
草地退化影响高原鼠兔血清总IgG水平、肠道寄生物感染及肝脏和肾脏指数
2022-09-21 14:37吴学琴陈慧青张璐尚国珍吴雁边疆晖
兽类学报 2022年5期
吴学琴 陈慧青 张璐 尚国珍 吴雁 边疆晖
(1 中国科学院西北高原生物研究所,中国科学院高原生物适应与进化重点实验室,西宁810001)(2 中国科学院大学,北京100049)(3 青海省动物生态基因组学重点实验室,西宁810001)(4 杭州师范大学生命科学学院,杭州 310012)
高原鼠兔(Ochotona curzoniae) 隶属于兔形目(Lagomorpha) 鼠兔科(Ochotonidae) 鼠兔属,广泛栖息于海拔3 200~ 5 200 m 植被低矮的高寒草甸地区,是青藏高原特有的非冬眠小型植食性哺乳动物,也是高寒草甸生态系统的关键种(Smith and Foggin,1999;Wilson and Smith,2015;Smithet al.,2019)。近几十年来,由于过度放牧引起草地退化,导致高原鼠兔种群数量暴发式增长,最高时可达2 400 只/公顷(来德珍等,2006)。有关草地退化与高原鼠兔种群数量关系的研究表明,放牧降低了植被的高度和盖度,为高原鼠兔提供了开阔的栖息环境,有利于其躲避天敌,降低被捕食的风险,进而导致种群暴发(施银柱,1983;边疆晖等,1994),但引起高原鼠兔种群暴发的生理机制,目前尚不清楚。
免疫力是影响植食性小哺乳动物种群波动的重要调节因子(Mihoket al.,1985;Sinclair and Lochmiller,2000;张志强和王德华,2005),动物免疫力降低可增加病原体的感染(Nordlinget al.,1998;Holt and Roy,2007;Cornellet al.,2008;Hawley and Altizer,2011;Duet al.,2016),继而降低种群数量(Yanget al.,2018;Shanget al.,2019);反之,个体免疫力增强,种群死亡率降低,越冬存活率增加(Książeket al.,2014),则促进种群增长(Lochmiller,1996;Nelson,2004)。在器官水平上,肝脏和肾脏对氧化降解和排除植物次生化合物具有重要作用(Caldwell,1982;Lindroth and Batzli,1983;Foley,1992;李俊年等,2003a,2003b;何岚等,2010),对小哺乳动物的生存至关重要(张志强等,2007)。在青藏高原地区,草地不同退化程度为高原鼠兔提供了不同的植物资源,而不同的食物资源对动物免疫力及生长发育有不同的效应。例如高蛋白质食物可增强个体免疫力(Lochmilleret al.,1993;Davis and Lochmiller,1995);高次生化合物含量食物抑制个体的生长发育(段传人等,2003;冶娜娜,2015)。
据此推测,高原鼠兔的种群数量随草地退化而增加,可能是因为退化草地增强了高原鼠兔个体的免疫力,同时缓解了植物次生化合物对肝脏和肾脏的损伤。为了验证该假设,本研究在青海海北高寒草地生态系统国家野外科学观测研究站地区,分别调查了轻度、中度、重度3类退化草地中高原鼠兔洞口数量并测定了高原鼠兔血清总IgG含量、肠道寄生物感染状况以及肝脏和肾脏指数。通过分析不同退化草地对高原鼠兔天然免疫力能力及肝脏和肾脏的效应,可为揭示高原鼠兔种群暴发的生理机制提供部分理论依据。
1 研究方法
1.1 实验地点与设计
