
肠道中谷氨酸的功能及其吸收转运调控
2021-10-27 04:12陈明霞刘贤旭梁常杰
畜禽业 2021年10期
陈明霞,刘贤旭,梁常杰
(广东科贸职业学院动物科技学院,广东 广州 510430)
0 引言
研究表明,谷氨酸转运载体SLC1A1(EAAT3)功能缺失性突变是二羧基氨基酸尿症的原因[1]。谷氨酸可以减缓肠道诱发性细胞旁路渗透性过高,有利于保护肠道的完整性[2],显示出谷氨酸及其转运载体对人体和动物健康的重要性。
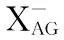
本文围绕肠道中谷氨酸发挥上皮细胞能源物质、生物活性分子前体物质和肠道神经系统神经递质的功能,以及肠上皮细胞主要的谷氨酸转运载体及其表达和转运活性调控进行综述,为谷氨酸营养在动物,尤其是幼龄动物肠道发育和健康方面的应用提供参考。
1 肠道中谷氨酸的功能
1.1 能源物质
肝脏是氨基酸分解代谢和氧化的主要场所,而对小肠而言,其不仅是营养物质消化吸收的主要组织,在日粮氨基酸的代谢中也扮演着重要的角色。肠道是谷氨酰胺、谷氨酸和天冬氨酸等非必需氨基酸分解代谢的主要场所之一[5]。
氨基酸是小肠黏膜的主要能源物质,日粮中大部分谷氨酰胺、几乎所有的谷氨酸和天冬氨酸没有进入门脉循环流到小肠以外的组织,大部分谷氨酸在跨膜转运到肠上皮细胞的过程中就被代谢,使得首过代谢(first-pass)成为研究肠道中谷氨酸代谢不可忽视的问题。肠道中谷氨酸代谢主要在肠上皮细胞中进行,氧化代谢是其主要代谢形式。肠道中谷氨酸代谢非常迅速,98%以上的谷氨酸转化为α-酮戊二酸,然后进入三羧酸循环途径并产生ATP[6]。肠上皮细胞中ATP的生产和利用非常活跃,上皮细胞每几天就更新一次,对能量的需求比较大,尽管胃肠道只占体重的约5%,但氧消耗却占到整个机体20%左右。在星形细胞中,谷氨酸被吸收进来后在谷氨酰胺合成酶的作用下酰胺化为谷氨酰胺。谷氨酸和谷氨酰胺对肠上皮细胞氧化供能有促进作用。然而,小肠中谷氨酰胺和谷氨酸的代谢是高度区分的。当谷氨酸和谷氨酰胺同时出现在肠上皮细胞时,谷氨酸可以抑制谷氨酰胺的利用和氧化[7]。
1.2 前体物质
肠黏膜中的谷氨酸不但可以用于合成蛋白质,同时可以作为生成天冬氨酸、丙氨酸、脯氨酸、鸟氨酸和瓜氨酸的前体物质。在肠上皮细胞细胞质和线粒体中,谷氨酸在天冬氨酸氨基转移酶、丙氨酸氨基转移酶、支链氨基酸氨基转移酶和谷氨酸脱氢酶的作用下进行转氨代谢[23]。大鼠肠道原位灌注以及仔猪和人的在体研究表明,大部分谷氨酰胺(55%~70%)、谷氨酸(52%~64%)和天冬氨酸(52%)氧化成为CO2,剩余的氨基酸将转化为乳酸、丙氨酸、脯氨酸、瓜氨酸和精氨酸,然后进入门静脉循环。
谷氨酸与半胱氨酸、甘氨酸是肠上皮细胞质合成谷胱甘肽的前体物质,具有抗氧化作用的谷胱甘肽的合成主要受半胱氨酸和谷氨酸的限制。仔猪肠黏膜谷胱甘肽主要由谷氨酸直接代谢衍生而来,仔猪哺乳后小肠中的谷氨酸脱氢酶活性增加了将近3倍[8]。因此,谷氨酸对提高肠道的抗氧化作用也起到一定的作用[9]。
肠上皮细胞中的谷氨酸还可以参加其他一些代谢通路,例如N-乙酰谷氨酸的合成。N-乙酰谷氨酸由乙酰辅酶A和谷氨酸在N-乙酰谷氨酸合成酶的作用下合成。N-乙酰谷氨酸是氨基甲酰合成酶的别构激动剂,因此,谷氨酸衍生N-乙酰谷氨酸代谢过程可能对肠上皮细胞生成瓜氨酸发挥一定的作用。
1.3 神经递质
谷氨酸是主要的兴奋性神经递质,在脑中含量非常丰富,但大部分都是在胞内,细胞膜两侧谷氨酸浓度有几千倍的差异。谷氨酸的周转周期很短,胞内的谷氨酸持续不断地释放到胞外,胞外的谷氨酸也持续不断地转运到胞内。谷氨酸通过谷氨酸受体发挥信号传导作用[10],谷氨酸受体位于能够表达其的细胞表面,因此,细胞周围的细胞外液决定受体兴奋性的程度,胞外谷氨酸浓度保持在较低水平非常关键。
消化系统中有其自身的神经系统,通常称之为肠道神经系统,肠道神经系统非常复杂,含有各种功能不同的神经元,它在一定程度上受中枢神经的指挥,同时可以反馈调节中枢神经,在不受外部神经信号影响的条件下也可以完成一系列反射和整合活动。谷氨酸可能在肠道神经系统中发挥神经递质的作用,EAAT3在肠道神经元中广泛表达[11],是调节肠道神经系统谷氨酸浓度的关键功能基因。越来越多的证据表明,谷氨酸在肠道神经系统中具有信号分子的作用,与鲜味[12]和胃肠道营养信号[13]共同调节神经内分泌反射。
2 肠道中谷氨酸的吸收转运
2.1 肠道中谷氨酸转运载体
上皮细胞中谷氨酸代谢的第一步是谷氨酸转运,肠腔中的谷氨酸跨过顶膜转运到肠上皮细胞中主要是通过谷氨酸转运载体来完成。
