
茴香薄翅野螟病原真菌鉴定及其致病力评估
2022-11-04 13:54马有花来有鹏
江西农业大学学报 2022年5期
马有花,来有鹏
(青海大学 三江源生态与高原农牧业国家重点实验室/青海省农业有害生物综合治理重点实验室,青海 西宁 810016)
【研究意义】茴香薄翅野螟(Evergestis extimalisScopoli,1763)又称茴香螟、油菜螟、角野螟,在分类上属于鳞翅目(Lepidoptera),野螟蛾科(Crambidae),薄翅野螟亚科(Evergestinae),薄翅野螟属(Evergestis),寄主主要有油菜、茴香、白菜、甘蓝等[1]。茴香薄翅野螟是青海高原春油菜田的毁灭性害虫,年均发生面积为油菜种植总面积的20%以上,造成严重的经济损失[2]。茴香薄翅野螟对其寄主植物主要以幼虫卷叶,取食心叶、种芽的方式为害,在油菜上主要以幼虫啃食嫩茎、钻蛀油菜角果取食籽粒的方式为害[2-3]。目前生产上以化学防治为主,并且防治时使用的农药品种单一[3],由于该虫已对常用农药产生较强的抗性,再加上农药残留对于人畜的安全隐患,化学农药防治已受到了一定限制[4-5],因此探索一种高效、低毒、低残留的微生物杀虫剂迫在眉睫。【前人研究进展】昆虫病原真菌作为一类广泛存在于自然界的重要资源,能感染昆虫并在其体表或体内增殖,使昆虫发病甚至死亡[6],进而高度有效地调控昆虫种群数量动态。现阶段,在利用昆虫病原真菌防治农林害虫方面,半知菌亚门的白僵菌和绿僵菌因具有寄主范围广、致病力强等优点而受到各界的广泛关注[7]。王海鸿等[8]研发了防治西花蓟马(Frankliniella occidentalisPergande)的150 亿孢子/g 球孢白僵菌可湿性粉剂,在该害虫的生物防治中取得了很好的效果并已商业化;Shehzad 等[9]研究表明在接种物直接喷施法及浸叶法2 种不同的施用方法下,球孢白僵菌(Beauveria bassiana)和金龟子绿僵菌(Metarhizium anisopliae)对小菜蛾(Plutella xylostella)2 龄和3 龄幼虫都有较强的毒力。毛霉属真菌广泛分布在自然界中,寄生于动植物或腐生于土壤、动植物残体等基物[10-11],部分毛霉菌是一些鳞翅目、鞘翅目和双翅目昆虫的病原体[12]。Heitor[13]从甘蓝夜蛾(Mamestra brassicae)幼虫个体上分离出一株有致毒作用的冻土毛霉(Mucor hiemalis);Reiss 等[14]发现冻土毛霉(M.hiemalis)对卤虫(Artemia salina)具有致病性;Konstantopoulou 和Mazomenos[15]从患病的橄榄果实蝇(Bactrocera oleae)蛹和蛀茎夜蛾(Sesamia nonagrioides)幼虫中分离到6种昆虫病原真菌,试验表明2株冻土毛霉(M.hiemalis)对地中海实蝇(Ceratitis capitata)幼虫有较好的防治效果。