
球孢白僵菌对小亚璃眼蜱的毒力测定及其孢外水解酶活性分析
2021-12-11 05:18任巧云漆晶晶仲富萍
中国动物检疫 2021年12期
孙 明,任巧云,漆晶晶,聂 英,马 超,赵 鑫,潘 丽,仲富萍
(1.甘肃省民勤县畜牧兽医工作站,甘肃武威 733000;2.中国农业科学院兰州兽医研究所,家畜疫病病原生物学国家重点实验室,甘肃省动物寄生虫病重点实验室,甘肃兰州 730046;3.甘肃省武威市畜牧兽医总站,甘肃武威 733000)
小亚璃 眼 蜱(Hyalomma anatolicum anatolicum)是我国目前仅报道分布于新疆维吾尔自治区半荒漠地区和草原地区的人畜共患病的重要传播媒介,特别在地方性血液原虫病方面危害更为严重[1-2]。据吴永红等[3]在南疆部分散养户牛场收集的蜱种统计,小亚璃眼蜱占收集蜱总数的20%~30%,且在当地主要传播牛环形泰勒虫病,是制约当地牧区畜牧业发展,危害人畜健康的重要因素。随着人们对公共卫生、食品安全等问题的关注,化学药物防治蜱类所引起的环境污染、蜱虫耐药性提升、动物产品中药物残留等问题日益突出,因而实现蜱的无害化长效控制是当前该研究领域的热点。
利用蜱的自然生物天敌研制生防制剂,这一防控模式在植物病虫害上已经发挥了显著作用。在蜱生物防治方面,为建立和改进蜱的防治模式,国内外研究学者对其病原微生物,特别是栖息在土壤环境中的昆虫真菌类病原微生物进行了广泛采集、筛选和研究[4-5],取得了可喜成效,如中国农业科学院兰州兽医研究所研制的白僵菌与苦皮藤素的杀蜱新制剂、绿僵菌与溴氰菊酯的杀蜱新制剂正在推广应用。
毒力测定是评价高毒力菌株应用潜力的基础。为探索一种比较快捷的方法替代目前繁琐的生物测定,人们一直尝试利用生化方法(酶活性的测定)来筛选强毒菌株[6]。St leger 等[7]在昆虫体壁离体培养时,首次发现了蛋白酶的活性,并指出其与入侵昆虫表皮存在联系。在孢子萌发、入侵和完成侵染过程中,蛋白酶和几丁质酶被认为是与毒力相关的主要酶类,尤其孢外蛋白酶在侵染宿主过程中降解宿主表皮成分,并在宿主血腔中使血淋巴酚原氧化酶超量表达,导致宿主因中毒而死亡[8]。冯明光[6]研究也认为,孢外蛋白酶对血黑蝗的毒力相关性达到0.67,就可以作为菌株初期筛选的指标。李建庆等[9-10]用过滤除去小分子毒素的球孢白僵菌全蛋白提取液进行了生物测定,4 d 内可将6 头试虫全部毒死,首次提取出了球孢白僵菌大分子毒蛋白;通过进一步试验从虫生真菌中提取对松墨天牛有致死作用的代谢物进行研究,可以确定是毒蛋白导致了昆虫死亡,为研究高毒力蛋白和菌株筛选工作奠定了基础。昆虫病原真菌对寄主的入侵是机械压力和酶共同作用的结果,对蜱病原真菌的毒力测定却一直停留在以蜱死亡率为基础的生物测定上,与蜱病原真菌毒力相关的水解酶及其酶活性研究极少。因此,为填补酶类及其活性研究作为蜱致病性真菌筛选的补充指标,进行了以下研究,以期为有效利用球孢白僵菌对蜱虫的生物防控奠定基础。
1 材料与方法
1.1 试验材料
