
过氧化物酶体增殖物激活受体β(δ)对肌纤维类型转化和肉品质的影响及其调控因素研究进展
2021-09-06 12:34侯艳茹
动物营养学报 2021年8期
孙 冰 侯艳茹 苏 琳 靳 烨
(内蒙古农业大学食品科学与工程学院,呼和浩特 010018)
肌纤维作为组成肌肉组织的基本结构单位,其类型组成、比例分布及生物学特性与畜禽的生长发育和肉品质紧密相关[1]。不同类型的肌纤维具有不同的特性,氧化型肌纤维的电子传递链含有较高含量的肌血球素和肌红蛋白,具有更高的氧化能力和较低的肌球蛋白ATP酶活性。相反,酵解肌纤维具有更高的ATP酶活性和少量的糖原。肌肉纤维类型组成与肌肉的形态特征有直接关系,是改善肌肉功能的重要研究对象。氧化型肌纤维在肌肉组织中占比高时,肌肉大理石花纹丰富、色泽鲜红、细嫩多汁且保水性较好。酵解型肌纤维在肌肉组织中占比高时,宰后过程中会导致pH降低和蛋白质结构退化,更易产生白肌肉(PSE肉)[2]。因此,长期以来,通过调控肌纤维的类型组成来改善畜禽肉品质是国内外学者研究的热点。
不同类型肌纤维在完成转化过程中涉及一些复杂的生物学信号通路,骨骼肌具有高度可塑性,机体在生长发育进程中或受到某些外界刺激时,细胞内相关的信号通路就会发挥调控作用,激活一些特异性因子的表达,进而完成转化。不同类型的肌纤维之间遵循以下转化路径:Ⅰ型(慢收缩氧化型)↔Ⅱa型(快收缩氧化酵解型)↔Ⅱx/d型(中间型)↔Ⅱb型(快收缩酵解型)。骨骼肌的肌纤维类型转化受多种转录因子调控,其中过氧化物酶体增殖物激活受体β(δ)[peroxisome proliferator-activated receptor β(δ),PPARβ(δ)]较为重要[3-4]。目前过氧化物酶体增殖物激活受体(peroxisome proliferator activated receptors,PPARs)有3种结构亚型被相继发现,并分别命名为过氧化物酶体增殖物激活受体α(peroxisome proliferator-activated receptor α,PPARα)、PPARβ(δ)、过氧化物酶体增殖物激活受体γ(peroxisome proliferator-activated receptor γ,PPARγ)。这3种结构亚型由不同功能的基因编码而成,因其在分布和组织上不同的表达,功能也略有差异。相比于其他2种亚型,PPARβ(δ)在各种组织中都广泛表达,尤其在骨骼肌中[5]。PPARβ(δ)作为生物体内重要的核受体,其所引起的核内转录变化对肌纤维类型的转化和肌肉有氧耐力的提高有着关键的调控作用。本文主要对PPARβ(δ)的生物学功能、PPARβ(δ)对肌纤维类型转化的影响及其内在调控机制、激活PPARβ(δ)的因素以及其与肉品质的关系进行综述,以期为今后通过调控肌纤维类型组成,进而改善畜禽肉品质提供新思路。
1 PPARβ(δ)概述
作为新型的固醇类核受体,PPARβ(δ)可调节目的基因的表达,其所编码的蛋白质与生物体内其他核受体具有极为相似的功能区和结构区[6]。PPARβ(δ)的结构域由A~F 6个结构区组成,如图1所示。结构域A/B区位于N端,该区功能主要是作为反式调控元件及调节其与DNA相结合的能力。C区称为DNA结合域(DNA-binding domain,DBD),能特异性识别靶基因序列,实现与靶基因的结合,与DNA上相匹配的反应元件结合,从而完成对基因转录过程的调节。D区称为铰链区(hinge),主要调节转录活性,易形成异聚体,并通过与DNA的结合连接DNA结合域和配体结合域(ligand biding domain,LBD)。E区位于C端,是配体结合域;E区内部有1个“Y”型的疏水袋,PPARβ(δ)的配体袋较狭窄,PPARα、PPARγ的配体相对较大无法与之结合[7]。PPARβ(δ)通过配体结合域与配体相结合,将自身转化成一种特殊的活性形式,从而能够与DNA结合并调节目的基因表达。除上述功能区域外,在靠近C端还有一段F区,功能尚未见报道[8]。