Konstantopoulou 等[16]研究发现冻土毛霉(M.hiemalisSMU-21)的代谢提取物具有杀虫活性。【本研究切入点】目前关于茴香薄翅野螟的病原真菌未见文献报道。【拟解决的关键问题】本研究从青海省大通回族土族自治县采集的茴香薄翅野螟幼虫尸体上分离纯化得到了一株致病真菌,通过形态学及ITS 分子鉴定确定其分类地位,研究菌株对茴香薄翅野螟的致病性,以期为生防剂的研发、绿色防控提供理论依据。
1 材料与方法
1.1 供试虫源及培养基
在青海省西宁市大通县、湟中县采集茴香薄翅野螟卵及幼虫,置于20 ℃,相对湿度80%的条件下饲养。
1.2 病原真菌的分离纯化
参考Hallouti 等[17]的方法,将感病的茴香薄翅野螟幼虫,浸入含有0.3%次氯酸钠的蒸馏水溶液中消毒1 min,接着用无菌蒸馏水冲洗4 次后放入底部铺有无菌滤纸的一次性培养皿中保湿培养。待虫体长出菌丝或孢子时,在超净工作台中挑取少量菌丝或孢子接种于PDA 培养基,放置25 ℃培养箱培养3 d。将长好的菌丝挑入新的PDA平板,如此往复纯化4次后的菌株于4 ℃冰箱保存。
1.3 病原真菌的适宜培养基筛选
从8种供试培养基(表1)中筛选出适宜病原真菌生长的培养基,以供后续试验使用。将活化的菌株打菌饼(d=8 mm)接种于各培养基平板中心,于25 ℃恒温箱培养3 d,每天观察菌落形态并用十字交叉法测量菌落直径,计算日平均长速;3次重复,培养6 d后在无菌条件下用无菌水洗脱孢子,4层纱布过滤,制成孢子悬浮液,于光学显微镜下用血球计数板计数产孢量。
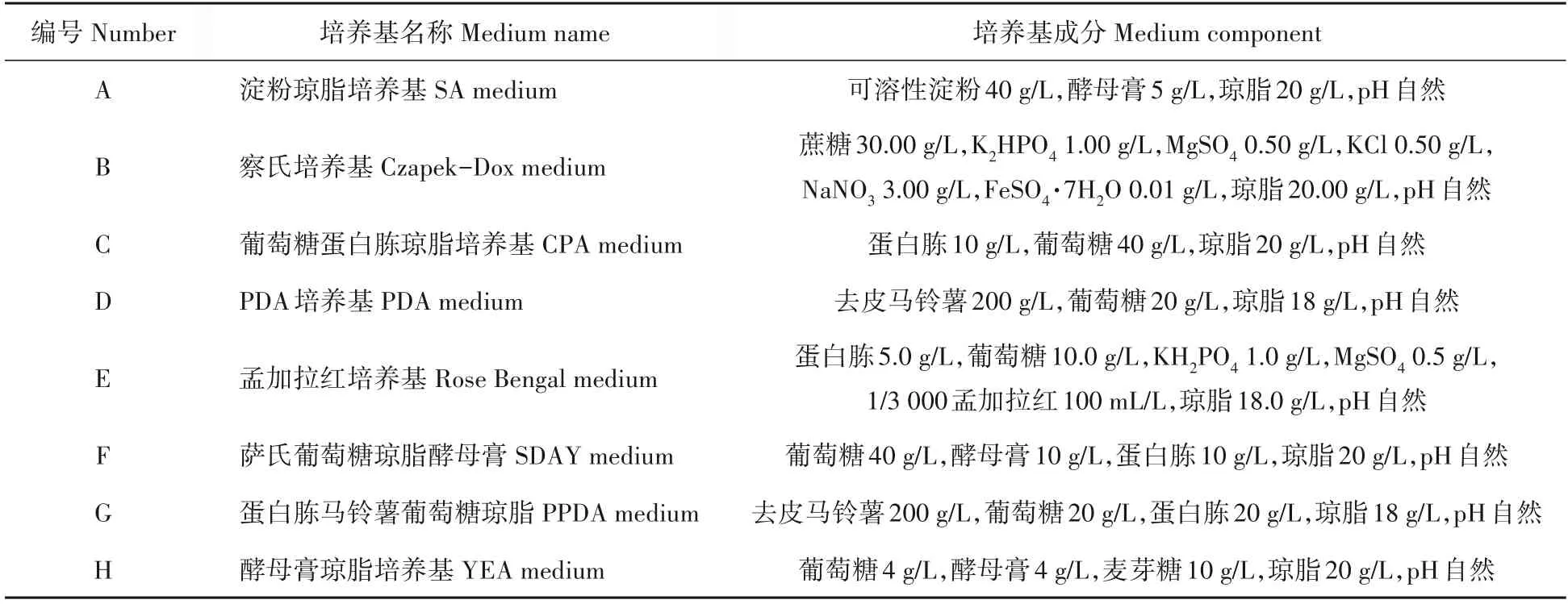
表1 供试培养基及其配方Tab.1 Test medium and its formula
1.4 病原真菌的形态学观察
取活化的菌株打菌饼(d=8 mm)接种于PDA 平板,将灭菌盖玻片斜插入培养基中,于25℃恒温箱培养2~3 d,经乳酸酚棉蓝染色液染色后,用光学显微镜观察菌株形态,参照真菌鉴定手册[18],对病原菌进行形态学鉴定。
1.5 病原真菌的分子生物学鉴定
用CTAB 法[19]提取菌株基因组DNA,运用真菌通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)、ITS4(5′-TCCTCCGCTTATTGATATGC-3′)进行ITS-PCR 扩增,引物均由上海生工生物有限责任公司合成。PCR 扩增反应体系:ITS1/ITS4 引物各1 μL、模板DNA 1 μL、PCR MasterMix 12.5 μL、ddH2O 9.5 μL,反应条件:95 ℃预变性5 min;94 ℃变性30 s,57 ℃退火30 s,72 ℃延伸90 s,30 个循环72 ℃延伸10 min。将扩增产物送往上海生工生物有限责任公司进行测序。
