[J].南京林业大学学报(自然科学版),2015,39(4):51-56.LIN S Y,LI J,ZHAO R,et al.The development of flowering bud differentiation and male gametophyte ofBambusamultiplex[J].J Nanjing For Univ (Nat Sci Ed),2015,39(4):51-56.DOI:10.3969/j.issn.1000-2006.2015.04.009.
[15]唐国建,杨金梅,丁雨龙,等.青丝黄竹花形态与结构研究[J].南京林业大学学报(自然科学版),2016,40(2):71-75.TANG G J,YANG J M,DING Y L,et al.Studies on the flower morphology and structure inBambusaeutuldoidesMcClure var.viridi-vittata(W.T.Lin) Chia[J].J Nanjing For Univ (Nat Sci Ed),2016,40(2):71-75.DOI:10.3969/j.issn.1000-2006.2016.02.012.
[16]王雨珺,罗剑,陈楠楠,等.绵竹花形态结构及雌、雄配子体的发育研究[J].植物研究,2017,37(4):492-498.WANG Y J,LUO J,CHEN N N,et al.Floral morphology and development of female and male gametophyte ofBambusaintermediaHsueh et Yi[J].Bull Bot Res,2017,37(4):492-498.DOI:10.7525/j.issn.1673-5102.2017.04.002.
[17]杜凡,薛嘉榕,杨宇明,等.15年来云南竹子的开花现象及其类型研究[J].林业科学,2000,36(6):57-68.DU F,XUE J R,YANG Y M,et al.Study on flowering phenomenon and its type of bamboo in Yunnan in past fifteen years[J].Sci Silvae Sin,2000,36(6):57-68.DOI:10.3321/j.issn:1001-7488.2000.06.009.
[18]王正平, 叶光汉, 杨雅玲, 等.中国植物志:第九卷, 第一分册[M]. 北京:科学出版社, 1996.
[19]耿伯介.试论竹类的花序及其演变[J].武汉植物学研究,1986,4(4):323-336.KENG P.A preliminary study of the inflorescence type arising from bamboos and its variation[J].J Wuhan Bot Res,1986,4(4):323-336.
[20]林树燕,万雅雯,傅华君,等.竹类植物花序建成及花序类型修正[J].南京林业大学学报(自然科学版),2018,42(6):1-6.LIN S Y,WAN Y W,FU H J,et al.Research on inflorescence establishment and revision of inflorescence type in bamboo plants[J].J Nanjing For Univ (Nat Sci Ed),2018,42(6):1-6.DOI:10.3969/j.issn.1000-2006.201801053.
[21]WANG S G,PU X L,LIN S Y,et al.Reproductive characteristics of three bamboo species[J].Pak J Bot, 2015,47(6): 2301-2308.
[22]林树燕,李洁,赵荣,等.南京地区孝顺竹的开花生物学特性研究[J].南京林业大学学报(自然科学版),2015,39(2):52-56.LIN S Y,LI J,ZHAO R,et al.A research on the flowering biological characteristics ofBambusamultiplexin Nanjing City[J].J Nanjing For Univ (Nat Sci Ed),2015,39(2):52-56.DOI:10.3969/j.issn.1000-2006.2015.02.009.
[23]李娟,王一方,初彩华,等.硬头黄竹花的形态与结构研究[J].林业科学研究,2020,33(1):28-34.LI J,WANG Y F,CHU C H,et al.Studies on flower morphology and structure ofBambusarigida[J].For Res,2020,33(1):28-34.DOI:10.13275/j.cnki.lykxyj.2020.01.004.
[24]钟远标,岳晋军,楼崇,等.麻竹的花器官与繁育系统[J].林业科学,2017,53(1):1-10.ZHONG Y B,YUE J J,LOU C,et al.Floral organ and breeding system ofDendrocalamuslatiflorus[J].Sci Silvae Sin,2017,53(1):1-10.DOI:10.11707/j.1001-7488.20170101.
[25]初彩华,黄玲,王曙光.新小竹花形态结构及雌、雄配子体的发育研究[J].西北植物学报,2019,39(5):763-769.CHU C H,HUANG L,WANG S G.Floral morphology and development of female and male gametophytes ofNeomicrocalamusprainii[J].Acta Bot Boreali-Occidentalia Sin,2019,39(5):763-769.DOI:10.7606/j.issn.1000-4025.2019.05.0763.
[26]林树燕, 傅华君, 万雅雯, 等. ‘霞早’绿竹花形态特征及花药发育的组织学观察. 南京林业大学学报 (自然科学版), 2019, 43(2):7-13. LIN S Y, FU H J, WAN Y W, et al. Anther development and floral morphology characteristics ofBambusaoldhami‘Xia Zao’ZSX[J].J Nanjing For Univ (Nat Sci Ed),2019,43(2):7-13. DOI:10.3969/j.issn.1000-2006.201711031.
猜你喜欢
国际种业前沿动态(2020年18期)2020-12-23
农业科技与信息(2020年6期)2020-12-18
农业科技与信息(2020年8期)2020-12-18
纺织科学研究(2017年3期)2017-05-17
世界竹藤通讯(2015年4期)2015-12-29
世界竹藤通讯(2015年4期)2015-12-29
河南科技(2014年5期)2014-02-27
江西农业大学学报(2013年3期)2013-11-05
延边大学学报(自然科学版)(2011年2期)2011-01-15
世界竹藤通讯(2010年2期)2010-08-15