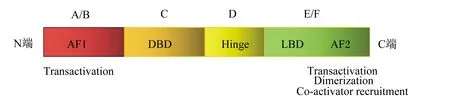
AF1:活化功能1 activation function 1;DBD:DNA结合域 DNA-binding domain;Hinge:铰链区;LBD:配体结合域 ligand biding domain;AF2:活化功能2 activation function 2;Transactivation:转录激活;Dimerization:二聚化;Co-activator recruitment:结合协同激活剂。
E区在核酸定位和受体二聚化过程中起关键作用,受体二聚化过程是PPARβ(δ)在核内发挥活性所必不可少的。通过配体结合域完成与PPARβ(δ)的结合,PPARβ(δ)一旦经配体激活后,转录活性便立即发生改变,可与DNA特定位点结合调节目的基因表达,该位点由一个碱基对分隔的六聚体序列组成,位于靶基因的启动子/增强子区域。配体的结合触发构象变化,导致共阻遏物的释放、辅激活因子的募集和最后的靶基因表达[9]。
2 PPARβ(δ)对肌纤维类型转化的影响及其调控机制
2.1 PPARβ(δ)对肌纤维类型转化的影响
PPARβ(δ)的生物学效应主要体现在能促进脂肪细胞分化和脂肪生成、调节机体内糖脂平衡、提高机体对胰岛素的反应性和抑制炎症基因等方面[10-11]。随着研究的深入,“马拉松鼠”的问世初次揭示了PPARβ(δ)在肌纤维类型转化、线粒体生物发生、提高肌肉有氧耐力等方面发挥着重要作用[12]。因此,PPARβ(δ)对促使肌纤维类型发生转化进而提升肌肉耐力一直备受关注。PPARβ(δ)是肌肉组织中能够驱动功能性Ⅰ型肌纤维形成的转录因子,其激活机制历经复杂的途径,能促使快肌纤维功能和形态特征发生变化,使其向慢肌纤维转化,提高生物体抗疲劳性和改善骨骼肌氧化代谢功能。PPARβ(δ)经激活后能调控肌纤维类型分布并能启动相应的神经调节,使神经末梢产生适应,进而可生成新的运动单位[12]。
Luquet等[13]和Wang等[12]先后建立了肌肉特异性PPARβ(δ)过表达的动物模型以探寻PPARβ(δ)在骨骼肌中的生物学效应,结果表明骨骼肌中PPARβ(δ)过表达时可促使酵解型肌纤维更多的向氧化型肌纤维转化,氧化代谢相关基因表达量显著增加,并且这极大影响了小鼠肌肉的发育和氧化代谢能力。这提示我们PPARβ(δ)可能是骨骼肌中促进线粒体生物发生和肌纤维类型发生转化的调控因子,在肌肉重塑中起到了不容忽视的调控作用,同时PPARβ(δ)在提高骨骼肌耐力的生物学调控作用方面给人以启发。荆海军[14]通过对C2C12成肌细胞株和小鼠原代成肌细胞分化过程进行深入研究,发现PPARβ(δ)的过表达能显著上调氧化型肌纤维的表达量。以上研究结果说明骨骼肌中PPARβ(δ)基因特异性高表达能诱导骨骼肌纤维重塑的部分特征,如增强氧化酶和线粒体标志蛋白的表达。同时,PPARβ(δ)的激活能够诱导骨骼肌收缩力提高[15]。将PPARβ(δ)转基因小鼠放在输氧的封闭跑步机上,它们抗疲劳能力更强,能够维持的时间和距离分别增加了67%和92%;相反,PPARβ(δ)基因敲除小鼠能维持的时间和距离比野生型小鼠均短约30%[12]。以上这些研究结论都提示我们,PPARβ(δ)作为信号转导过程重要的靶点,对促进肌纤维类型转化、线粒体生物发生和提升骨骼肌耐力适应中有着积极的作用。
