
不同海拔高原鼠兔个性特征与SERT基因多态性关联
2022-09-21 14:37朱红娟李婧王苏芹唐琦蓝贤勇曲家鹏
兽类学报 2022年5期
朱红娟 李婧 王苏芹 唐琦 蓝贤勇 曲家鹏
(1 中国科学院西北高原生物研究所,西宁810001)(2 青海省动物生态基因组学重点实验室,西宁810001)(3 中国科学院大学,北京100049)(4 西北农林科技大学动物科技学院,杨凌712100)(5 青海三江源草地生态系统国家野外科学观测研究站,西宁 810001)
同一物种的动物在适应不同的栖息地环境过程中,往往会进化出差异显著的表型与遗传特征(Kawecki and Ebert,2004)。在表型分化方面,以往研究主要集中于比较不同生境下种群之间形态和生活史策略的差异(Des Rocheset al.,2018)。近年来,越来越多的研究表明,动物个体之间存在行为差异,这种差异随时间的推移和环境的改变持续存在且保持相对稳定(Réaleet al.,2007;Schuettet al.,2010),即个性(personality)。个性主要包括活跃性、探究性、勇敢性、攻击性以及社会性等(Réaleet al.,2010a)。个性是动物对变化环境长期适应与进化的结果(Wolf and Weissing,2012)。其中,探究性是个体面临新环境、新食物或新物体时的行为,反映了动物对捕食风险和新事物的反应能力。例如探究性强的披肩榛鸡(Bonasa umbellus)在冬季更容易寻找到温暖的生境作为避难所,存活率更高(Shipleyet al.,2020);探究性强的雌性欧亚红松鼠(Sciurus vulgaris)活动范围更广,获得的食物资源更多。由此可见,探究性可以影响动物的空间利用策略以及获得食物、配偶的概率(Wauterset al.,2021)。勇敢性是动物对非新奇环境中危险事物的反应。例如栖息在城市中的黑线姬鼠(Apodemus agrarius) 比栖息在农村的种群更大胆、探究性更强,能及时应对外界环境的快速变化(Dammhahnet al.,2020)。高活跃性的无斑雨蛙(Hyla arborea) 被捕食风险高,但获得食物资源的概率更大,获得交配的机会更多(Jaquiéryet al.,2010)。在不同生境下动物个性特征的不同参数相互关联,能更好地适应特定的环境(Dochtermannet al.,2015)。例如随着纬度的升高,切胸蚁(Temnothorax longispinosus)群体表现出更高的探究性和勇敢性,更低的攻击性;且探究性、勇敢性和攻击性之间的关联性从正关联转变为负相关(Segevet al.,2017)。
越来越多的证据表明,基因多态性影响动物的个性特征(Gartlandet al.,2022),DRD4和SERT基因被认为是调节个性特征变异的两个重要候选基因(Holtmannet al.,2016)。对野生大山雀(Parus major)、猕猴(Macaca mulatta)、白领姬鹟(Ficedula albicollis)、黄顶巧织雀(Euplectes afer)以及小鼠、人类等的研究均表明,多巴胺受体D4(DRD4)基因多态性与个体的探究性、勇敢性等有关(Klugeret al.,2002;Fidleret al.,2007;Korstenet al.,2010;Matthews and Butler,2011;Garamszegiet al.,2014;Muelleret al.,2014;Coyneet al.,2015)。