将菌株的序列测定结果提交至GenBank 核酸序列数据库中,进行BLAST 比对[20],从中选取同源性较高的序列,利用MEGA 7.0[21]软件,按照邻接法,运行1 000 次bootstrap 计算,构建系统发育树(Neighbor joining tree)对菌株进行分类鉴定。
1.6 病原真菌的致病性测定
依据柯赫氏法则验证菌株对茴香薄翅野螟幼虫的致病性。供试菌株接种于SDAY 平板,25 ℃活化培养6 d,在超净工作台用10 mL 无菌水溶液(含0.5 mL 0.05%吐温80)洗脱孢子,4 层纱布过滤菌丝,制成孢子悬浮液,用血球计数板调节孢子液浓度为1×107孢子/mL,10 mL 无菌水溶液(含0.5 mL 0.05%吐温80)为空白对照。采用浸渍法进行生物测定。用毛笔挑选2~5 龄幼虫,放入孢子悬浮液中,浸渍20 s挑出,晾干后,移入自制的养虫盒(8 cm×14 cm×5 cm)中,以新鲜的白菜饲养。各处理15 头幼虫,重复4次,每天观察幼虫存活情况,将死亡幼虫于铺有湿润滤纸的培养皿(d=9 cm)中进行观察[22],虫体长出白色菌丝者即为感染。
1.7 不同孢子浓度对茴香薄翅野螟2龄幼虫的毒力
将培养8 d 的菌株按1.6 所述方式制孢子液后,稀释成105,106,107,108孢子/mL 悬浮液,选取同等大小的2 龄幼虫浸入不同浓度孢子悬浮液中20 s,自然风干后,于养虫盒中饲喂白菜并定期更换菜叶。各处理15头幼虫,重复3次,按时观察记录幼虫死亡情况,将死亡幼虫保湿培养[22]。
1.8 数据处理

利用Excel整理数据,SPSS 26.0软件进行处理分析,统计处理幼虫的累计死亡率和累计校正死亡率,求得回归方程、致死中时及致死中浓度,用Graphpad prism 8.0作图。
2 结果与分析
2.1 病原真菌的适宜培养基筛选
据图1及表2可知,病原菌在试验所选的8种培养基上都能生长。菌株在孟加拉红培养基、SDAY 培养基、PPDA 培养基中菌丝发达致密,边缘整齐;在CPA 培养基、PDA 培养基、YEA 培养基中菌丝前期致密,后期稀疏;相对其他培养基而言,菌株在SA培养基和察氏培养基中长势较差、稀疏。
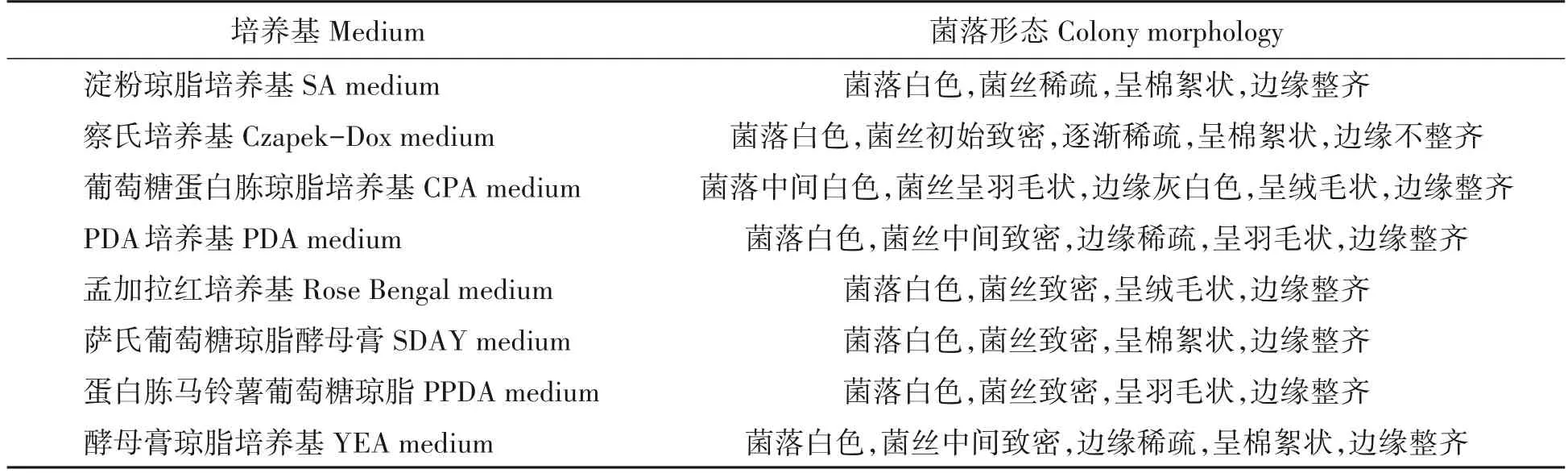
表2 菌株在不同培养基上的菌落形态Tab.2 Colony morphology of strain on different media
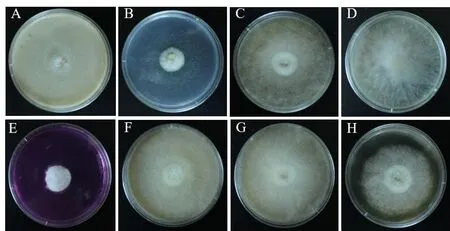
