
植物miR396-GRF模块的生物学功能及其潜在应用价值
2023-01-05 08:45山琦贾惠舒姚文博刘伟灿李海燕
生物技术通报 2022年10期
山琦 贾惠舒 姚文博 刘伟灿 李海燕
(1.吉林农业大学生命科学学院,长春 130118;2.海南大学热带作物学院,海口 570228)
植物miRNA(microRNA)是一类具有调控作用的小分子RNA,其本身并不编码功能蛋白,而是通过负调控靶mRNA行使功能。miR396是植物中保守存在的miRNA家族之一,并且植物中保守的生长调节因子(GRF)基因家族的多数成员已被证实是其作用的靶基因[1]。近年来的研究发现,miR396通过负调控生长调节因子GRF家族基因,可以影响植物叶片及花器官的形态建成、根和茎组织的伸展、种子的大小、数量等表型变化[2],进而影响植物的生物量及作物产量[3],并可改善植物吸收氮和磷肥的能力及植物耐受逆境胁迫的响应能力[4-5]。此外,最新的研究结果证明,GRF基因在基因工程技术的遗传转化过程中,不仅可以显著提高外植体的再生效率,还可拓展受体植物的基因型[6]。因此,miR396介导的GRF调控(miR396-GRF)模块在改良植物生物量、作物产量、抗逆能力及提高植物遗传转化效率方面,已展现出了潜在的应用价值。本文介绍了miR396-GRF模块的作用模式,综述了近年来miR396-GRF模块的生物学功能及分子机制研究进展。为进一步深入研究miR396-GRF模块及其开发应用提供思路和参考。
1 植物中的miR396与GRF基因及其互作模式
目前,在miRNA的序列数据库(miRBase)中,共收录了植物47个物种中的138个前体miR396(pre-miR396)和175个成熟体miR396(mat-miR396)序列(包括132个miR396-5p和43个miR396-3p序列)。序列比对分析发现,不同物种中的mat-miR396序列具有高度的同源性(图1)。并且,大多数matmiR396序列与拟南芥中的ath-miR396a-5p和athmiR396b-5p两种成熟序列形式相同,其二者在序列长度上一致,但末位具有一个不同的核苷酸碱基(图1)。并且,在目前已有的报道中,这两种matmiR396序列也已被证实相比miR396家族的其它成员,具有更显著的表征作用。

图1 不同植物物种中miR396基因家族成熟区序列比对分析Fig.1 Alignment analysis of mature region sequences of miR396 gene family in different plant species
生长调节因子GRF是一类保守存在于植物中的转录因子,在调控植物生长发育等过程中起重要作用。GRF蛋白的N端普遍含有QLQ和WRC两个保守功能结构域(图2)。在QLQ结构域中,除保守的Q/Gln-L/Leu-Q/GLn氨基酸外,还含有疏水性的酸性氨基酸残基 F/Phe、Y/Tyr、L/Leu、E/Glu和P/Pro等,或其他理化性质类似的氨基酸(图2-A)。这些氨基酸残基可能在GRF蛋白与靶蛋白互作中起关键作用。