5-羟色胺转运体(SERT)基因能调控神经元突触缝隙中5-羟色胺激素分泌,与动物的探究性等密切相关(Melke,2003;Savitz and Ramesar,2004;Riyahiet al.,2015)。例如灰海豹(Halichoerus grypus)SERT基因上的1 个位点突变与勇敢性显著相关。对分布在城市和农村两个乌鸫(Turdus merula)种群的研究发现,分布在农村的乌鸫种群探索性的进化压力与SERT基因外显子存在极显著关联(Mülleret al.,2013)。大山雀SERT启动子上SNP290 位点的遗传多态性与恐惧行为密切关联(Riyahiet al.,2022),而SNP226 位点上具有纯合子(AA)等位基因的雀形目鸟类探究性更强(Grunstet al.,2021),SERT基因敲除的小鼠探究性下降(Murphyet al.,2008)。总之,个性特征受到环境因素和遗传变异的共同制约,探讨不同生境下个性特征的差异及其遗传变异,对理解个性特征的产生及其适应与进化具有重要意义(Moironet al.,2020)。
世界上海拔最高的青藏高原,不同海拔地区的气候、环境等差异明显(Zhanget al.,2020),低温低氧、食物匮乏等环境因素不仅影响高原动物的生存与繁殖等生活史对策(Yuet al.,2018;Jiaet al.,2020),在物种形成、适应进化和遗传变异中也发挥着重要作用(Jianget al.,2021;Quet al.,2021),同时,环境因子还会影响动物的栖息地选择、行为、生理特征和遗传结构等(Stuberet al.,2022)。高原鼠兔(Ochotona curzoniae)隶属兔形目鼠兔科,是伴随青藏高原隆升进化而来的土著动物(Kojuet al.,2017),广泛分布于海拔3 200~5 300 m 的高寒草甸(Smithet al.,2010),也是青藏高原的特有种和关键种(Lai and Smith,2003)。随着海拔升高,为了适应不同的生境,高原鼠兔的代谢率、低氧低温适应对策等显著不同(Liet al.,2018;Speakmanet al.,2021),生活史对策、个性特征等亦存在显著差异(Quet al.,2013,2019)。为进一步理解高原鼠兔个性特征适应进化的分子机制,本研究比较了5个海拔地区高原鼠兔个性特征的差异;生物信息学分析的比对结果显示:高原鼠兔与其他动物DRD4基因的同源性与保守性较低,而SERT基因的同源性与保守性较高,因此选择高原鼠兔SERT基因进行mRNA 表达量及其遗传多态性检测,并分析基因多态性与个性特征的关联性,以期探讨不同海拔地区高原鼠兔个性特征的适应与变异机制,更好地理解高原动物对极端环境的适应与进化。
1 研究方法
1.1 实验动物采样地
2021 年7—8 月,高原鼠兔分别捕获于青海省海西蒙古族藏族自治州格尔木市昆仑山垭口(北纬38°33′,东经100°18′,海拔4 767 m),青海省果洛藏族自治州玛多县(北纬34°88′,东经98°23′,海拔4 193 m)、玛沁县(北纬34°45′,东经100°27′,海拔3 694 m),青海省海南藏族自治州贵德县拉脊山(北纬36°36′,东经101°41′,海拔3 733 m)和贵南县(北纬35°50′,东经100°66′,海拔3 321 m)。王建梅(2006) 发现栖息在海拔3 960 m 和4 790 m 地区的高原鼠兔体重、瘦素、甲状腺素T3含量以及肝脏线粒体蛋白含量、COX 活性等能量代谢相关参数均显著高于栖息在海拔3 200 m 的个体,个性特征与能量代谢密切相关(Careauet al.,2015),为更好地比较不同海拔地区高原鼠兔个性特征和SERT基因的差异,将格尔木和玛多采样地区划为高海拔地区,将拉脊山、玛沁和贵南采样地区划为低海拔地区。