2.2 PPARβ(δ)对肌纤维类型转化的调控机制
在过去的20多年里,控制肌纤维类型转换的基因调节通路已经被广泛描述,随着近些年研究的不断深入,更多的结论证实核受体在骨骼肌能量代谢和肌纤维类型转换的调节中起着至关重要的作用[16],其中核受体PPARβ(δ)可通过结合其他激活因子、激酶级联等产生多种信号事件,这些信号通过改变机体内关键核受体的转录活性、微小RNA(microRNA,miRNA)的稳定性等,最终诱导骨骼肌纤维类型发生转化。在关于PPARβ(δ)调控肌纤维类型发生转化的过程中,其可能的内在作用机制主要是过氧化物酶体增殖物激活受体γ辅激活剂1α(peroxisome proliferator-activated receptor gamma co-activator 1-α,PGC-1α)/PPARβ(δ)/雌激素相关受体γ(estrogen-related receptor γ,ERRγ)/miRNA途径(图2)。
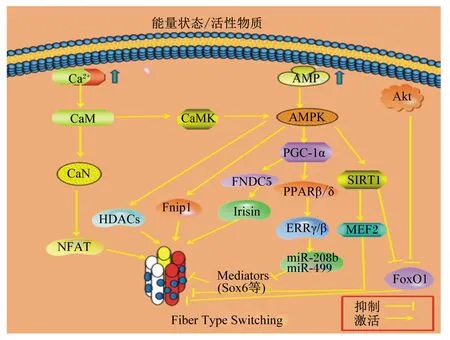
AMPK:腺苷酸活化蛋白激酶AMP-activated protein kinase;SIRT1:沉默信息调节因子1 silent information regulator 1;CaM:钙调素 calmodulin;CaMK:钙调素依赖性蛋白激酶 calmodulin dependent protein kinase;PGC-1α:过氧化物酶体增殖物激活受体γ共激活因子-1α peroxisome proliferator-activated receptor gamma co-activator 1 alpha;MEF2:肌细胞特异性增强结合因子2 myocyte-specific enhancer-binding factor 2;CaN:钙调磷酸酶 calcineurin;HDACs:组蛋白脱乙酰酶 histone deacetylases;NFAT:T细胞核因子 nuclear factor of activated T cells;FNDC5:纤维连结蛋白Ⅲ型域包含蛋白5 fiber links protein Ⅲ domain contains 5;Fnip1:卵泡素互作蛋白1 folliculin interacting protein 1;FoxO1:叉头框转录因子O亚族1 forkhead box transcription factor O1;Irisin:鸢尾素;Akt:蛋白激酶B protein kinase B;PPARβ/δ:过氧化物酶体增殖剂激活受体β/δ peroxisome proliferators-activated receptor β/δ;ERRγ/β:雌激素相关受体γ/β estrogen-related receptor γ/β;Mediators:介质;Fiber Type Switching:纤维类型转化。