在WRC结构域中,含有保守的W/Trp-R/Arg-C/Cys氨基酸残基,以及由3个C/Cys和一个H/His组成的C3H motif,这些高度保守的氨基酸位点可能在决定GRF蛋白核定位和结合下游基因顺式作用元件过程中起重要作用(图2-B)[7]。值得注意的是,紧随C3H motif区域之后,有一个保守的“RSRK-VE区”(图2-B),该区域对应的核苷酸编码序列大多高度保守,且与成熟的miR396序列近乎互补。目前已证实,miR396在此保守位点的“CGUUCAAGAA”和“AGCNUGUGGAA”之间剪切GRF基因的mRNA序列(图3),进而负调控GRF基因的表达[1]。但是,也有少数GRF成员的“RSRK-VE”区对应的核苷酸序列不能与成熟miR396序列互补配对,因此不被miR396剪切调控。例如,在拟南芥9个AtGRF家族成员中,AtGRF5和AtGRF6虽含有保守的“RSRKVE”,但其核苷酸编码序列却不能与成熟miR396序列互补,因此不受miR396调控[8]。此外,植物中GRF蛋白家族的C端氨基酸序列高度可变,推断其可能与GRF蛋白功能的多样性或转录激活有关[9]。
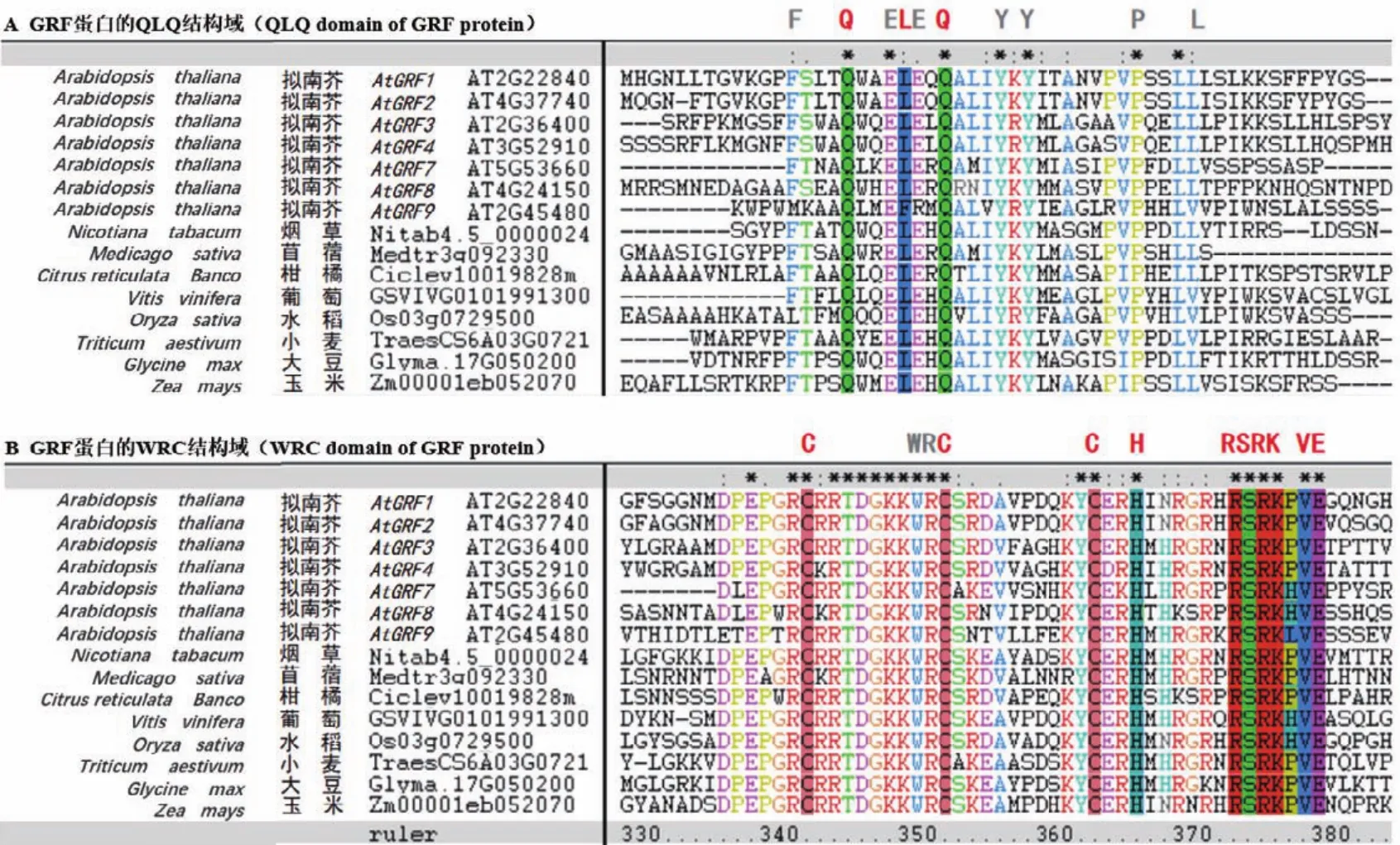
图2 不同植物物种中GRF蛋白家族氨基酸序列的保守结构域比对分析Fig.2 Alignment analysis of the conserved domains of GRF protein family amino acid sequences in different plant species
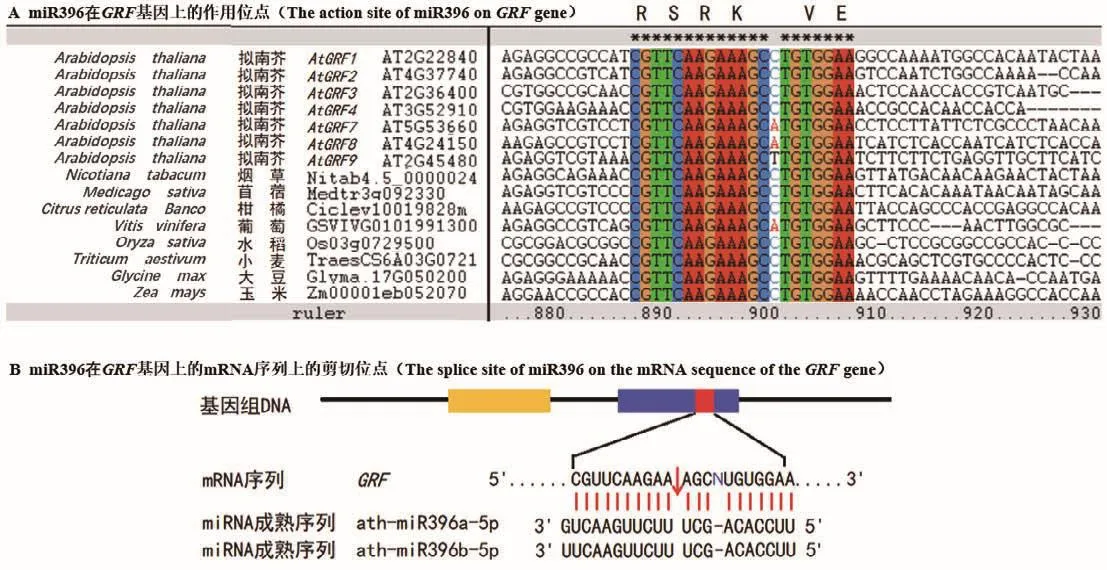
图3 不同植物物种中miR396在GRF基因序列上的作用位点及剪切位点Fig.3 Action sites and splicing sites of miR396 on GRF gene sequences in different plant species
2 miR396-GRF模块的生物学功能及应用研究进展
2.1 miR396-GRF模块对植物生长发育及生物量的影响
自miR396-GRF模块被发现以来,越来越多的研究证实了miR396-GRF模块具有影响植物组织器官形态建成过程的调控功能。当突变体中的miR396基因被插入失活,或是miR396基因被敲除表达,以及在超表达GRF基因的植物材料中,miR396-GRF模块通常表现出促进植物组织生长发育过程的趋势。例如,在拟南芥中过表达内源的AtGRF3基因[10]、异源的烟草 NtGRF8[11]、白菜 RbGRF8[12],或是在水稻中过表达rOsGRF1基因(氨基酸同义突变,但核酸序列上消除了miR396靶位点)等[13],都促使转基因植株变高,叶片变大,根系粗壮,种子及花器官发育延缓等形态变化[14-16]。并且,同义突变处理后的rGRF基因促进植物增长的效果更加显著[10]。相反,当在植物中超表达miR396基因,或是突变体中的GRF基因被插入失活,以及GRF基因在植物材料中被抑制表达,miR396-GRF模块通常会表现出抑制植物组织生长发育过程的趋势。例如,在拟南芥[17]和水稻[18]等植物中过表达同源或异源miR396的转基因株系,都表现出生长迟缓,植株矮小,根系变短,叶片也变得窄小的形态变化,这些表型与grf突变体相似[19]。因此,miR396与GRF基因在调控植物组织器官生长发育的过程中呈负相关关系,二者之间的靶向作用平衡制约着植物的形态表征。