分别在格尔木市昆仑山垭口捕获高原鼠兔21 只,在玛多、拉脊山、玛沁和贵南县各捕获高原鼠兔19 只。实验个体捕获后,立刻测定其个性特征。高原鼠兔的个性特征具有较高的重复性(Quet al.,2018,2019),因此本研究不再进行重复性测定。个性特征测定结束后,将高原鼠兔麻醉、处死(舒泰-50,LAT-STWS,北京莱艾特科技发展有限公司) 并解剖,将骨骼肌肌肉和下丘脑组织保存在液氮中,带回实验室直到进一步的提取和分析。所有实验动物操作均已获得中国科学院西北高原生物研究所实验动物伦理委员会的许可。
1.2 个性测定
1.2.1 温顺性
参考Montiglio 等(2012) 的方法,采用网袋实验测定高原鼠兔的挣扎状况,代表个体的温顺性。将捕获的高原鼠兔置于20 cm × 15 cm 的尼龙网袋中悬空,记录1 min 内个体静止不动的时间,即为温顺性(Réaleet al.,2007)。静止时间越久,个体越温顺(图1a)。测量结束后,清除网袋内的粪尿并用75%的乙醇进行清洁,风干后用于后续测定。
1.2.2 探究性
温顺性测定结束后,将高原鼠兔置于不透明的亚克力旷场(长×宽×高:65 cm×38 cm×45 cm)中,将旷箱底部划分为中央区与边缘区用于后续分析。在旷场中心正上方60 cm 处放置摄像机(Cannon EOS-1D X Mark Ⅲ,佳能(中国) 有限公司),记录3 min 高原鼠兔的各种行为(Careauet al.,2015)。保持拍摄过程安静、无干扰、光线均匀,避免外界环境干扰(图1b)。测定结束后,使用75%的乙醇清洁旷场,风干后用于下一次测定。使用Ethovision 14 动物运动轨迹跟踪系统(Noldus Information Technology,Wageningen,Netherlands)分析行为视频数据,统计3 min 内动物在旷场中的各项行为,包括移动距离和时间、中心区和边缘区停留时间和穿越频次等。动物在中心区域停留时间越长、穿越中心区域频次越多表示探索能力越强(Gould,2009)。
1.2.3 勇敢性
探究性测量结束后,将18 cm×10 cm×12 cm的隐蔽物随机置于旷场的角落,开口朝向旷场底部中心。高原鼠兔快速进入隐蔽物后,记录2 min内其第一次离开隐蔽物的时间,计为勇敢性。如果2 min 内高原鼠兔未离开隐蔽物,则勇敢性为120 s。记录的时间越短,则勇敢性越强(图1c)。
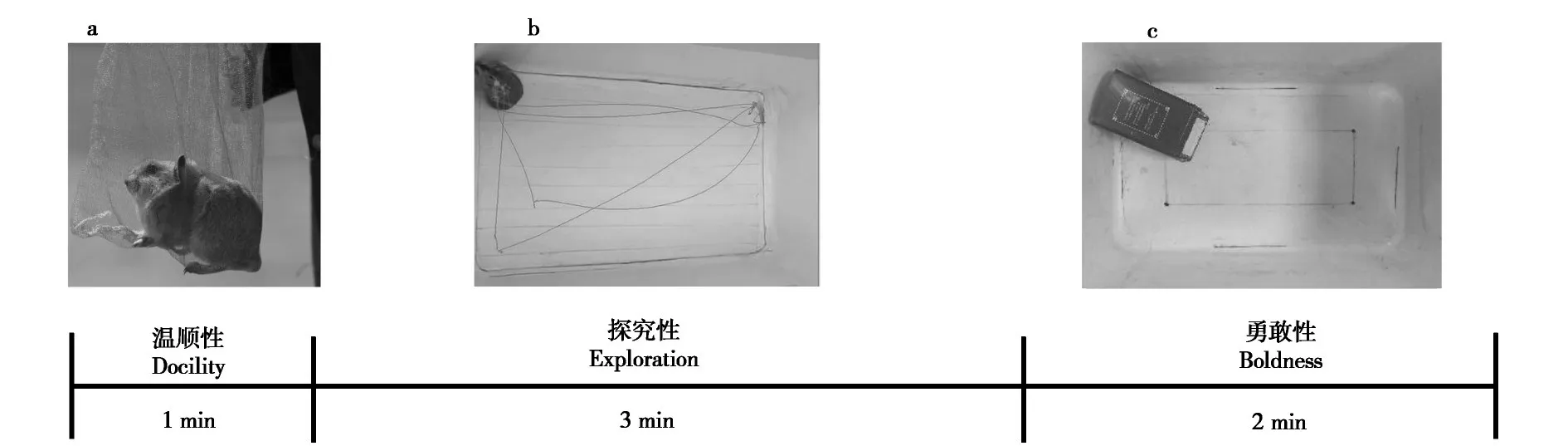
图1 高原鼠兔个性的测量流程图Fig.1 The flowchart for measuring the personality of the plateau pikas
1.3 SERT基因SNP位点筛选与基因分型
1.3.1 基因组DNA提取
采用酚氯仿法提取骨骼肌组织中的基因组DNA(李标,2019),使用NanoDrop 2000 分光光度计,按照A260/A280和A260/A230测定提取的基因组DNA的质量。随后将基因组DNA稀释至30 ng/μL,储存于-20°C的冰箱中用于后续研究。