本实验于2021年7月在青海海北高寒草地生态系统国家野外科学观测研究站进行。根据实地访问调查的放牧强度(放牧家畜以藏系绵羊和牦牛为主),确定并选择了轻度(lightly degraded grassland,LD;每公顷4.14 个羊单位)、中度(moderately degraded grassland,MD;每公顷8.77 个羊单位)和重度退化草地(heavily degraded grassland,HD;每公顷14.44 个羊单位)各5 块样地,共计15 块样地,每块样地约4 hm2,不同程度退化草地各样地间相距300 m以上,以避免样地间的假重复。
首先,采用样线法计数每块样地中半径为50 m的圆面积样方(3个重复)内高原鼠兔洞口数量,以此估算种群密度。随后采用绳套法活捕各样地中的高原鼠兔,采用毛细吸管经眼眶静脉窦采血100~200 μL 于0.5 mL 的离心管中,并将离心管保存在有冰袋的采样盒中静置30 min,带回实验室后,4 000 r/min,4℃离心15 min,取上清液,-20℃冰箱保存,留测血清总免疫球蛋白G (Immunoglobin G,IgG) 含量。采血后立即将高原鼠兔脱臼处死,置于冰盒,带回实验室称其体重,并解剖成体高原鼠兔(雄性体重大于130 g,雌性体重大于120 g;周立等,1987),剥离高原鼠兔的肝脏和肾脏,用干净滤纸吸去器官上的血液后称重(精确到0.001 g),随后收集结肠内容物,用于测定肠道寄生物感染状况。
1.2 血清总IgG含量测定
采用酶联免疫法(Enzyme linked immune sorbentassay,ELISA) 测定成体高原鼠兔血清总IgG含量。酶联免疫试剂盒购自上海酶联生物科技有限公司(货号:ml027974),具体测定方法如下:取96 孔酶标包被板,分别设置标准品孔、空白孔和待测样品孔。标准品孔各加不同浓度的标准品50 μL,待测样品孔中先加样品稀释液45 μL,再加高原鼠兔血清5 μL,空白孔加样品稀释液50 μL。接着每孔加入酶标试剂100 μL(空白孔除外) 后用封板膜封板置 于37℃温育60 min。然后弃去液体,甩干,使用1 × 洗涤液洗涤5 次,每次静置30 s 甩干。随后每孔分别先后加入显色剂A 和B 各50 μL,37℃避光显色15 min。最后,每孔加入终止液50 μL,于450 nm 波长依序测量各孔的吸光值。依据测定的吸光值绘制标准曲线并计算高原鼠兔血清总IgG 含量。
1.3 寄生物感染的测定
肠道寄生物的分类计数采用改良的麦克马斯特法(Wetzel,1951)。用电子天平称取0.200 g (精确到0.001 g)粪便样品,置于研钵,加入2 mL 饱和NaCl 溶液研磨,依次通过40 目铜网筛和100 目尼龙网筛过滤后,将滤液用饱和NaCl 溶液定容至5 mL。采用饱和盐水漂浮法,10 倍物镜下统计计数板网格内各类寄生物卵囊数(聂绪恒等,2014),并计算其感染率和感染强度(Bushet al.,1997)。感染率=样地取样个体感染数/样地取样的总个体数,感染强度为每克粪便中的卵囊数(Oocysts per gram,OPG),OPG=(寄生物计数×5)/(0.15×2×粪便重)。
1.4 统计分析
采用广义线性混合模型(Generalized Linear Mixed Models,GLMMs),以不同退化草地类型和性别作为固定因子,样地作为随机变量,检验高原鼠兔洞口数量、肠道寄生物感染率和感染强度、血清总IgG 含量、肝脏指数和肾脏指数在各因子及各因子间交互效应的显著性。由于肝脏和肾脏的绝对重量与体重具有显著相关性(相关系数分别为肝脏:r=0.730,P<0.01;肾脏:r=0.466,P <0.001),所以将肝脏和肾脏的绝对重量换算为脏器指数(脏器湿重/体重×100%)以排除体重的效应,并对数据经Kolmogorov-Smirnov 和Levene 进行正态性和方差齐性检验,发现肝脏和肾脏指数不符合正态分布,故分析前进行反正弦转换。由于肠道寄生物感染强度的范围在0~253 111,所以分析前对其进行ln(OPG+1)转换;采用Bonferroni进行多重比较,数据表示为平均值±标准误(mean±SE),显著水平为P<0.05。所有统计均在SPSS 21.0 软件中进行。