图1 菌株在不同培养基上的菌落形态Fig.1 Colony morphology of strain on different media
由图2所示,菌株在不同培养基上的日平均长速和产孢量存在差异。菌株在PPDA 培养基上生长速度最快,达2.81 cm/d,SDAY 培养基和CPA 培养基次之,长速分别为2.80 cm/d、2.71 cm/d,彼此无显著差异,孟加拉红培养基中菌丝生长最慢,仅为0.50 cm/d;培养6 d 后病原真菌在SDAY 培养基上产孢量最高,为2.9×107孢子/mL,孟加拉红培养基及PPDA 培养基次之,分别是2.8×107孢子/mL、2.25×107孢子/mL,察氏培养基产孢量最低,为0.06×107孢子/mL,显著小于其他培养基。因此,综合菌落形态、生长速度及产孢量,选择SDAY 培养基为最适培养基。

图2 菌株在不同培养基上的日长速及产孢量Fig.2 Comparison of growth rate and sporulation yield of strain on different media
2.2 病原真菌的形态学特征
菌株在PDA培养基培养2~3 d,菌丝生长迅速,呈白色羽毛状,中间致密,边缘稀疏,菌落圆形(图1A);在光学显微镜下观察到菌株菌丝无隔,孢囊梗直立单生,顶生球状孢子囊,含大量孢囊孢子,孢子椭圆形、圆形(图1B)。根据以上观察结果,参考魏景超[18]的研究,初步鉴定该菌为毛霉属真菌。
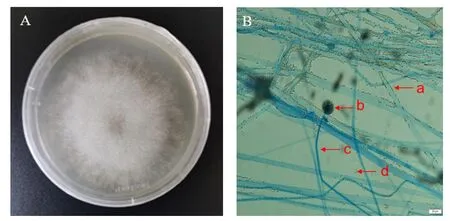
图3 病原真菌生物学特征Fig.3 Biological characteristics of pathogenic fungi
2.3 病原真菌的分子生物学鉴定
通过测序得到535 bp DNA 序列,提交至NCBI 数据库(登录号:OK427267)进行BLAST 比对,结果表明分离株与冻土毛霉(Mucor hiemalis)对应序列的同源性高达100%,构建系统发育树(图4),发现菌株与M.hiemalis聚为一类,支持率为100%,结合形态学和ITS 序列相似性分析,最终确定该菌为冻土毛霉,菌株编号QH01。

图4 基于ITS序列构建的菌株QH01系统发育树Fig.4 Phylogenetic tree of strain QH01 based on ITS sequence
2.4 病原真菌的致病性分析