1.1.1 菌株和蜱种 供试菌株,为本实验室分离保藏的球孢白僵菌菌株(B.bAT03、B.bAT11、B.bAT14、B.bAT17 和B.bAT25);供试蜱虫为小亚璃眼蜱,采自新疆维吾尔自治区莎车县牛和羊的体表,经实验室人工繁育后,选择饱血雌蜱用于试验。
1.1.2 培养基 基础培养基为PDA 培养基,明胶琼脂培养基(GA)和壳聚糖培养基(CA)配制方法参考文献[11],蛋白酶诱导培养基和几丁质酶诱导培养基配制方法参考文献[12],几丁质酶完全培养基配制方法参考文献[13]。
1.2 方法
1.2.1 菌株接种、培养和观察 配制各菌株孢子悬浮液,含孢量约为1×107个/mL,用移液器各吸取2 μL 分别点接于GA 和CA 平板上,每个培养皿点接3~4 个菌落,使菌落间相隔呈正三角形或正方形,接种后置于26 ℃恒温保湿培养箱中培养,GA 和CA 培养基平板各做3 个重复。各菌株在GA 平板上培养6 d 后,将适量15%升汞溶液滴加在GA 平板上使其完全覆盖平板,待透明圈显露稳定时倒去升汞溶液,然后用十字交叉法测量各菌落直径和透明圈直径并记录。CA 平板用5% NaOH溶液处理,透明圈的测量方法同上。
1.2.2 胞外蛋白酶诱导培养和活性测定 将孢子悬液(107个/mL)按1% 的量接种到蛋白酶诱导培养基中,26 ℃、180 r/min 培养96 h,用灭菌双层纱布过滤,所得滤液即为酶液。采用Folin-酚法测定蛋白酶活性,以每分钟催化分解生成1 μg 酪蛋白的酶量,定义为1 个蛋白酶活力单位(IU/mL),重复测定3 次[13]。
1.2.3 几丁质酶诱导培养和活性测定 将孢子悬液(107个/mL)按1%的量接种到几丁质酶完全培养基中,26 ℃、180 r/min 培养96 h,用灭菌双层纱布过滤,无菌水洗涤菌体;相同条件下接入几丁质酶诱导培养基振荡培养24 h;用灭菌双层纱布过滤菌体,所得滤液即为酶液。采用DNS 法测定几丁质酶活性,以每分钟水解胶体几丁质产生1 μg还原糖所需的酶量,定义为1 个蛋白酶活力单位(IU/mL),重复测定3 次[14]。
1.2.4 不同菌株对小亚璃眼蜱毒力测定 将球孢白僵菌菌株分别接种于PDA 培养基上培养14 d,将所产孢子粉用含0.05%吐温-80 的无菌水配制成108个/mL 的孢子悬液;将小亚璃眼蜱饱血雌蜱随机分组,每组15 只;采用浸渍法,每组供试蜱分别单个浸渍于孢子悬液5 min 后单独置入一次性无菌培养皿中,在26 ℃、相对湿度为80%的条件下观察小亚璃眼蜱死亡情况,以含0.05%吐温-80 的无菌水作对照,做好记录,并对死亡虫体进行保湿培养(相对湿度大于90%),观察其是否变为僵虫。
1.3 数据统计分析
对所有测定数据利用DPS 数据分析软件进行统计和处理。
2 结果
2.1 不同菌株在GA 平板上透明圈和菌落直径测定