1.3.2 引物设计
利用NCBI Primer Blast 在线工具(ncbi.nlm.nih.gov/tools/primer-blast/),跨外显子设计扩增引物,引物送至北京奥科鼎盛生物公司合成(表1)。
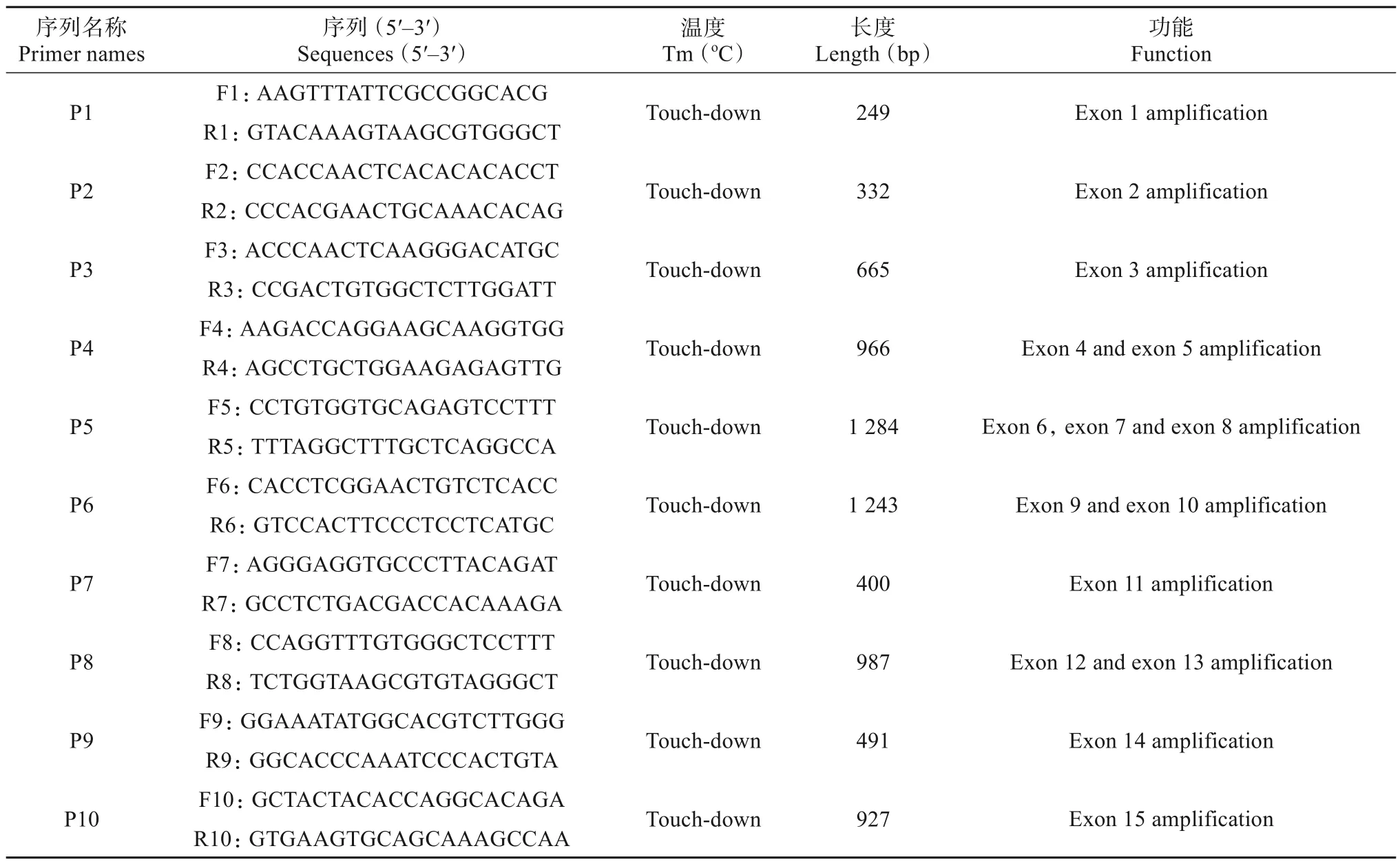
表1 高原鼠兔SERT基因外显子序列的PCR扩增引物Table 1 PCR amplification primers for exon sequence of plateau pika’s SERT gene
1.3.3 PCR扩增
在30 μL 反应体系中进行,包括2 × Taq Master mix (生工,西安) 15 μL,上下游引物各1 μL(10 μmol/μL),基因组DNA 模板1 μL(30 ng/μL)及ddH2O 12 μL 。PCR 扩增程序均采用Touch-Down PCR程序:95℃预变性5 min,94℃变 性30 s,68℃退火延伸20 s,72°C 条件下,延伸80 s,循环18 次,退火温度每个循环降1°C。用2%琼脂糖凝胶电泳检测PCR 产物,随后将有条带的PCR 产物送至生工生物工程有限公司进行测序。使用Chromas v2.4.1软件分析测序结果。
1.4 高原鼠兔SERT基因的mRNA表达检测
采用TRIzol 法从下丘脑组织中分离提取总RNA,使用NanoDrop 2000 分光光度计(Thermo Scientific,Waltham,MA,美国)和2%琼脂糖凝胶电泳检测总RNA的质量和完整性。然后,用HifairⅢ第一链cDNA 合成试剂盒(Yeasen 生物技术有限公司,上海) 将定量的RNA 反转录为cDNA。使用SYBR qPCR Master Mix (Vazyme,江苏)和引物对在荧光定量基因扩增PCR 检测系统(西安天隆科技有限公司)中进行实时荧光定量PCR。qRT-PCR 扩增体系(12 μL) 分别为:5 μL 的cDNA(20 ng/μL)、6 μL 的SYBR qPCR Master Mix、上游引物0.5 μL和下游引物0.5 μL。用于SERT表达分析的qRTPCR 引物序列列于表2。将SERT表达值标准化至GAPDH表达值(2-ΔΔCT分析)。
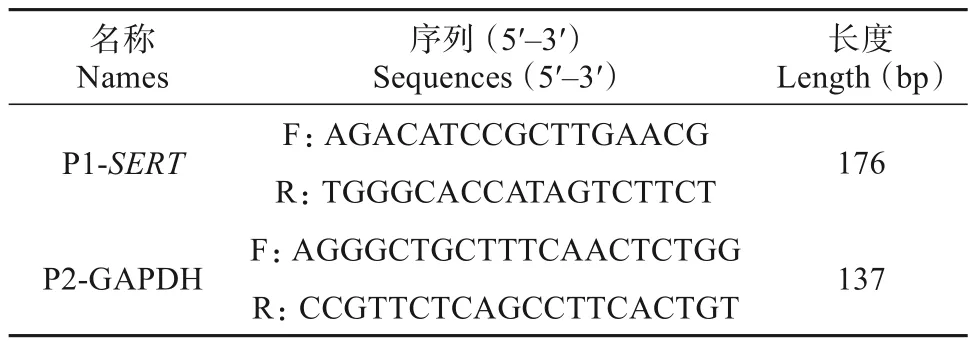
表2 高原鼠兔SERT基因qRT-PCR引物序列Table 2 qRT-PCR primer sequences of plateau pika’s SERT gene
1.5 数据处理
对行为数据进行正态分析(Shapiro-Wilk test)和方差齐性检验(Levene tests),随后采用主成分分析对探究性数据进行降维处理,采用单因素方差分析比较不同海拔之间高原鼠兔个性特征的差异。采用Chromas 2 软件对测序的结果进行校对比较,查看测序的结果,同时结合BLAST 分析确定SNP峰高测量以及等位基因频率的估算,用MWSnap测量各SNP位点等位基因的对应峰值高度。根据数据库SHEsis Main 里的方法(Nei,1973)统计基因分型频率、杂合度(He)、纯和度(Ho)以及多态信息含量(PIC)。不同海拔之间的高原鼠兔SERT基因型差异和定量表达差异采用单因素方差分析。采用卡方检验(Chi-Squared Test)分析不同海拔与基因型频率的关系。所有统计分析均在R 4.1.3上实现。
2 结果