此外,miR396-GRF模块在一些经济类植物中的表达调控,也表现出了一些有益的表型变化。例如,Wang等[20]在我国木材主要来源的杨树(Populus alba × P.glandulosa)中,证实了PagGRF12a基因的表达调控作用可增加杨树木质部的宽度,促进木材的建成。Liu等[21]在可用于生物燃料生产的植物柳枝稷(Panicum virgatum L.)中,证实了rPVGRF9基因的过表达可改变其生物量及原料质量。此外,在生菜(Lactuca sativa)中过表达LsaGRF5基因后,生菜叶片明显变大,可显著提升蔬菜的生物量产出[22]。以及在可用作牧饲材料的苜蓿[23]中,利用Target mimicry技术敲降表达miR396后的MIM396苜蓿材料,叶片的生物量也显著增加,并且根系粗壮发达。综上所述,miR396-GRF模块的调控作用在提升植物生物量方面展现出了潜在的应用价值。
2.2 miR396-GRF模块对产量等农艺性状的影响
基于对植物生长发育过程的影响作用,miR396-GRF模块也展现出了对作物产量等农艺性状的影响能力。例如,miR396沉默体 MIM396水稻[14]、mir396ef突变体水稻[24]和GRF基因过表达后的水稻和小麦植株[25-27],都表现出了穗数的显著增加,以及粒长和粒宽的增长现象,并且它们相对野生型能获得更高的粮食产量。在果蔬植物番茄中,miR396通过调控不同的GRFs靶基因,也影响着番茄果实的多方面变化,过表达rGRF4和rGRF5的转基因番茄可通过增加果皮厚度和果实心室数使果实重量明显增加[28]。在油菜中,本源表达BnGRF2基因后的转基因油菜的种子千粒重和含油量也得到显著提升[29]。因此,miR396-GRF模块的调控作用在提升作物产量及品质方面也展现出了潜在的应用价值。
另外,目前的研究报道显示,miR396-GRF模块还具有提升植物氮肥和磷肥利用率的能力。在缺氮条件下,过表达ZmGRF1基因的玉米可正向调控玉米氮素相关途径基因的表达,促进氮肥吸收[30]。OsGRF4蛋白在水稻中的高水平积累也可以提高光合作用和氮肥利用效率[5]。此外,在高供磷条件下,过表达SlGRF9的转基因番茄对土壤中磷的利用率也显著提高。所以,miR396-GRF模块在植物调控肥料吸收能力方面也有益于农艺生产。
2.3 miR396-GRF模块在逆境胁迫响应过程中的调控作用
在农业生产中,植物体对逆境环境的适应能力也是影响田间生产的重要因素。近年来的研究表明,miR396-GRF模块在调控植物响应逆境胁迫的过程中发挥着积极的调控作用。例如,MIM396番茄植株中由于miR396的下调表达,导致其在缺水胁迫下的蒸腾速率(E)降低,水分利用效率(WUE)显著提高[31]。并且,在番茄中过表达AtGRF9基因,可提升根系蔗糖和干物质的含量,增强根系分泌质子的能力。所以,miR396-GRF模块的调节作用对于番茄响应干旱等多种逆境胁迫过程都具有积极的意义[32]。