2 结果
2.1 不同退化草地高原鼠兔洞口数量
不同退化草地中高原鼠兔的洞口数量差异显著(F2,12=17.71,P<0.001) (图1),重度退化草地显著高于中度和轻度退化草地(P<0.05),且中度退化草地显著高于轻度退化草地 (P<0.05)(图1)。

图1 不同退化草地中高原鼠兔的洞口数量(平均值±标准误).图中不同小写字母表示差异显著(P <0.05)Fig.1 The number of burrow entrances of plateau pika in lightly(LD),moderaterly(MD)and heavily degraded grassland(HD)(mean±SE).The lowercase indicates significant difference(P <0.05)
2.2 不同退化草地高原鼠兔血清总IgG含量
草地退化显著影响高原鼠兔血清总IgG 含量(F2,138=3.38,P=0.037)(图2)。重度退化草地中高原鼠兔血清的总IgG 含量显著高于轻度退化草地(P<0.05),而中度与轻度和重度退化草地之间均无显著差异(P>0.05) (图2)。性别(F1,138=0.00,P=0.991) 及性别与草地退化的交互作用(F2,138=1.26,P=0.287) 对高原鼠兔的血清总IgG 水平无显著影响。
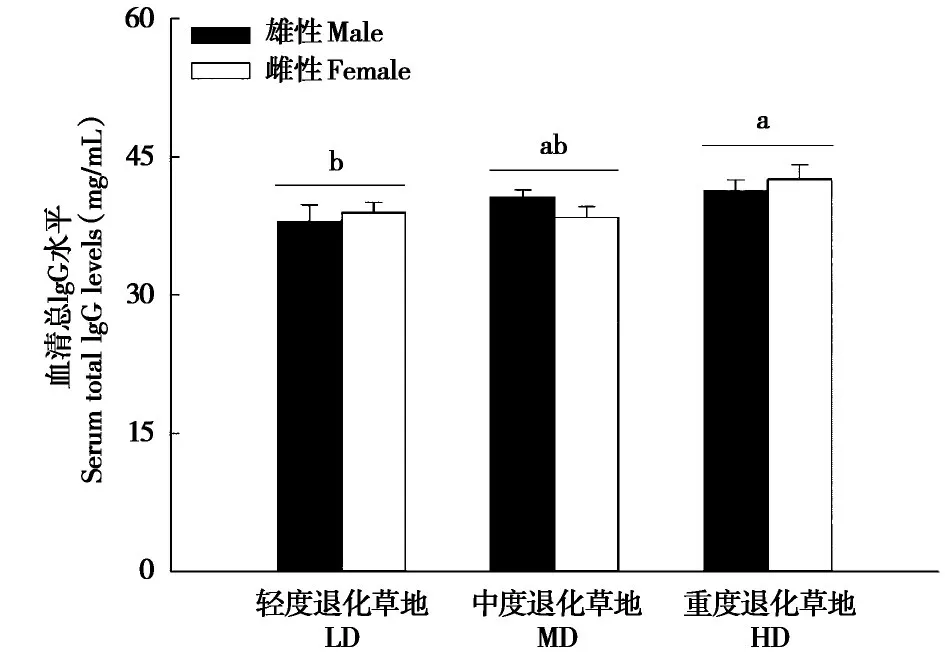
图2 不同退化草地中高原鼠兔血清总IgG 水平(平均值± 标准误).轻度、中度和重度退化草地的雄性样本量分别为17、25 和23,雌性样本量分别为26、33 和20.图中不同小写字母表示差异显著(P <0.05)Fig.2 Serum total IgG levels of plateau pika in LD,MD and HD(mean±SE). n=17,25 and 23 for males and n=26,33 and 20 for females in LD,MD and HD,respectively.The lowercase indicates significant difference(P <0.05).For abbreviations,see Fig.1
2.3 不同退化草地高原鼠兔肠道寄生物感染状况
高原鼠兔在3类退化类型草地中的球虫感染率均为100%,草地退化显著影响高原鼠兔对绦虫(F1,8=7.34,P=0.027) 和线虫(F2,12=6.22,P=0.014)的感染率(图3A、B、C)。在中度退化草地中,高原鼠兔未感染绦虫,且对线虫的感染率显著低于轻度退化草地(P<0.05);在重度退化草地中,对绦虫和线虫的感染率均显著低于轻度退化草地(P<0.05)(图3B、C)。性别显著影响高原鼠兔对绦虫的感染率(F1,8=5.41,P=0.049),雄性的感染率显著低于雌性(P<0.05),但是对线虫的感染率无显著影响(F1,12=4.06,P=0.067)。草地退化与性别的交互作用显著影响高原鼠兔对绦虫(F1,8=9.11,P=0.017)和线虫(F2,12=5.48,P=0.020)的感染率,雌性高原鼠兔在重度退化草地中对二者的感染率显著低于轻度退化草地(P<0.05);而雄性高原鼠兔在不同退化草地中对二者的感染率无显著变化(P>0.05)。
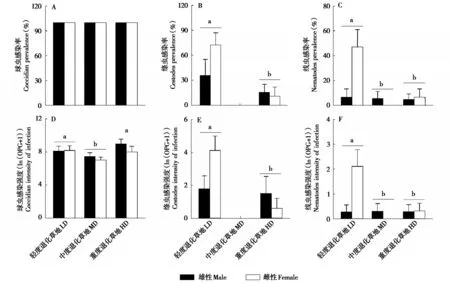