2.1 不同海拔地区高原鼠兔个性特征差异
对高原鼠兔的探究性数据进行主成分分析,前两个主成分解释了81.36%的贡献率,将第一主成分作为探究性用于后续分析(表3)。高海拔格尔木和玛多地区高原鼠兔的探究性显著高于低海拔贵南地区(P<0.05)(图2A)。
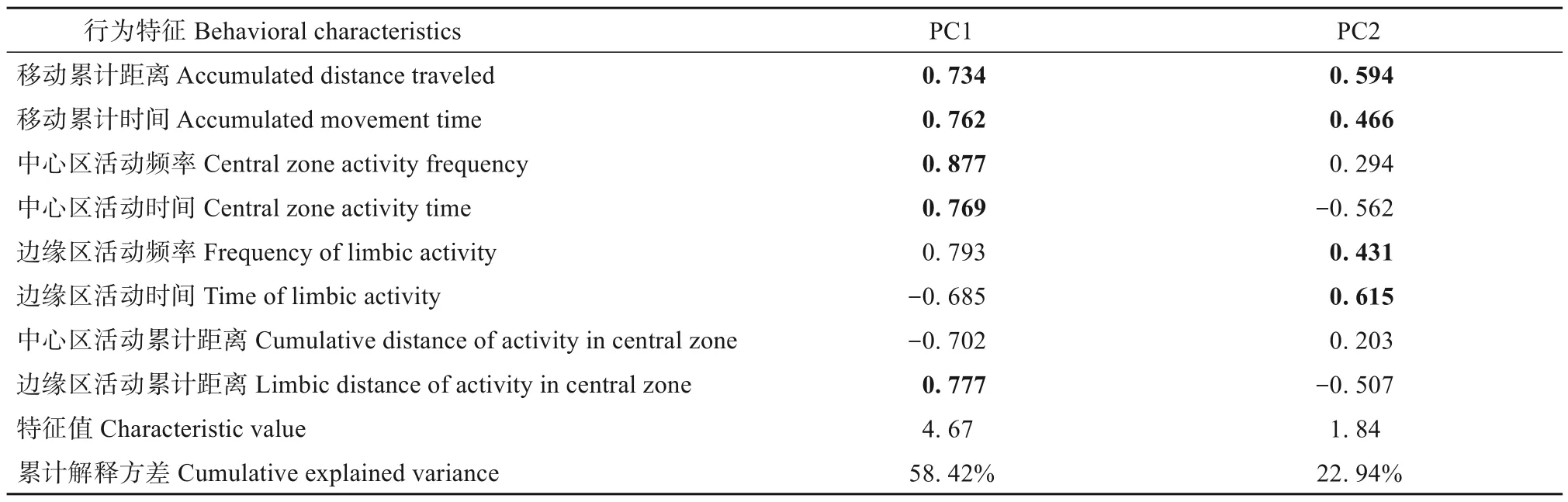
表3 8个高原鼠兔行为变量的主成分分析Table 3 Principal components analysis of measures of plateau pikas from the 8 behaviours
高海拔格尔木和玛多地区高原鼠兔的温顺性显著低于低海拔贵南和拉脊山地区,低海拔地区中贵南地区高原鼠兔的温顺性显著高于玛沁地区(P<0.05)(图2B)。
高海拔格尔木和玛多地区高原鼠兔的勇敢性显著高于低海拔玛沁和贵南地区(P<0.05)(图2C)。
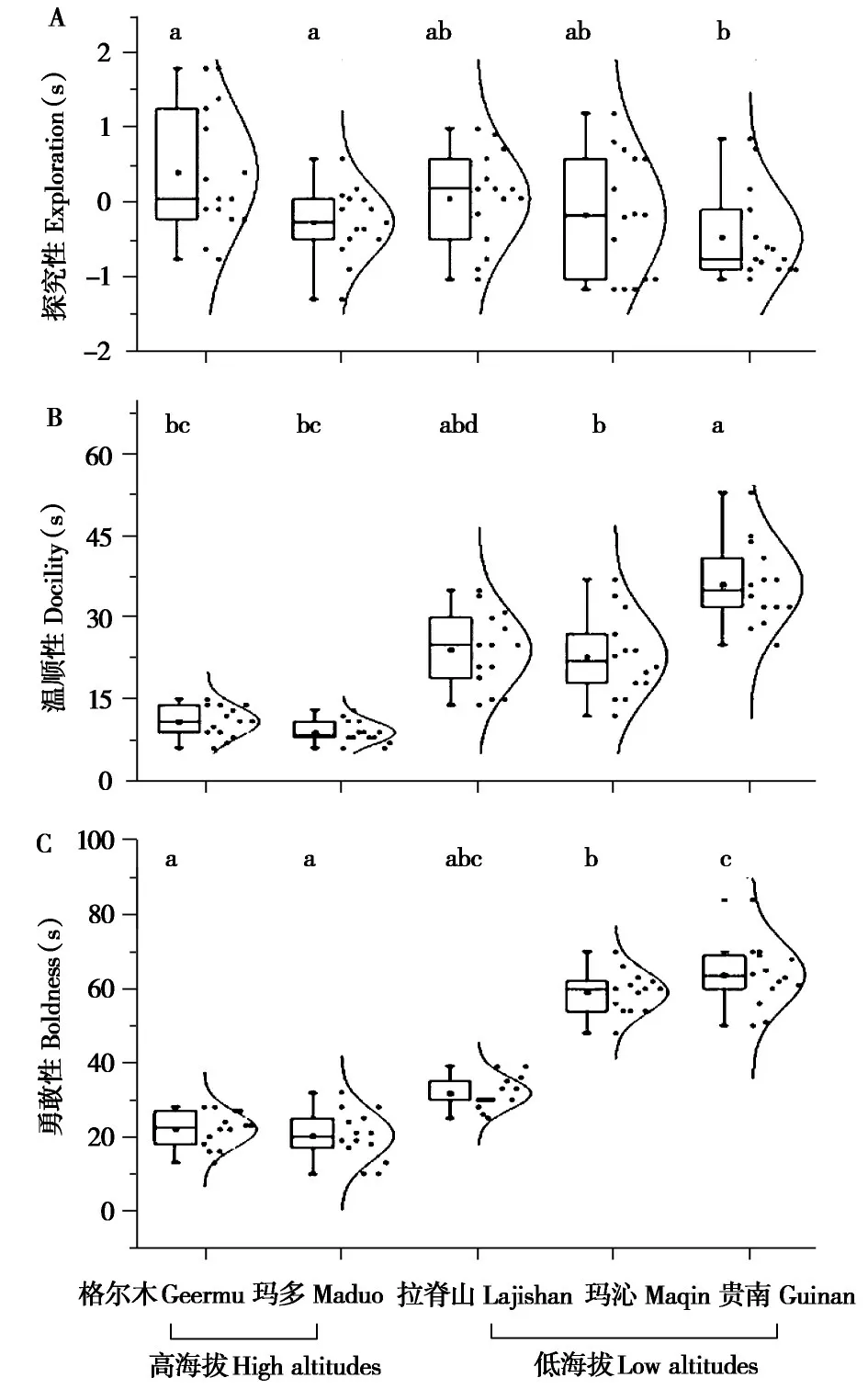
图2 不同海拔地区高原鼠兔个性特征的比较.柱上不同字母表示组间差异显著(P <0.05)Fig.2 Comparisons of personality traits of plateau pikas at different altitudes.Different letters above the bars indicate significant differences(P <0.05)
2.2 高原鼠兔下丘脑SERT基因的mRNA表达差异
实时荧光定量PCR 分析表明,下丘脑组织中SERT基因的mRNA 表达量在低海拔贵南地区显著高于低海拔玛沁和拉脊山地区(F13,182=25.43,P<0.05),低海拔玛沁和拉脊山地区显著高于高海拔玛多和格尔木地区(F13,182=28.52,P<0.05)(图3)。
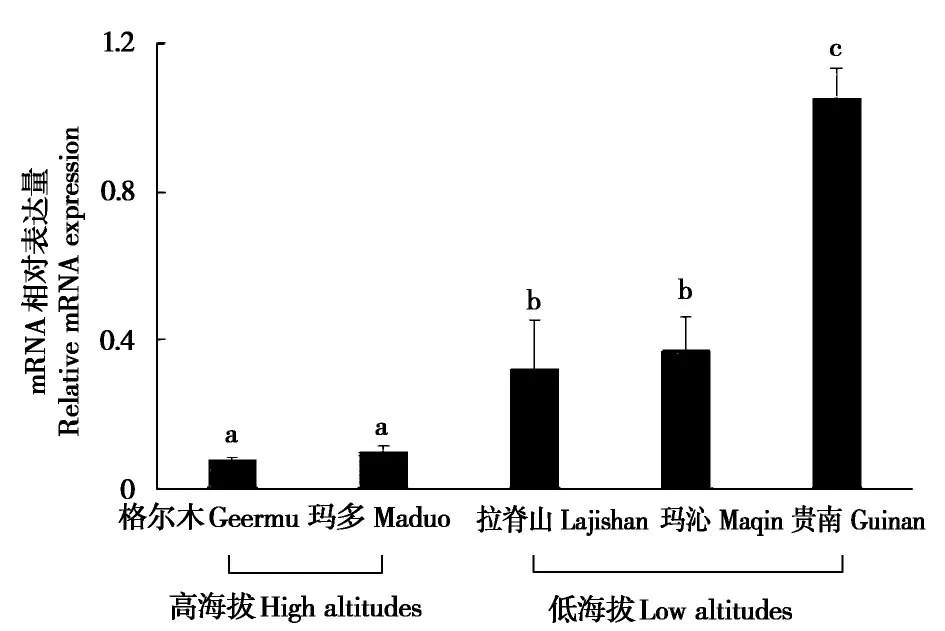
图3 不同海拔地区高原鼠兔SERT 基因的mRNA 表达差异.柱上不同字母表示组间差异显著(P <0.05)Fig.3 Differences in mRNA expression of SERT gene in plateau pikas at different altitudes.Different letters above the bars indicate significant differences(P <0.05)
2.3 高原鼠兔SERT基因多态性
高原鼠兔SERT基因不同目标基因片段的凝胶电泳结果如图4所示,各PCR片段实际大小与预期相符。使用Chromas v2.4.1 软件查看测序结果,在鼠兔SERT基因上发现6 个SNP 变异,其中5 个SNP 位于第3 外显子(图5a),根据其cDNA 位置,分别命名为c.C294T、c.G341A、c.C392T、c.C422T和c.T439C;另外1 个SNP 位于第5 外显子,命名为c.A1063C (图5b)。c.C422T 和c.A1063C 为 同义突变,分别编码丝氨酸(Ser) 和丙氨酸(Ala);c.T439C(Val147Ala)为错义突变(Val >Ala),其余3个SNP位于非翻译区。
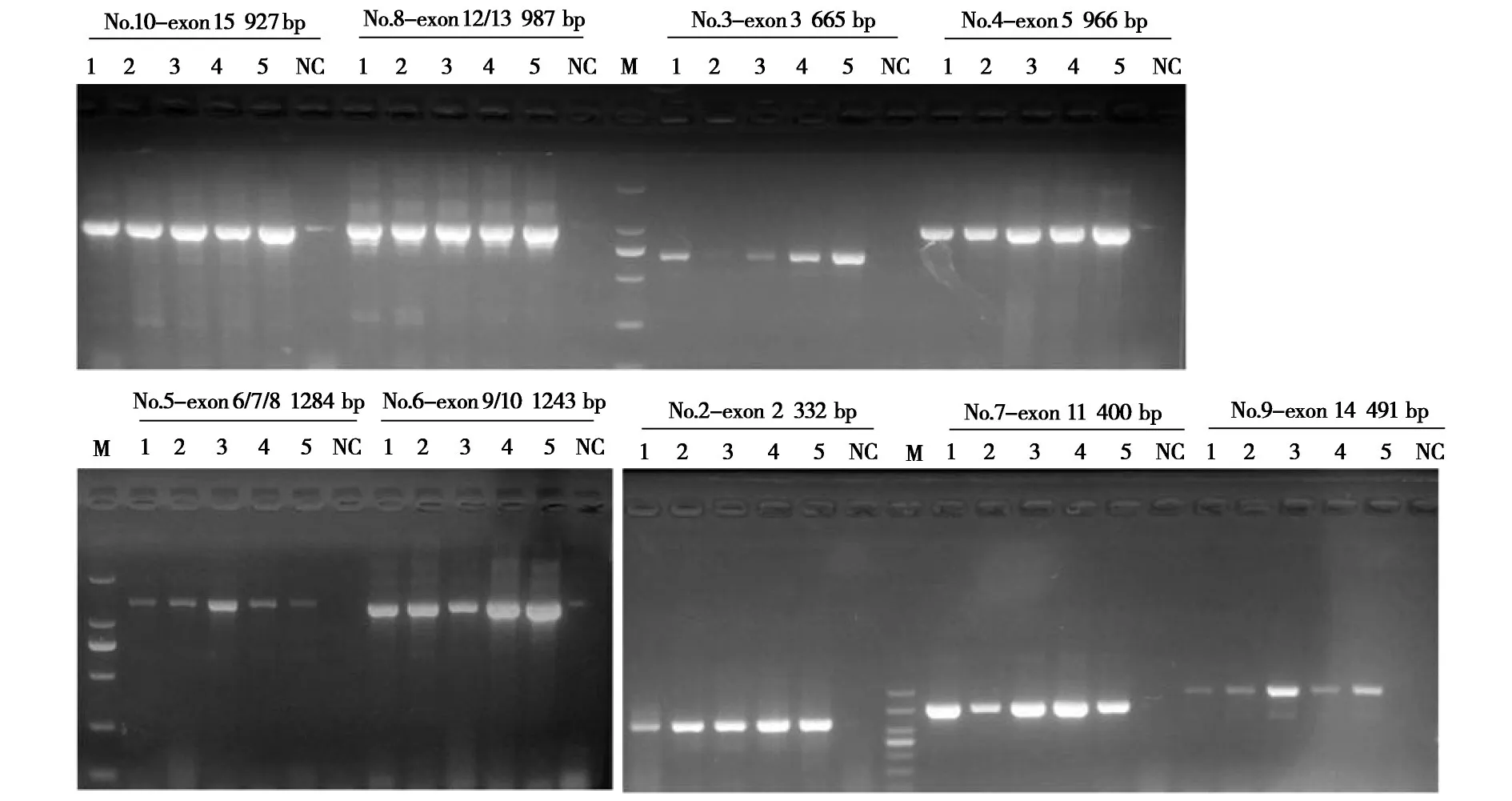
图4 高原鼠兔SERT外显子PCR产物扩增电泳图Fig.4 Electrophoretic diagram of SERT exon PCR product amplification in plateau pikas