在草坪草匍匐剪股颖(Agrostis stolonifera)中,超表达Osa-miR396c的转基因植株在高盐度环境中保水性增强、叶绿素含量相对较高、且细胞膜具有较好的完整性和钠离子排除能力,由此表明其具有较强的盐胁迫适应能力[33]。此外,OsGRF4基因突变体水稻在苗期的耐寒性增强,冷胁迫处理后幼苗的存活率显著升高[3]。拟南芥经UV-B辐射后,miR396抑制AtGRF1/2/3表达,降低细胞增殖速率的同时抑制叶片生长,起到耐紫外线的作用。因此,miR396-GRF模块在调控植物响应多种非生物胁迫的过程中具有积极意义。
在植物响应生物胁迫的过程中,miR396-GRF模块也同样具有调控功能。在大豆中,通过过表达或敲除实验干扰miR396-GmGRF调控途径,会抑制大豆孢囊线虫幼体向成体的发育[34]。此外,稻瘟病毒严重影响着水稻的生长和产出。研究发现,OsamiR396d抑制OsGRF6/7/8/9表达,延缓水稻生长发育,减少病毒向新生组织扩散,从而提高了抵抗稻瘟病的能力[35]。因此,miR396-GRF模块在调控植物响应生物胁迫的过程中也展现出了积极作用。
2.4 GRF提高植物再生效率的研究
转基因受体材料遗传转化效率低及基因型的限制(即小麦等物种中仅有少数基因型的品种材料才可以被成功遗传转化)是基因功能研究及基于基因组编辑育种技术的主要瓶颈。2020年Kong等[6]研究发现,将AtGRF5及其同源基因导入甜菜、油菜、大豆和向日葵中均可有效提高转化效率并产生完全可育的转基因植株。紧随其后,Pan等[36]进一步证实了在AtGRF5基因的辅助转化作用下,西瓜外植体材料的转化效率提高了40倍。同期,Debernardi等[37]研究发现小麦TaGRF4-TaGIF1嵌合体的转基因表达显著提高了4个品种小麦的再生效率,并且外植体再生过程周期短,不依赖激素调节,且转化体没有负面表型影响,从而拓展了小麦被转化受体材料的基因型。此后,高彩霞团队通过共转化瞬时表达GRF-GIF嵌合体蛋白也证实了其有效性,并且小麦TaGRF4-TaGIF1嵌合体与基因编辑体系共表达也被证实可以高效地获得编辑成功的突变体材料[38]。目前已被证实,GRF基因及其嵌合体相比以往使用的提高遗传转化效率的生长调节基因如BBM、WUS、IPT等更加有效[39]。显然,GRF基因在改善双子叶和单子叶植物外植体再生效率,及提高基因编辑效率方面都展现出了诱人的应用前景。
3 植物miR396-GRF模块作用的分子机制研究
3.1 GIF作为辅激活因子可增强GRF蛋白的活性及功能
目前,在多篇研究报道中,都证实了不同物种中的 GRF 和 GIF 之间存在着蛋白互作关系[7,20,40],GIF蛋白也由此而得名为GRF蛋白相互作用因子(GRF-interacting factor,GIF)。Lu 等[13]研究发现,过表达OsGIF1基因的水稻没有显著的表型变化,而OsGIF1基因敲除后的水稻表现为植株矮小等表型形态,与OsGRF1基因敲除后的水稻形态相似。并且,mOsGRF1基因过表达和OsGIF1基因敲除的杂交系水稻植株也呈现矮小状态。因此推断,GIF蛋白虽然不能独立行使功能,但确是生长调节因子GRF调控植物生长发育过程中不可或缺的因素。