EAAT3主要是在小肠中表达,胃和大肠中的表达量都比较低,EAAT1和EAAT2在大多数胃细胞的细胞器中都有表达,但小肠中的表达相对较低。仔猪、大鼠和反刍动物上的研究表明,小肠上皮细胞中检测不到EAAT4和EAAT1蛋白的表达,仔猪和反刍动物中可检测到EAAT2的表达。EAAT3转录和翻译水平的表达量随着隐窝-绒毛轴上皮细胞的分化程度而增加[16-19]。肠上皮细胞中,EAAT2和EAAT3分别在基底膜和顶膜发挥作用,EAAT3是小肠上皮细胞底膜主要的谷氨酸转运载体。
2.2 肠道中谷氨酸转运调控
肠道中谷氨酸转运效率调控主要包括2种途径:①可通过改变谷氨酸转运载体的表达量进行调控。②也可以通过调节谷氨酸转运载体转运活性来实现。
Mott等的研究表明,慢速生长型鸡肠道EAAT3 mRNA表达高于快速生长型鸡,这可能是因为EAAT3高表达有利于提高谷氨酸/天冬氨酸的吸收,从而提供能量促进其他营养物质转运载体的基因表达和肠道的发育[20]。同样地,对3~14日龄雏鸡的研究表明,EAAT3 mRNA表达丰度在不同品系中也存在显著性差异[21],EAAT3 mRNA表达受品系、日粮蛋白组成、肠段和日龄的影响[22]。黎相广等通过对温氏土鸡和隐性白洛克鸡胚小肠的研究发现,温氏土鸡鸡胚小肠EAAT2和EAAT3 mRNA丰度高于隐性白洛克,其表达量受发育水平的影响,存在“品种×胚龄”互作效应,而且EAAT3 mRNA丰度高于EAAT2 mRNA[23]。
雷帕霉素靶蛋白(target of rapamycin,TOR)是调节蛋白质合成的重要信号通路,谷氨酸能上调m TOR 通路的表达。Makky等研究表明,脊椎动物肠道上皮细胞的发育受TOR信号通路的调控[24],TOR作为PI3K/Akt信号通路的下游基因可调节EAAT2的表达[25],SGK1(serum-and glucocorticoid-Inducible kinase 1)、PDK1(phosphoinositide-dependent kinase 1)和PDGF(platelet-derived growth factor)通过活化Akt/PI3K信号通路上调EAAT3的表达,GTRAP3-18(glutamate transporter associated protein 3-18)或者DOR(δ-opioid receptor)的交互作用可以下调EAAT3的表达[26],肠道中EAAT2和EAAT3基因表达差异是否受TOR信号通路的调控有待进一步研究。
在谷氨酸药理学作用和不同类型谷氨酸转运载体功能研究方面,常常用到各种类型的谷氨酸转运载体(EAATs)抑制剂,这些抑制剂大多数是谷氨酸或天冬氨酸的类似物,可以特异性抑制谷氨酸的吸收。
大部分第一代EAATs抑制剂是一些竞争性的底物,可竞争性地抑制谷氨酸的吸收,同时可以诱导兴奋性氨基酸(谷氨酸和天冬氨酸)从细胞质中转运到细胞外[27]。阻断剂(非转运性抑制剂)含有可以非损伤性结合但产生移位的特异性取代基,可以有效地阻断EAATs对谷氨酸的转运,这些阻断剂的发展有利于EAATs内在特性和生理功能的研究[28],L-trans-pyrrolidine-2,4-dicarboxylate(t-2,4-PDC)和DL-threo-b-benzyloxyaspartate(DL-TBOA)分别是竞争性底物和阻断剂的代表[29]。
3 结语
谷氨酸作为非必需氨基酸,在肠道发育和肠上皮细胞更新能量提供、生物活性物质生成以及肠道神经系统信号传导方面发挥着重要的作用。肠道中谷氨酸的吸收转运主要依赖于EAAT3,其表达水平和转运活性的调控影响谷氨酸转运吸收以及功能的发挥。谷氨酸对动物,尤其是幼龄动物肠道发育和健康具有重要作用,但是目前这方面的研究较少,谷氨酸调控动物肠道发育和维持肠道健康的生产应用有待进一步研究。
猜你喜欢
中国动物保健(2022年9期)2022-09-25
食品与生物技术学报(2021年4期)2021-01-17
中成药(2017年6期)2017-06-13
中国洗涤用品工业(2017年2期)2017-04-16
中国调味品(2017年2期)2017-03-20
化工设计通讯(2017年10期)2017-03-02
中国比较医学杂志(2017年5期)2017-01-17
中外医疗(2015年11期)2016-01-04
听力学及言语疾病杂志(2015年5期)2015-12-24
医学研究杂志(2015年12期)2015-06-10