根据柯赫氏法则验证菌株对茴香薄翅野螟幼虫的致病性,结果表明:菌株QH01对2~5 龄幼虫均有一定致病力。以2、4 龄幼虫为例,幼虫死亡时变为褐色或黑红色,虫体软化,保湿培养2~3 d,体表覆盖白色菌丝和黑色孢子囊(图5)。表3 所示,接菌处理后茴香薄翅野螟2~5 龄幼虫累计死亡率随龄期的增长而下降,侵染第3~5天各龄幼虫累计死亡率存在极显著差异,但在第7~9天3龄幼虫与4龄幼虫的累计死亡率无显著差异;低龄幼虫比高龄幼虫对菌株更敏感,以1×107孢子/mL 孢子液处理9 d 后2龄幼虫的累计死亡率为71.78%,而5 龄幼虫的仅为28.89%。

图5 茴香薄翅野螟幼虫侵染症状Fig.5 Infection symptoms of Evergestis extimalis larvae

表3 冻土毛霉处理后不同时间茴香薄翅野螟幼虫累计死亡率Tab.3 Cumulative mortality of Evergestis extimalis larvae at different times after treatment with Mucor hiemalis
由图6可知,孢子悬浮液接种茴香薄翅野螟2~5龄幼虫后,随着时间的推移,累计死亡率都呈现逐渐上升的趋势。2 龄幼虫接菌1 d 后出现死亡,3~5 d 死亡率增长较快,随后渐趋平稳,接菌9 d 后全部死亡,累计校正死亡率达66.7%;侵染3 龄幼虫1 d 后出现感菌死亡,1~5 d 增长较快,10 d 后全部死亡,累计校正死亡率为47.50%;接种4 龄幼虫2 d 后初见感染,11 d 后供试幼虫全部死亡,累计校正死亡率为41.28%;5 龄幼虫处理3 d 后感菌且在1~3 d 对照累计死亡率高于处理组,11 d 后幼虫全部死亡,累计校正死亡率仅28.50%。
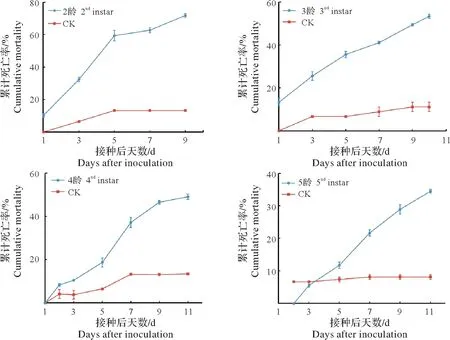
图6 冻土毛霉1×107 个/mL的孢子浓度对茴香薄翅野螟幼虫的致病力Fig.6 The virulence of Mucor hiemalis at 1×107 spores/mL concentration on Evergestis extimalis larvae
2.5 不同孢子浓度对茴香薄翅野螟2龄幼虫的毒力
随着经冻土毛霉侵染时间的延长,冻土毛霉对茴香薄翅野螟2龄幼虫的LC50减小,在处理后第5天,对茴香薄翅野螟2龄幼虫的LC50最大,为1.38×106孢子/mL,当侵染第8天后,对茴香薄翅野螟2龄幼虫的LC50最小,为1.02×105孢子/mL(表4);冻土毛霉对茴香薄翅野螟2 龄幼虫的LT50随着接种浓度的增加而缩短,当冻土毛霉孢子浓度为108孢子/mL 时,对茴香薄翅野螟2 龄幼虫的LT50最短,为3.73 d,而当冻土毛霉孢子浓度为105孢子/mL时,茴香薄翅野螟2龄幼虫的LT50最大,为6.74 d,见表5。

表4 冻土毛霉对茴香薄翅野螟2龄幼虫的LC50Tab.4 LC50 of Mucor hiemalis against Evergestis extimalis second instar larvae