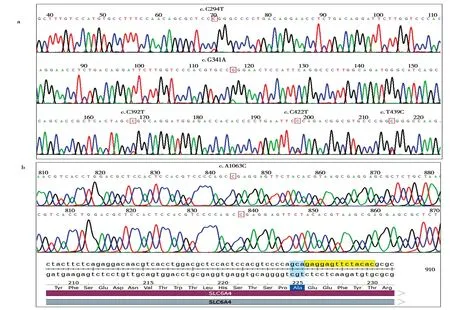
图5 高原鼠兔SERT外显子的多态性.a:第5外显子SNP的PCR产物测序;b:第3外显子SNP的PCR产物测序Fig.5 The polymorphism of SERT exons in plateau pikas.a:PCR product sequencing of SNPin exon 5;b:PCR product sequencing of SNPin exon 3
2.4 高原鼠兔SERT基因多态性与海拔关联分析
对SERT基因上6 个SNP 突变位点进行基因型频率、杂合度、纯和度以及多态信息含量统计。第5 外显子上的c.A1063C 突变位点存在AA、AC和CC 3 种基因型;第3 外显子上的c.C294T、c.C392T、c.T439C 和c.C422T 突变位点存在CC、CT 和TT 3 种基因型,c.G341A 突变位点存在AA、GA和GG 3种基因型(表4)。
对6 个SNP 位点与海拔之间的关系进行分析,结果表明,第3 外显子上的c.C294T、c.G341A、c.C392T、c.C422T、c.T439C 与海拔之间均没有显著相关性(P>0.05),第5外显子上的c.A1063C与海拔之间存在显著相关性(χ2=60.87,P=0.01)(表4) 。高海拔格尔木和玛多地区CC 基因型的分布频率显著高于低海拔地区(P <0.05)(图6)。
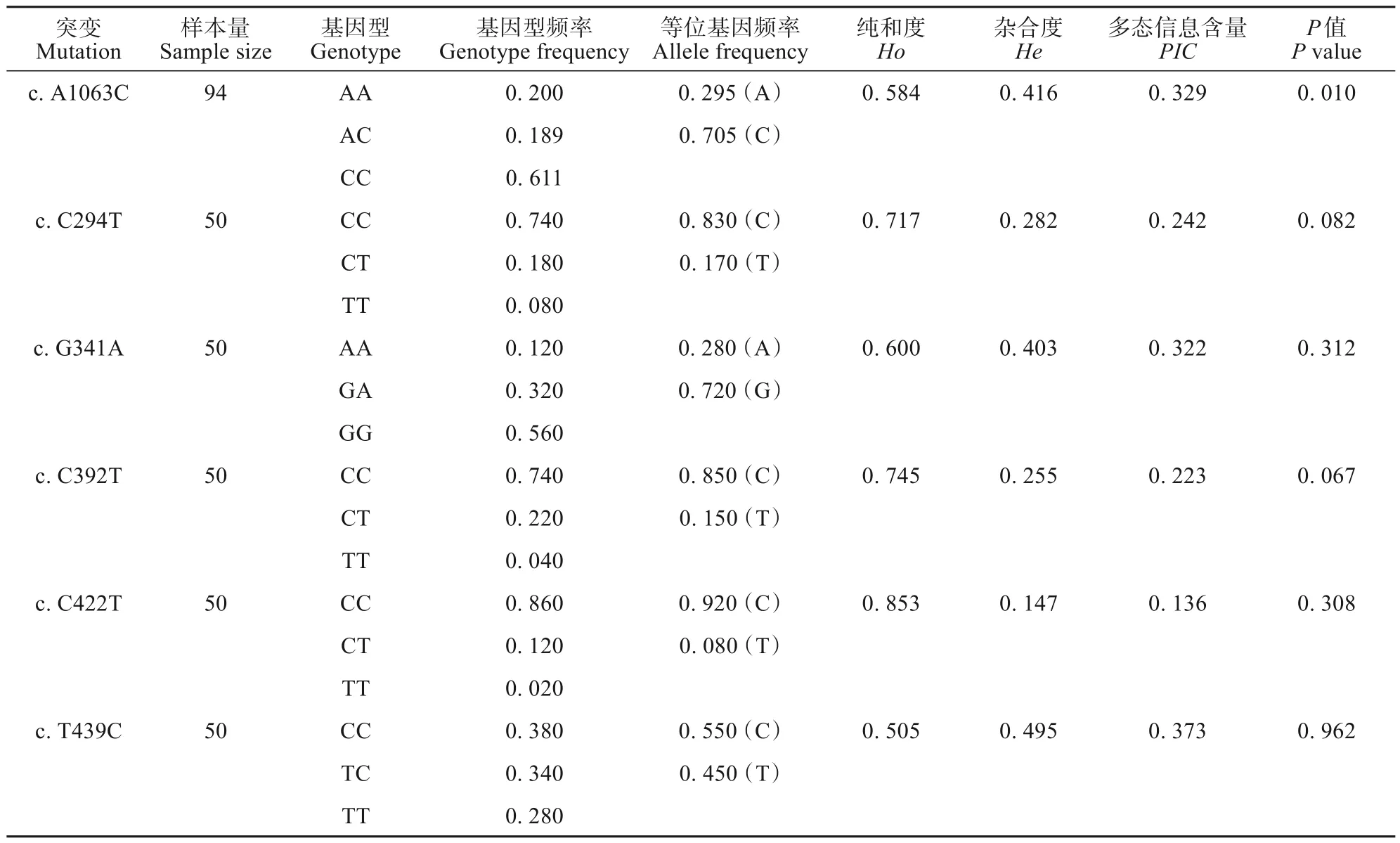
表4 高原鼠兔SERT基因等位基因频率估算Table 4 Estimation of allelic frequency of plateau pika’s SERT gene
2.5 c.A1063C位点基因型与个性特征关联分析
高原鼠兔SERT基因的c.A1063C 位点不同基因型(AA,AC 和CC)与探究性或勇敢性均无显著相关性(P>0.05),但与温顺性显著相关(P<0.05),c.A1063C 杂合子(AC 型)的高原鼠兔比纯合子(AA或CC型)个体更温顺(图7)。

图7 高原鼠兔SERT基因c.A1063C不同基因型与个性特征的相关分析.*表示不同基因型间有显著差异(P <0.05).A-D分别表示探究性(移动时间)、探究性(中心区活动频率)、勇敢性和温顺性Fig.7 Correlations between different genotypes of c.A1063C within the SERT gene and personality characteristics in plateau pikas.*represent a significant difference at 0.05 level between different genotypes (P <0.05).A-D represent exploration (accumulated movement time),exploration(central zone activity frequency),boldness and docility,respectively
3 讨论