PGC-1α是一种转录的协同激活因子,参与肌纤维转化、线粒体生物合成和能量代谢等骨骼肌生理学的多个方面,在Ⅰ型肌纤维中高表达[19]。PGC-1α共激活许多转录因子,包括PPARs和雌激素相关受体(estrogen-related receptor,ERR)、核呼吸因子1/2(nuclear respiratory factor 1/2,NRF1/2)和肌细胞增强因子2(myocyte enhancer factor 2,MEF2)。PGC-1α直接与其转录因子相互作用,通过组蛋白乙酰转移酶活性来募集介导染色质重塑的分子,并通过募集转录机制的成分,如RNA聚合酶Ⅱ来实现基因转录[20]。PGC-1α的表达可由多种生理刺激诱导,如冷暴露、禁食和运动训练,PGC-1α可以协调多个转录因子的活性,以适应各种外部刺激,从而控制肌肉中的代谢和结构适应[21-22]。
小鼠骨骼肌中过表达PGC-1α可导致Ⅰ型肌纤维比例的增加、线粒体代谢标志性基因的表达量上调[23],当小鼠缺失PGC-1α时,会降低其线粒体关键基因表达量和运动氧化能力[24]。Luquet等[13]研究揭示,PGC-1α在没有配体的情况下便可迅速激活PPARβ(δ),也有学者认为PGC-1α对PPARβ(δ)的激活作用是通过蛋白质之间的互作进行的[25]。这说明PGC-1α对肌纤维类型转化的影响极有可能是通过激活其下游效应器PPARβ(δ)来实现的。Lin等[23]通过建立PGC-1α转基因小鼠模型,发现PGC-1α过表达诱导线粒体基因和线粒体酶细胞色素C氧化酶亚基Ⅱ(cytochrome C oxidase Ⅱ,COXⅡ)、细胞色素C氧化酶亚基Ⅳ(cytochrome C oxidase Ⅳ,COXⅣ)基因表达量增加,并且Ⅰ型肌纤维标志基因,如肌红蛋白和肌钙蛋白的表达量也显著增加,说明PGC-1α在高含量下能诱导更完全的肌纤维类型转化;这其中伴随着肌细胞增强因子2(myocyte enhancer factor 2,MEF2)表达量的显著提高,该研究结果揭示在肌纤维转化过程中PGC-1α与MEF2能协同激活转录过程。Krämer等[26]对人体肌肉进行活体组织检查后发现,与运动量正常的人相比,运动员的PGC-1α和PPARβ(δ)mRNA 表达量与骨骼肌中氧化型肌纤维含量变化趋势一致,提示我们PPARβ(δ)可能位于PGC-1α的下游,受PGC-1α的调控,从而影响骨骼肌有氧代谢以及肌纤维类型的转化。PGC-1α的表达受很多因素影响,已确切证实PGC-1α可以与多数核内激素受体包括PPARβ(δ)在内的超家族成员,以不同的方式与其完成分子对接任务,进而调控机体生物学功能[27]。以上结论提示我们今后可以考虑从激活PGC-1α上调PPARβ(δ)的表达,进而以PPARβ(δ)调控肌纤维转化机制为切入点进行研究观察。
miRNA作为基因调控过程的关键角色,这一重要发现开拓了基因表达控制的一个新维度[28]。miRNA是一类保守性极高的非编码RNA,此前已有大量研究证实miRNA在控制骨骼肌发育、肌球蛋白表达和肌纤维表型等方面有重要作用。多数miRNA编码在宿主基因的内含子中,表明它们与其宿主基因共同参与调控过程,miRNA可以通过翻译或基因沉默来调节基因表达[29]。其中有一类内含子miRNA在肌肉中特异性表达并在肌纤维发育及类型转化中起关键作用,被称为肌源性miRNA(myomiRNA),例如miR-1、miR-499-5p、miR-208a、miR-208b和miR-499等,其中典型的慢肌基因miR-208a、miR-208b和miR-499分别是由其相应的肌肉特异性肌球蛋白重链基因(myosin heavy chains,Myh)Myh6、Myh7和Myh7b所编码[30]。