图3 不同退化草地中高原鼠兔对球虫、绦虫和线虫的感染率及感染强度(平均值±标准误).轻度、中度和重度退化草地的雄性样本量分别为12、11 和11,雌性样本量分别为13、19 和10.图中不同小写字母表示差异显著(P <0.05).由于所检测的高原鼠兔均感染了球虫,所以未对球虫的感染率进行统计分析.中度退化草地中高原鼠兔未感染绦虫,所以统计分析时删除了该组Fig.3 Prevalence and intensity of intestinal parasites infection of plateau pika in LD,MD and HD(mean±SE). n=12,11 and 11 for males and n=13,19 and 10 for females in LD,MD and HD,respectively.The lowercase indicates significant difference(P <0.05).Because all the plateau pikas were infected with coccidia,the coccidian prevalence was not analyzed.The plateau pikas in MD were not infected with cestode,so the group was excluded during the statistical analyses.For abbreviations,see Fig.1
草地退化显著影响高原鼠兔对球虫(F2,70=17.07,P<0.001)、绦虫(F2,42=5.09,P=0.029)和线虫(F2,70=5.20,P=0.008) 感染强度(图3D、E、F)。中度退化草地球虫感染强度显著低于轻度和重度退化草地(P<0.001),而轻度与重度退化草地之间无显著差异(P>0.05)(图3D);重度退化草地绦虫感染强度显著低于轻度退化草地(P<0.05)(图3E);中度和重度退化草地线虫感染强度均显著低于轻度退化草地(P<0.001),但彼此间无显著差异(P>0.05)(图3F)。性别对球虫(F1,70=1.59,P=0.211)、绦虫(F2,42=2.62,P=0.113) 和线虫(F1,70=3.16,P=0.080)的感染强度均无显著影响。草地退化与性别的交互作用对球虫感染强度无显著影响(F2,70=0.69,P=0.505),但显著影响绦虫(F2,42=11.26,P=0.002) 和线虫(F2,70=5.41,P=0.007) 的感染强度。雌性在重度退化草地中对二者的感染强度均显著低于轻度退化草地(P<0.01),而雄性对二者的感染强度在不同退化草地中无显著变化(P>0.05)。
2.4 不同退化草地中高原鼠兔肝脏和肾脏指数
草地退化显著影响肝脏(F2,176=17.53,P<0.001;图4A) 和肾脏(F2,176=5.18,P=0.007;图4B)指数。重度退化草地中高原鼠兔的肝脏和肾脏指数均显著低于轻度和中度退化草地(P<0.05),而二者在轻度与中度退化草地之间无显著差异(P>0.05) (图4)。性别影响肝脏指数(F1,176=21.68,P<0.001),雄性个体肝脏指数显著低于雌性(P<0.05)。肾脏指数无性别差异(F1,176=1.59,P=0.209);退化草地与性别的交互作用对肝脏(F2,176=2.04,P=0.133) 和肾脏(F2,176=0.61,P=0.543)指数无显著影响。

图4 不同退化草地中高原鼠兔的肝脏指数和肾脏指数(平均值±标准误).轻度、中度和重度退化草地的雄性样本量分别为22、30和28,雄性样本量分别为32、41 和29.图中不同小写字母表示差异显著(P <0.05)Fig.4 The liver and kidney indices of plateau pika in LD,MD and HD (mean ± SE). n=22,30 and 28 for males and n=32,41 and 29 for females in LD,MD and HD,respectively.The lowercase indicates significant difference(P <0.05).For abbreviations,see Fig.1