本研究比较了不同海拔地区高原鼠兔种群个性特征和SERT基因表达量的差异,以及SERT基因多态性和个性特征之间的相关性。结果表明,高海拔地区高原鼠兔的探究性和勇敢性均显著高于低海拔地区,而温顺性相反;在SERT基因上发现了6个SNP位点,其中5个SNP位于第3外显子,1 个SNP 位于第5 外显子(c.A1063C),高海拔地区c.A1063C 位点CC 基因型的分布频率显著高于低海拔地区。进一步研究发现,c.A1063C 不同基因型与探究性或勇敢性均无显著相关,但与温顺性显著相关,低海拔地区SERT基因的mRNA 表达量显著高于高海拔地区。这表明不同海拔地区高原鼠兔温顺性的差异可能与SERT基因有关。
3.1 不同海拔高原鼠兔个性特征差异
根据动物行为综合征假说(Pace-of-life syndrome hypothesis,POLS),动物的行为与生理特征、生活史对策等密切相关,共同应对外界环境变化(Smith and Blumstein,2008;Réaleet al.,2010b;Rangassamy,2015)。例如探究性较强的大山雀在面临外界环境变化时,能迅速迁移、更好地适应新环境,有利于个体发育或繁殖(Réaleet al.,2007)。探究性强的西伯利亚花栗鼠(Tamias sibiricus)比探究性弱的个体生育力高,但被捕获率更大、死亡率更高(Boyeret al.,2010)。在新环境中探究性强的捷蛙(Rana dalmatina),往往比探究性弱的个体扩散距离更远(Urszánet al.,2015)。勇敢的巨型海蟾蜍(Rhinella marina)在面临高捕食风险时,更容易暴露在危险环境中,而胆怯的个体倾向于逃避(Narayanet al.,2013)。在食物丰富的年份,活跃的雌性北美红松鼠(Tamiasciurus hudsonicus) 子代生长速率更快,而在食物匮乏的年份则相反,表明活跃性在北美红松鼠子代发育过程中发挥重要作用(Boonet al.,2010)。黄腹旱獭(Marmota flavaviventris) 越温顺,被捕获的概率越低(Petelleet al.,2013)。动物为适应不同的栖息地环境,进化出了相应的生活史策略,产生了“快—慢生活节奏连续谱”(Ricklefs,2000)。例如栖息在较冷水域中的食蚊鱼(Gambusia holbrooki) 进化出较慢的生活史对策,性成熟晚、繁殖率低;而栖息在温暖水域的种群进化出较快的生活史对策,性成熟早、繁殖率高,但存活率低(Polverinoet al.,2018)。高海拔地区高原鼠兔的探究性和勇敢性均显著高于低海拔地区,而温顺性相反。这与高海拔地区红领带鹀(Zonotrichia capensis)具有较高探究性的结论一致(Pobleteet al.,2018)。快生活节奏的种群个体探究性、攻击性和活跃性强,代谢率高,反之亦然(Del Giudice,2020)。在种群间水平上,高海拔地区高原鼠兔种群属于慢生活节奏种群,探究性弱、繁殖力低;而低海拔地区高原鼠兔属于快生活节奏种群,探究性强、繁殖力高(Quet al.,2012)。动物的探究性与其栖息地的食物、领域和配偶资源紧密相关(Herbornet al.,2014)。青藏高原高海拔地区草地生物量更加匮乏(刘哲等,2015;Wanget al.,2007),植被高度和鼠兔可利用食物资源亦显著低于低海拔地区(谭春桃等,2020),但高海拔地区鼠兔的静息代谢率更高(Zhuet al.,2022),高原鼠兔为了满足更多的能量需求,进化出了更高的探究性和勇敢性,以获取更多的食物资源和生存机会。
3.2 高原鼠兔SERT 基因多态性与个体特征的关联分析