有学者初步预测miR-206可能通过抑制慢肌纤维转录的阻遏物(Sox6)的表达来促进慢肌纤维的形成[31],但尚未得到证实。2009年,Van Rooij等[32]首次提出核受体PPARβ(δ)和ERRγ协同作用能够激活Myh7和Myh7b基因的转录,从而在氧化型肌纤维能量代谢中发挥关键作用。Gan等[16]对从转基因小鼠MCK-PPARs鼠中分离出的miRNA表达谱进行研究并结合基因表达结果发现了PPARs和miRNA之间的联系,ERRγ和miR-208b、miR-499表达量与MCK-PPARβ(δ)小鼠肌肉中Ⅰ型肌纤维标志基因的表达量呈正相关。与此相反,miR-208b、miR-499表达量在MCK-PPARα小鼠肌肉中被抑制,同时缺乏对ERRγ的诱导和对Ⅰ型肌纤维程序的抑制。对骨骼肌细胞功能获得和丧失的研究证实PPARβ(δ)和PPARα在通过miR-208b、miR-499回路调节Ⅰ型肌纤维程序中具有相反的作用。这些结果证实PPARs信号位于miR-208b、miR-499的上游,并且PPARβ(δ)激活而PPARα抑制肌肉中的miRNA回路。然而,在miR-208b、miR-499启动子中没有发现一致的PPARs结合位点,却发现了另一种核受体ERRγ的高度保守位点,ERRγ已被证明在骨骼肌中促进Ⅰ型肌纤维的形成[33];此外,该研究组还发现,ERRγ通过高度保守的ERR反应元件能直接激活Myh7(miR-208b)和Myh7b(miR-499)基因的转录,在这一调控网络中起着关键作用,考虑到ERRγ的表达量在MCK-PPARβ(δ)小鼠肌肉中升高,ERRγ被认为是PPARs和miR-208b、miR-499之间联系的桥梁,PGC-1α在这一调节机制中极有可能通过其对ERRγ和PPARβ(δ)的协同激活效应而起到助推作用[33]。通过后续对小鼠做功能缺失和获得试验,以及结合人体肌肉的组织切片,证明PPARβ(δ)通过ERRγ对miR-499和miR-208b产生刺激作用,确实能够上调慢肌纤维比例[34],Sox6是Sox基因家族的转录因子,是慢肌纤维特异性基因的阻遏物,在骨骼肌分化发育过程中具有重要的作用。在调控肌纤维类型方面,肌肉组织中缺失Sox6基因后,Myh7(miR-208b)的表达量显著提高,酵解型肌纤维在向氧化型肌纤维转化过程中伴随慢肌纤维特异性基因肌钙蛋白1(troponin I type 1,TNNI1)、肌红蛋白(myoglobin)(慢肌纤维中高表达)以及线粒体标志蛋白表达量的显著提高[35]。迄今,有大量研究在大鼠、小鼠、猪和人的试验中证实Sox6是miR-499-5p的下游靶基因[36-40]。
由此可见,PPARβ(δ)通过ERRγ、miRNA网络及Sox6调控骨骼肌慢肌纤维形成的具体信号途径中伴随着复杂多样的作用机制,还需要进行更加深入的研究。
3 激活骨骼肌PPARβ(δ)表达的因素
PPARβ(δ)表达的激活受到多种因素的影响,包括配体和运动,这些因素通过激活PPARβ(δ)高表达进而使肌纤维类型发生转化。
3.1 配体
PPARβ(δ)配体根据来源分为天然配体和人工合成配体2种,其中天然配体包括大部分不饱和脂肪酸、支链脂肪酸等。亚油酸(C18∶2)、α-亚麻酸(C18∶3)和花生四烯酸(C20∶4)等是PPARβ(δ)较为有效的天然配体。根据作用可分为激动剂和拮抗剂,拮抗剂或激动剂与配体结合域结合后,PPARβ(δ)能更加稳定地与协同激动子或协同抑制子结合发挥作用[4]。