3 讨论
天然免疫功能是机体抵抗病原体或寄生物感染的第一道屏障,对动物的生存至关重要(Franket al.,2000;Lochmiller and Deerenberg,2000;Ochsenbein and Zinkernagel,2000;Rickert,2005)。大量的研究中都以血清总IgG水平或白细胞数量衡量机体的天然免疫能力(Greiveset al.,2006;Xuet al.,2012;Forbeset al.,2014,2016)。尽管本研究中仅测定了血清总IgG水平,但实验结果与寄生物感染率和感染强度的降低相互印证地说明,在高寒草甸生态系统中,草地退化增强了高原鼠兔的天然免疫功能。
在自然界中,食物和捕食是影响动物免疫功能的重要因素(Nohret al.,1985;Lordet al.,1998;徐德立和王德华,2015;Shanget al.,2019)。大量研究表明,食物短缺可抑制动物的免疫功能(Nohret al.,1985;Lordet al.,1998;徐德立和王德华,2015),尤其是食物蛋白质对动物免疫功能具有重要影响。Davis 和Lochmiller (1995) 以及Lochmiller 等(1993)发现,即使中度限制蛋白质摄入水平,也会显著降低棉鼠(Sigmodon hispid) 的免疫功能。在本研究中,不同退化草地中高原鼠兔天然免疫力的变化,可能与其获取的食物蛋白质密切关联。在青藏高原地区,Li 等(2019)通过放牧控制实验的结果表明,高放牧强度可显著增加豆科植物的相对生物量。虽然豆科植物对家畜而言是有毒牧草,但对高原鼠兔而言,则是富含蛋白质的喜食食物(蒋志刚和夏武平,1985;樊乃昌和张道川,1996;刘伟等,2008)。我们的另一研究发现,在中度和重度退化草地中,高原鼠兔胃内容物蛋白质含量显著高于轻度退化草地,且草地退化为高原鼠兔提供了丰富的高蛋白质食物资源(未发表资料)。由此可见,重度退化草地通过提供丰富的高蛋白质食物,使其免疫力维持在较高水平。另一方面,高原鼠兔免疫力变化还可能与捕食风险有关。高原鼠兔具有警戒、观察和啄食式取食模式等反捕食行为特征(边疆晖和樊乃昌,1997;边疆晖等,1999),而过度放牧通过降低植被高度和盖度为高原鼠兔提供了开阔视野(施银柱,1983;边疆晖等,1994);反之,未退化或轻度退化的郁闭生境可通过抑制高原鼠兔的反捕食行为而增大其捕食风险,捕食风险的增大可引起个体的捕食应激反应,进而导致免疫功能降低(Boonstraet al.,1998;Bianet al.,2005;Sheriffet al.,2010,2011;Shanget al.,2019)。因此,草地退化对高原鼠兔天然免疫力的增强效应可能是退化草地中高蛋白质食物资源与低捕食风险的耦合效应所致。
血清总IgG 水平升高可说明动物个体的身体状况更加健康,抵抗寄生物感染的能力更强(Forbeset al.,2016)。球虫是高原鼠兔的主要肠道寄生物(曹伊凡等,2011;边疆晖等,2011),高原鼠兔为其直接宿主。本研究发现,中度退化草地高原鼠兔球虫感染强度显著低于轻度退化草地,但在重度退化草地中,没有发现显著变化。球虫感染强度不仅与免疫力相关,而且与种群密度正相关(聂绪恒等,2014)。例如球虫感染与红松鼠(Sciurus vulgaris) 种群密度呈显著正相关(Bertolinoet al.,2003)。在本研究的重度退化草地中,高原鼠兔洞口密度为751 个/公顷,是轻度退化草地的5.3 倍,在这种高密度条件下,高原鼠兔球虫感染强度通过粪便污染食物而加剧,加之高原鼠兔具食软便习性(刘全生和王德华,2004),从而导致重度退化草地中球虫感染强度增加,而绦虫和线虫需中间宿主才能完成其生活史,从虫卵到终宿主体内发育成成虫需较长时间,因而,其感染与宿主天然免疫力有更直接的关联性。