环境和遗传因素共同影响动物的个性特征(Crostonet al.,2015;Dochtermannet al.,2015;Charmantieret al.,2017)。高原鼠兔SERT基因第5 外显子SNP(c.A1063C)与海拔显著相关,c.A1063C 位点不同基因型与探究性或勇敢性均无显著关联,但与温顺性显著相关。这与Edwards等(2015)的研究结论一致,其研究发现塞岛苇莺(Acrocephalus sechellensis)的探究性和勇敢性均与SERT基因变异没有明显关联。个性特征复杂的遗传机制需要大量的样本来验证,有关高原鼠兔SERT不同位点SNP 的基因型与温顺性之间的关系还需要进一步研究。尽管未发现SERT基因SNP 位点突变与不同海拔地区高原鼠兔探究性和勇敢性的关联,但不排除可能受到生长发育或其他外部因素调节的影响(Deans and Maggert,2015)。例如蓝山雀(Cyanistes caeruleus) 仅在雏鸟时期攻击性表现出较强的遗传相关性,可能受到生长发育相关基因表达变化的影响(Class and Brommer,2015)。高海拔格尔木和玛多地区c.A1063C 基因的CC 基因型分布频率较高,可能导致不同环境和遗传因素共同影响高原鼠兔的个性特征(Weisset al.,2021)。
本研究中,SERT基因第5 外显子SNP 位点c.A1063C 与海拔显著相关,并且c.A1063C 为同义突变。同义SNP 不改变SERT 蛋白质的氨基酸序列(Tilgar and Koosa,2019),因此不影响SERT 的结构和功能。但是,最近研究表明:同义SNP 可以通过影响蛋白质的转录、剪接和折叠等,进而影响蛋白质的表达水平和功能(Leeet al.,2020)。同义SNP 可能与功能性变体有关。例如同义SNP影响人类疾病相关蛋白质的表达水平和功能(Sharmaet al.,2022)。有关大山雀SERT和(或)DRD4基因多态性的研究大多数涉及同义SNP,且将SNP与个性特征变异联系在一起,表明同义SNP 可能与个性特征的遗传变异有关(Muelleret al.,2013;Timmet al.,2018,2019)。低海拔地区高原鼠兔SERT基因表达量显著高于高海拔地区,而温顺性亦显著高于高海拔地区,表明SERT基因表达可能影响动物的个性特征,其分子关联机制有待进一步研究。
综上所述,随着海拔升高,高原鼠兔的探究性和勇敢性增加,温顺性降低,有助于在相对较为恶劣的高海拔地区获得更多的食物资源,增加生存机会,其中SERT基因多态性可能与不同海拔区高原鼠兔的温顺性差异有关,反映了个性特征海拔差异可能潜在的分子关联机制。由此提示,进一步深入研究自然选择压力下动物个性特征的遗传变异,将有助于更好地揭示个性特征产生的原因和适应进化机制。
致谢:感谢本课题组技术员来有荣和青海大学韩宗元同学等在野外工作中提供的帮助。
猜你喜欢
军事文摘(2022年14期)2022-08-26
散文诗(2021年22期)2022-01-12
中国生殖健康(2020年4期)2021-01-18
环球时报(2019-05-23)2019-05-23
学周刊·中旬刊(2017年1期)2017-02-05
青年时代(2016年32期)2017-01-20
戏剧之家(2016年4期)2016-03-25
艺海(2015年3期)2015-08-27
湖北农业科学(2014年11期)2014-09-10