3.1.1 不饱和脂肪酸
作为调节骨骼肌脂肪酸代谢的核受体,PPARβ(δ)由不饱和脂肪酸激活。Tumova等[41]研究表明,与几种类型的不饱和脂肪酸(如棕榈酸、亚油酸、花生四烯酸、α-亚油酸和二十二碳六烯酸)相比,油酸对PPARβ(δ)具有最高的配体活性,在脂肪组织和维持骨骼肌功能方面有重要作用。给老年大鼠补充油酸后,PPARα、PPARβ(δ)及PGC-1α的表达量显著增加,n-3多不饱和脂肪酸(n-3 PUFA)还可以改变肌纤维的氧化代谢,使快肌纤维向慢肌纤维转化[42]。与此一致,Watanabe等[4]研究表明油酸可激活PPARβ(δ)的表达,在转录水平可上调肌球蛋白重链(myosin heavy chain,MyHC)Ⅰ和PGC-1α的基因表达,且能够增加线粒体数量和最大呼吸体积并诱导C2C12肌管中慢肌纤维的形成。为进一步探究PPARβ(δ)与补充油酸诱导慢肌纤维形成之间的关系,该研究小组通过小分子干扰RNA(small interfering RNA,siRNA)转染技术在C2C12成肌细胞中进行了PPARβ(δ)敲除,结果表明PPARβ(δ)经油酸激活后作为转录因子,诱导脂肪酸代谢相关基因的表达;然而MyHCⅠ和线粒体因子的表达并不依赖于PPARβ(δ)的机制调节,由此说明油酸作为PPARβ(δ)配体起激活作用,并诱导脂肪酸代谢基因[丙酮酸脱氢酶激酶同工酶4(pyruvate dehydrogenase kinase isozyme 4,PDK4)、肉毒碱棕榈酰基转移酶1B(carnitine palmitoyltransferase 1B,CPT1B)和PPARα]的表达,从而促进机体氧化代谢。以上研究说明不饱和脂肪酸尤其是油酸能有效促进PPARβ(δ)的表达,同时能上调肌肉中氧化型肌纤维的比例,但是不饱和脂肪酸对肌纤维类型转化调控机制还有待于进一步深入研究。
3.1.2 激动剂
近年来已有多位国内外学者研究证实补充外源性PPARβ(δ)激动剂同样能够使骨骼肌纤维由酵解型肌纤维向氧化型肌纤维转化,但是内在转化机制尚不清楚。
Oliver等[43]在2003年研发出一种苯氧乙酸化合物(GW501516),是迄今最为人们关注的选择性PPARβ(δ)受体激动剂,这种化合物对激活PPARβ(δ)具有更强的选择性,可高出其他亚型1 000倍,在纳摩尔每升的浓度水平就可以使PPARβ(δ)激活。Wang等[12]灌胃小鼠PPARβ(δ)激动剂(GW501516)10 d后,小鼠的线粒体生物合成功能和呼吸氧化作用增强,骨骼肌慢肌纤维比例分布有所上调,且肉色得以改善,表明PPARβ(δ)是肌纤维转化过程的关键因子。为进一步证实体内过表达PPARβ(δ)对肌纤维类型的转化作用,姚国佳[44]对小鼠进行灌胃PPARβ(δ)激动剂(GW501516)后发现,小鼠骨骼肌中PPARβ(δ)mRNA和蛋白表达量有了显著提高,且小鼠骨骼肌中MyHCⅠ和MyHCⅡx基因表达量显著上调,同时MyHCⅡb基因表达量显著下调。方海琴等[45]通过建立小鼠外源性补充PPARβ(δ)激动剂(GW501516)并辅以运动的试验模型,证实了补充PPARβ(δ)激动剂可促进小鼠骨骼肌纤维类型从酵解型向氧化型发生转化,其作用机制可能是激动剂的补充激活了PPARβ(δ)受体,而后与辅激活因子PGC-1α协同作用,增强线粒体的生物合成过程,并促使小鼠骨骼肌纤维类型的转化。荆海军[14]将PPARβ(δ)特异性激动剂GW0742添加于成肌细胞分化培养基中发现,在成肌细胞株和小鼠原代成肌细胞分化过程中PPARβ(δ)的过表达能显著上调氧化型肌纤维的分布比例,这进一步证实了方海琴等[45]的研究,为进一步研究PPARβ(δ)通路诱导肌纤维类型转化机制提供了参考。