不同菌株在GA 培养基上培养6 d 后,用适量15%升汞溶液处理GA 平板,平板上均出现明显的透明圈。由表1 直径比可见,菌株蛋白酶分泌量差异明显,球孢白僵菌B.bAT17 和B.bAT11 菌株差异不显著,但其直径比均大于其他菌株,球孢白僵菌B.bAT17 菌株较B.bAT03、B.bAT14 和B.bAT25 菌株差异显著。
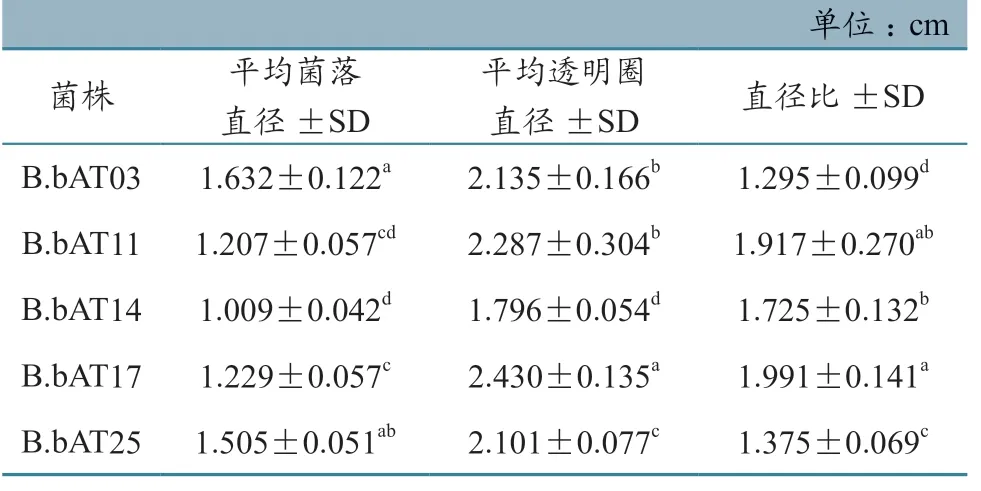
表1 不同菌株在GA 平板上培养6 d 后的透明圈和菌落直径
2.2 不同菌株在CA 平板上透明圈和菌落直径测定
不同菌株在CA 培养基上培养6 d 后,用适量5% NaOH 溶液处理CA 平板,平板上均出现明显的透明圈。由表2 可见,在几丁质酶活性上,球孢白僵菌B.bAT03、B.bAT14、B.bAT25 菌株间直径比差异不明显,B.bAT17 菌株产酶量低于B.bAT11且差异显著。
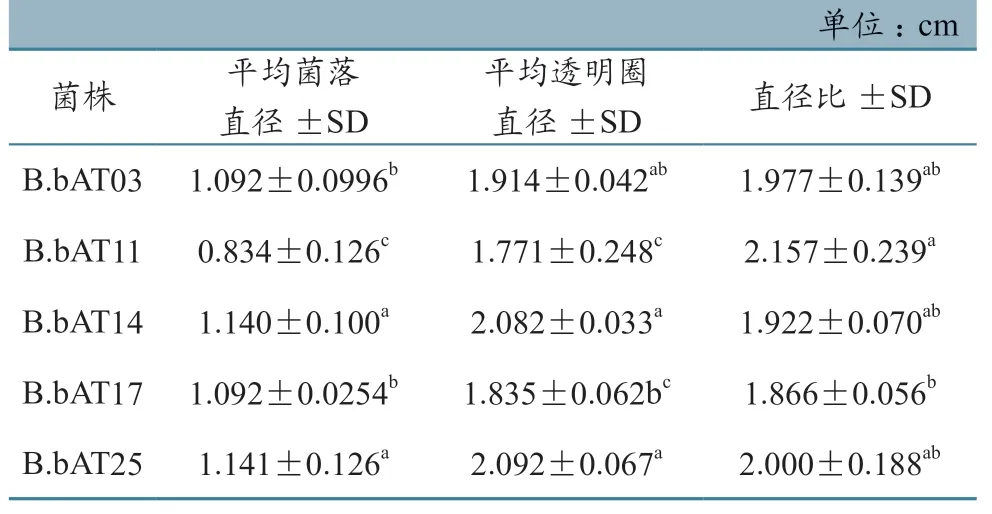
表2 不同菌株在CA 平板上培养6d 后的透明圈和菌落直径
2.3 不同菌株对蜱毒力测定
如图1 所示,在孢子密度为108个/mL 时,各菌株对小亚璃眼蜱饱血雌蜱均有不同程度的侵染性,菌株对蜱的累计致死率为40%~100%,其中球孢白僵菌B.bAT17 菌株显示了较高的毒力,致死率达到100%。试验统计中,被生防真菌侵染的饱血雌蜱初期行动呆滞,呈萎缩状态,体壁出现黑色小点,随着病情发展,黑点逐渐扩张。在感染后期,蜱开始出现死亡,全身变成深黑色,部分蜱表皮破裂,血液外渗,体表长出白色絮状菌丝,随后菌丝布满全身,产生分生孢子。
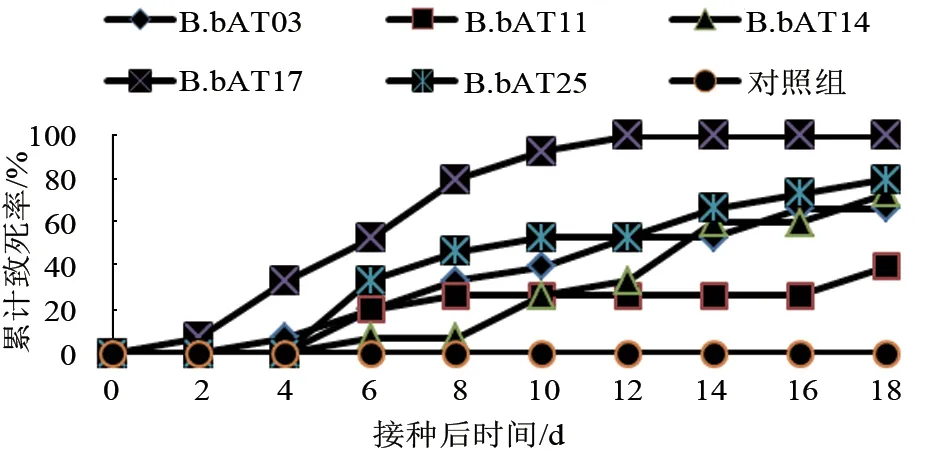
图1 不同菌株侵染小亚璃眼蜱的累计致死率变化趋势
2.4 不同菌株孢外水解酶活性与其对小亚璃眼蜱侵染毒力的相关性分析
不同菌株孢外蛋白酶活性和几丁质酶活性见表3。采用DPS 数据处理软件对菌株孢外水解酶活性与其毒力之间进行相关性分析和构建毒力方程。校正死亡率(Y)与蛋白酶活性(X1)间毒力方程为Y=9.202 5X1+0.852 5,相关系数R为0.671 9(P=0.225),球孢白僵菌5 个菌株的蛋白酶活性与其对小亚璃眼蜱饱血雌蜱侵染毒力无相关性(P>0.05)。校正死亡率(Y)与几丁质酶活性(X2)间毒力方程为Y=1.676 1-1.165 6X2,相关系数R为0.935 1(P=0.020),5 个球孢白僵菌菌株的几丁质酶活性与其对小亚璃眼蜱饱血雌蜱侵染毒力呈负相关关系,且差异显著(P<0.05)。

表3 不同菌株孢外水解酶的活性测定 单位:IU/mL
3 讨论
球孢白僵菌作为一种广谱性的微生物型杀虫剂,在农、林害虫的生防领域中受到极大重视,科研工作者对其杀虫机理和制剂的生产进行了大量研究和创新,通过基因工程的超量表达、基因敲除等方法,对其毒力因子进行验证,菌株“孢外水解酶”假说被科研人员所公认[15]。对于节肢动物蜱,国外学者研究已表明,真菌侵染的早期蛋白酶——类枯草杆菌蛋白酶(Pr1)由于具有氨基酸疏水性基团的结构特征,对蜱表皮蛋白延伸的疏水肽键有比较高的活性,在真菌孢子入侵蜱的早期对蜱表皮具有很强的降解能力,被认为是与真菌致病性有关的主要毒力因子之一[16]。Fang 等[17]构建了编码孢外蛋白酶和几丁质酶的融合基因(CDEP1:Bbchit1),菌株融合基因酶的表达量符合蛋白酶和几丁质酶的交互协同作用且增强了菌株毒力。