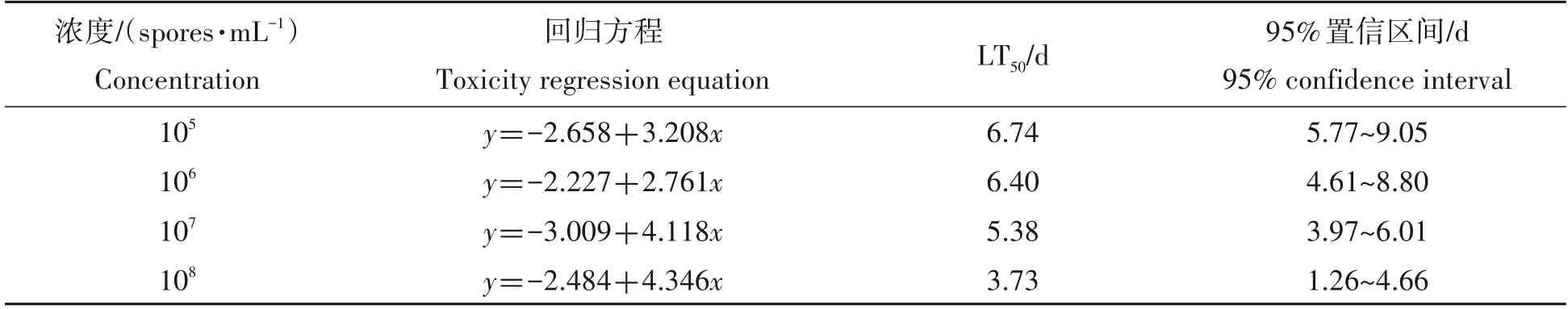
表5 冻土毛霉对茴香薄翅野螟2龄幼虫的LT50Tab.5 LT50 of Mucor hiemalis against Evergestis extimalis second instar larvae
3 讨论与结论
昆虫病原真菌资源丰富,生长繁殖快易生产,能寄生多种昆虫,且害虫很难产生抗性,因此在农林害虫防治上具有相当大的优势[23]。生防菌株资源的发现及鉴定是利用昆虫病原真菌进行害虫生物防治的基础[24-25]。相较其他昆虫,茴香薄翅野螟的生物防治资源较为匮乏。本研究从罹病的茴香薄翅野螟幼虫体上分离得到了一株病原真菌,结合形态学及分子生物学鉴定为冻土毛霉(M.hiemalis),通过柯赫氏法则验证其致病性,结果表明菌株对茴香薄翅野螟低龄幼虫有较强致病力,对高龄幼虫也有一定防治效果。研究病原真菌的培养条件对提高其利用率具有重要意义。赵鹏飞等[26]从6 种培养基中筛选出寄生曲霉Q527(Aspergillus nomiusQ527)生长最优培养基为PDA 培养基;刘思雨等[27]研究发现,SDAY 培养基是金龟子绿僵菌MAXD170705 生长及产孢的最适培养基,本试验表明在此培养基上冻土毛霉的表现相比其他培养基更好。近年来,许多研究表明昆虫病原真菌对多种鳞翅目害虫有较强的防治效果[28-31]。郑亚强等[32]经分离培养获得莱氏绿僵菌(Metarhizium rileyiZYSP190701),用1×108孢子/mL 浓度接种草地贪夜蛾3龄幼虫7 d后,幼虫的感染率达100%;何劲等[33]分离获得4株虫生真菌并测定菌株对小菜蛾的毒力,试验表明4株菌对小菜蛾都有一定毒力且致死率随时间的延长而提高,本研究结果与之相似;有研究者[34]在测定莱氏野村菌(Nomuraea rileyi)对斜纹夜蛾(Spodoptera litura)幼虫的致病力时发现菌株对2 龄幼虫的侵染率最高,本试验结果与其一致。
冻土毛霉QH01 在室内虽对茴香薄翅野螟2~5 幼虫均有致病力,但对高龄幼虫的侵染效果不佳,因此选用2 龄幼虫测定菌株对其的毒力。据报道,金龟子绿僵菌CHMA-005 对茶尺蠖(Ectropis oblique)幼虫的LC50随侵染时间的减少,呈现梯度上升趋势[35];雷妍圆等[36]试验表明莱氏绿僵菌GZSF-1对草地贪夜蛾2 龄幼虫第7 天的LC50是1.02×106孢子/mL,随孢子悬浮液浓度的升高,幼虫的LT50缩短,当浓度为1×109孢子/mL 时,幼虫的LT50为3.03 d。本研究也发现随着经冻土毛霉侵染时间的延长,冻土毛霉对茴香薄翅野螟2 龄幼虫的LC50递减,在侵染第8 天后,对茴香薄翅野螟2 龄幼虫的LC50最小,为1.02×105孢子/mL,接种浓度为108孢子/mL 时,对茴香薄翅野螟2 龄幼虫的LT50为3.73 d。综上,冻土毛霉QH01 有很好的生防潜能,值得发掘利用。