3.2 运动
运动作为一种刺激方式可以引起机体产生一系列应答反应。肌纤维与运动训练密切相关,氧化型肌纤维比例高能提升运动耐力,另外,运动训练可促进氧化型肌纤维的形成,从而改变骨骼肌纤维类型分布。但是,就目前所有结论而言,运动究竟能否能诱导骨骼肌PPARβ(δ)mRNA和蛋白的表达,从而使肌纤维类型发生转化还没有从根本上得到证明。
1997年,Zipper[46]首次发现大鼠经运动训练后肝脏中的PPARs基因表达量能显著增加100%~200%,这可能是由于运动时交感神经兴奋,肾上腺素、糖皮质激素和瘦素等分泌增加,胰岛素水平下降,从而与相应的受体相结合,激活其相应的信号转导途径,继而作用于相应的靶蛋白所致。Luquet等[13]为探讨运动与肌肉PPARβ(δ)表达之间的可能关系,研究了游泳训练对小鼠跖肌PPARβ(δ)蛋白表达的影响,结果表明,野生型小鼠经过3周的游泳训练后,骨骼肌PPARβ(δ)含量可增加1倍左右,训练6周后PPARβ(δ)蛋白表达量显著高于前3周训练的水平,这些结论支持PPARβ(δ)具有肌肉运动适应性反应的介质作用。Krämer等[26]研究发现,运动能使PPARβ(δ)的表达量增加,同时可增加肌纤维中MyHCⅠ的比例。在转基因小鼠上的试验也证实,PPARβ(δ)过表达确实引起了骨骼肌纤维类型的转化[12]。陈淦[47]研究发现,在4、8周的有氧运动结束后,小鼠骨骼肌PPARβ(δ)mRNA和蛋白表达量均显著上升,且在8周训练后增加量更为显著。这表明有氧运动诱导了小鼠骨骼肌中PPARβ(δ)的高表达,在肌肉的能量代谢调节方面具有一定的作用。也有研究证实PPARβ(δ)蛋白过表达可以提高机体有氧耐力运动能力,且耐力训练可使骨骼肌中PPARβ(δ)含量明显提高,增加骨骼肌对血液及组织中脂肪酸的氧化利用效率,从而增加能量供给[48]。苏丽等[49]对小鼠进行无负重游泳训练时发现,有氧训练诱导PPARβ(δ)mRNA表达量显著增加,增加量最高甚至可达104%。荆海军[14]研究发现,6周耐力训练后小鼠骨骼肌在转录和翻译水平上导PPARβ(δ)表达量显著增加,这表明小鼠骨骼肌中PPARβ(δ)发生过表达。该结论与Luquet等[13]的研究结果是一致的。然而也有研究表明运动确实能诱导骨骼肌PPARβ(δ)mRNA和蛋白表达量的提高,并使骨骼肌中氧化相关酶类的活性升高,运动耐力增强,但最终并没有使得骨骼肌纤维类型完成转化[50],该结论提示PPARβ(δ)可能在骨骼肌对运动训练的适应性反应中是必需的调节因子。这些结果表明PPARβ(δ)在肌肉发育和对环境变化的适应性反应中起着重要作用。
至于运动训练激活骨骼肌内源性PPARβ(δ)特异性表达的机理,Luquet等[13]认为有3种可能:第一,运动可能促使机体组织中PPARβ(δ)配体含量提高并充分促进脂肪酸的分化和燃烧,PPARβ(δ)内源性配体脂肪酸及其代谢物的增多能在一定程度上激活PPARβ(δ);第二,运动诱导了PPARs家族的转录辅激活因子PGC-1α的表达,从而激活PPARβ(δ);第三,运动训练过程中,PPARβ(δ)表达量的增加可能是通过其上游信号分子如钙离子(Ca2+)等相关信号通路激活所导致。