在蜱生物防治研究领域,国外学者通过扫描显微镜研究发现,在真菌感染蜱时,其分泌的孢外水解酶(蛋白酶、几丁质酶和酯酶等)在孢子芽管穿透体壁,降解蜱表皮角质层方面起着重要作用[18-19]。在菌株孢外蛋白酶和几丁质酶平板透明圈法测定中,各菌株表现出较高的孢外水解酶分泌量(表1~2)。孢外水解酶的产量多少与活性大小关系到菌株侵染力,人们甚至试图将其作为菌株毒力的参考指标。胡景江等[20]用GA 平板法测量的各球孢白僵菌菌株以及各世代胞外蛋白酶产酶水平与其对马尾松毛虫的毒力之间存在着明显线性关系,可以用定量的产酶水平来指示球孢白僵菌菌株对目标昆虫的毒力大小。本次试验也证实高毒力菌株B.bAT17 在GA 平板固体培养基上产生较大的蛋白酶水解圈,且菌株直径比与B.bAT03、B.bAT14、B.bAT25 相比差异显著。
在孢外水解酶活性与菌株毒力的相关性研究中,用DPS 分析软件处理数据表明,蛋白酶活性与菌株毒力间相关系数R=0.671 9(P>0.05),而几丁质酶活性与菌株毒力间相关系数R=0.935 1(P<0.05),具有相关性(表3),这可能是因为几丁质酶属于诱导酶,待早期蛋白酶将宿主表皮蛋白降解后,几丁质酶才开始大量分泌。不过,也有资料显示菌株产酶能力高低与其致病力之间不一定具有必然联系。冯明光[6]曾报道孢外蛋白酶活性作为毒力的参考指标需谨慎使用,St leger 报道球孢白僵菌无毒菌株比毒性菌株产生更多蛋白酶[7-8]。
生物防治研究在国内蜱生物防控中刚刚起步,对其酶制剂的研究极少,对酶的种类研究也比较单调,尚未将其水解酶的协同作用完全考虑。在蜱生防真菌的菌株筛选工作中,平板透明圈法是有效且快速便捷的初步筛选高毒力菌株的方法,克服了评价菌株繁琐的毒力测定步骤。同时,为进一步研究菌株水解酶活性对蜱虫的毒力,也可考虑通过基因工程等分子生物学方法,对病原真菌降解宿主体壁的几丁质酶和蛋白质酶进行分子改良,以期获得产酶活性高、性质稳定、应用潜力大、易于培养的蜱生防高毒力菌株。本研究结果为蜱生防高毒力菌株的筛选以及酶活性的初步分析提供了理论依据。
猜你喜欢
食品工业科技(2021年22期)2021-11-14
南方农业(2021年8期)2021-06-03
生物技术通报(2021年4期)2021-05-14
湖北林业科技(2019年2期)2019-05-05
中国酿造(2017年8期)2017-09-03
中成药(2017年5期)2017-06-13
上海农业学报(2016年2期)2016-10-27
华南农业大学学报(2016年4期)2016-07-18
广西林业科学(2016年4期)2016-03-16
浙江柑橘(2013年2期)2013-01-22