毛霉菌多为腐生,广泛应用于发酵工程[37],少数可致人和动物发病[38]。但毛霉菌对害虫有防治效果的相关报道很少。陈健鑫等[39]在云南香格里拉叉襀科稚虫虫体上得到一株冻土毛霉,该菌在稚虫处于亚健康状态及抵抗力降低时易感,会加速寄生过程使稚虫死亡;本试验表明冻土毛霉QH01 对健康的茴香薄翅野螟低龄幼虫有较高的致死率,用1×107孢子/mL 孢子液处理9 d 后2 龄幼虫的累计死亡率达71.78%,但对高龄幼虫的致死率较低,这种显著差异可能归因于不同的体壁结构和免疫能力,鳞翅目害虫的低龄幼虫体壁比高龄幼虫体壁较薄,致病菌更易侵染,高龄幼虫的角质层硬化增厚、免疫及代谢加强因而抗病性更强[40-41];以108孢子/mL 冻土毛霉孢子液接种2 龄幼虫的LT50最短,为3.73 d,因此利用该菌株防治茴香薄翅野螟幼虫的适宜期是低龄幼虫高峰期。王燕[42]从患病韭菜迟眼蕈蚊(Bradysia odoriphaga)幼虫中分离到一株冻土毛霉菌,经试验发现该菌对韭菜迟眼蕈蚊不同虫态的致病力存在显著差异,对韭菜迟眼蕈蚊幼虫有较强致病力,而对卵及蛹的致病力相对较弱;本试验仅测定了菌株QH01对茴香薄翅野螟幼虫的致病力,对其他虫态的致病性需进一步研究。许多昆虫病原真菌在室内条件下具有良好的致病性,但其田间效果取决于有利的环境条件,如温度、湿度和微生物特性等[43-44]。研究[45]表明18~28 ℃是冻土毛霉BO-1 生长和产孢的最适温度,经生物测定发现在此温度下该菌对韭菜迟眼蕈蚊幼虫的致病性高于其他温度。但冻土毛霉QH01的生物学特性、侵染机制等尚不清楚,仍需进一步探究。
本研究首次报道了冻土毛霉对茴香薄翅野螟幼虫的致病性,丰富了该虫的生防资源,为后续菌株对茴香薄翅野螟致病机理的深入研究奠定基础,也为该虫生防制剂的研发、绿色防控和农产品质量安全问题提供了理论依据。
猜你喜欢
金沙江文艺(2021年12期)2021-12-22
今日农业(2021年21期)2021-11-26
中老年保健(2021年8期)2021-08-24
莫愁·智慧女性(2021年8期)2021-08-16
小哥白尼(趣味科学)(2020年7期)2020-05-22
小哥白尼(趣味科学)(2018年9期)2018-12-18
微循环学杂志(2017年3期)2017-08-30
中国调味品(2017年2期)2017-03-20
湖北农业科学(2014年18期)2014-11-20
时代英语·高三(2014年5期)2014-08-26