4 PPARβ(δ)表达与肉品质的关系
PPARβ(δ)基因作为肌肉组织中Ⅰ型肌纤维的生理性重要调节因子,能促使快肌纤维形态及功能发生变化,使其转化为慢肌纤维,提高机体氧化代谢能力。深入了解核受体PPARβ(δ)信号通路及其分子调控机制,对于调控肌纤维类型转化至关重要,可为今后改善畜禽肉品质提供有效的途径。肌纤维类型组成及其比例分布与畜禽肉品质紧密相关,肉品质是一个综合性状,通常人们用肉的色泽、嫩度、pH等指标来评价肉品质的优劣[51-52]。不同类型的肌纤维具有不同的肌球蛋白ATP酶活性、糖原含量以及糖酵解潜力(glycolytic potential,GP),这对宰后肌肉的pH、色泽、剪切力、嫩度等肉品质指标都有重要的影响[53]。肌纤维的生物学特性与这些评定指标直接相关,是肉品质形成的基础。相比于其他评定指标,消费者更关注肉的嫩度,多项研究指出肌纤维的类型分布、数量、直径、横截面积与肉的嫩度间存在一定的相关性,肌纤维的直径越小则其肉质越细嫩;此外,肌纤维类型分布的差异造成了其肌内脂肪含量和结缔组织的差异,从而使肉的嫩度有所不同[54]。杨玉莹等[54]报道,牦牛肉Ⅰ型肌纤维比例越高,肉质越细嫩多汁。肌纤维直径与肉的剪切力呈正相关,与肉的嫩度呈负相关[55-56]。与酵解型肌纤维相比,氧化型(Ⅰ型+ⅡA型)肌纤维直径较细,剪切力略小,且有较高的肌红蛋白、血红蛋白和磷脂含量,当肌肉中氧化型(Ⅰ型+ⅡA型)肌纤维占比较高时,则肌肉更细嫩、风味更佳、色泽鲜红[57]。酵解型肌纤维含有较多的糖原且ATP酶的活性较高,通过糖酵解方式产生ATP,尤其是畜禽屠宰后机体主要供能方式转化为无氧糖酵解,因此富含酵解型肌纤维的肌肉更容易疲劳,糖酵解代谢在重复使用下使肌肉酸化,肌肉pH下降速率较快,易导致PSE肉[58]。由此可见,肌肉中氧化型肌纤维占比高的肉品质更好,可通过提高肌肉中氧化型(Ⅰ型+ⅡA型)肌纤维的占比来改善畜禽肉品质。
5 小 结
综上所述,PPARβ(δ)信号通路在对肌纤维类型转化的调控过程中发挥着至关重要的作用,但是相关的试验证据尚少。PPARβ(δ)通过ERRγ、miRNA网络及Sox6调控骨骼肌慢肌纤维形成的具体信号通路中,伴随着多种复杂的作用机制。就目前而言,PPARβ(δ)信号通路对肉品质影响的相关研究还不够深入,对于PPARβ(δ)信号通路与其他信号通路间的内在关联性,诱导肌纤维类型转化过程中PPARs间协同作用、组织特异性和表达的差异,运动、激动剂、其他辅激活因子对PPARβ(δ)影响的差异等都有待进一步深入研究。进一步了解PPARβ(δ)信号通路中的重要调控因子及差异表达的基因,将为今后通过调控肌纤维类型转化进而改善畜禽肉品质提供有效的途径。
猜你喜欢
中国临床医学影像杂志(2022年6期)2022-07-26
中国临床医学影像杂志(2022年5期)2022-07-26
国际放射医学核医学杂志(2021年10期)2021-02-28
天然产物研究与开发(2018年2期)2018-04-04
材料科学与工程学报(2016年4期)2017-01-15
中国运动医学杂志(2016年3期)2016-07-10
中国运动医学杂志(2016年3期)2016-07-10
合成化学(2015年4期)2016-01-17
医学研究杂志(2015年5期)2015-06-10
动物营养学报(2014年5期)2014-03-23