本研究发现,高原鼠兔肝脏和肾脏指数随草地退化而显著降低。在自然界中,引起动物肝脏和肾脏重量变化的因素众多,如光周期(张志强等,2007;郭子涵等,2022)、温度(张志强等,2007)、食物质量(Liu and Wang,2007;Zhanget al.,2012) 等。本研究在同一地区同一时期进行,因此,高原鼠兔肝脏和肾脏指数的变化可能与食物质量有密切关联。对于植食性小哺乳动物,在觅食过程中不可避免地摄入或多或少的植物次生化合物。肝脏和肾脏作为机体重要的解毒器官,其重量变化能够直接反映植物次生化合物的毒副作用。何岚等(2010) 研究发现,当饲喂高次生化合物含量(6%) 的食物时,可显著增加东方田鼠(Alexandromys fortis) 肝脏和肾脏指数,而高蛋白质含量食物(20%) 可减弱次生化合物对肝肾指数的负效应。在青藏高原地区,草地退化是通过长期过度放牧,家畜频繁地大量采食和践踏草地而导致,而频繁采食亦可导致植物的再生。相对于老的枝叶,再生的嫩枝叶富含较高的蛋白质和较低的次生化合物(白玉龙等,1999;李富娟和玉永雄,2006;戴鑫等,2012),这提示,相较于轻度退化草地,重度退化草地所提供的高蛋白质、低次生化合物食物可减轻植物次生化合物对高原鼠兔肝脏和肾脏的损伤,继而降低对高原鼠兔生长发育的不利影响,表现为重度退化草地的高原鼠兔肝脏和肾脏指数显著低于轻度退化草地。该结果也进一步佐证了重度退化草地高原鼠兔的较高天然免疫力可能与食物质量有关的推论。事实上,家畜选择性地采食可改善小哺乳动物食物质量(高蛋白质和低次生化合物) 的取食易化效应(feeding facilitation),在其他生态系统中已有大量的例证(Kruegeret al.,1986;Arsenault and Owen-Smith,2002;Cheng and Ritchie,2006)。
由此,验证了本文提出的草地退化可增强高原鼠兔的天然免疫功能,缓解植物次生化合物对高原鼠兔肝脏和肾脏损伤的假设。
在小哺乳动物种群波动中,免疫和寄生物是影响种群统计参数的重要因子(Boonstraet al.,1998;张志强和王德华,2005;Książeket al.,2014;Shanget al.,2019)。我们前期研究表明,应激—免疫—寄生物感染之间可形成恶性循环,导致种群死亡率增加,继而导致种群衰减(Yanget al.,2018),这从另一方面说明,重度退化草地可通过增强高原鼠兔天然免疫力,防止或削弱应激—免疫—寄生物感染之间的恶性循环,继而导致高原鼠兔种群数量的增加。基于上述结果与分析,本研究认为,草地退化可通过提高高原鼠兔天然免疫力、降低次生化合物对其生长发育的负效应,继而在高原鼠兔种群暴发中起促进作用。
猜你喜欢
广东农业科学(2022年5期)2022-07-21
动物医学进展(2022年7期)2022-06-10
环球时报(2022-04-07)2022-04-07
中国动物保健(2021年8期)2021-11-24
智慧健康(2021年5期)2021-03-30
现代牧业(2020年4期)2020-12-31
电脑报(2020年12期)2020-06-30
热带病与寄生虫学(2019年4期)2019-02-12
农民致富之友(2017年15期)2017-08-20
中国当代医药(2015年25期)2015-10-21