进一步的深入研究表明,GRF和GIF蛋白首先在细胞核内形成功能复合物,然后GIF蛋白负责招募SWI/SNF染色质重塑复合物,借助染色质重塑酶BRM,利用ATP水解产生的能量,破坏组蛋白与DNA的相互作用,使DNA和组蛋白之间变得松散,以便作为转录因子的GRF蛋白与顺势作用元件结合并激活或抑制下游目的基因的转录[9,39],进而发挥各种生物学功能。因此,GIF作为GRF蛋白的辅激活因子,具有增强GRF蛋白活性的功能。
此外,植物细胞的去分化和再生能力,直接影响着植物外植体材料的遗传转化效率。一般来说,分化细胞具有紧密的染色质状态,而去分化过程使染色质变为开放松弛状态[39]。由于GRF-GIF复合体可以使染色质重塑而变得松散,因此可以有助于提升植物外植体材料的再生能力和遗传转化效率。
3.2 miR396-GRF模块调控细胞增殖的机制研究
目前已有的研究结果显示,在植物成熟的叶片、根和茎等组织中,miR396的表达量相对较高,GRF基因的表达量相对较低;而在根尖、花芽和幼嫩的叶片等初生组织中,miR396的表达量相对较低,GRF基因的表达量却相对较高[41]。
其中,KNOX蛋白是促进茎尖组织分生的关键因子。Kuijt等[42]证实,水稻OsGRF3和OsGRF10基因通过与KNOX家族基因启动子区的相互结合,从而抑制OsKN2基因的表达,并影响植物分生组织的发育。
另外,在植物中过表达miR396或敲除GRF基因可观测到植株矮小,器官发育延缓等现象,但这种变化可以被GRF基因的表达所恢复[14-16]。细胞水平的检测结果证实,上述转基因材料叶片单位面积内的表皮细胞数量显著减少,细胞分裂增殖受到了抑制[1,19]。转录组测序和荧光定量PCR分析结果显示,miR396-GRF模块的遗传转化材料中,一些与DNA 复制、细胞分裂和细胞周期调节等有关的基因具有显著表达差异,如MAPK、CYC家族基因等[29,33]。因此说明,miR396-GRF 模块通过控制植物组织细胞的分裂增殖过程[19,33],进而影响着植物组织的生长发育过程。
3.3 miR396-GRF模块的激素调节机制研究
植物激素是植物细胞接受环境信号诱导产生的活性物质。在细胞分裂分化、开花结实、成熟衰老等多个方面,植物激素对植物的生长发育过程起着重要的调控作用[43-44]。已有的研究报道表明,miR396-GRF模块调控植物生长发育的过程与赤霉素(GA)、吲哚乙酸(IAA)等激素调节及合成过程有着密切关系(图4)。Zhang等[24]对水稻双突变体osmir396ef的叶片进行了液相质谱色谱联用及代谢组分析,发现GA3,GA4,GA7水平以及GA合成前体甲羟戊酸(mevalonic acid,MVA)水平均明显提高,由此说明,miR396-GRF模块间接影响着IAA的生物合成。此外,郭泾磊等[45]分析发现,在小麦TaGRF基因的启动子区存在大量IAA的响应元件,在外源施加IAA处理后,可显著上调TaGRF基因的表达,进而影响小麦分蘖。此外,在水稻和拟南芥中,都证实了AtGRF基因的下调表达,会进一步调控IAA生物合成途径的关键基因,如YUC和ARF基因等,进而影响IAA的生物合成。因此,GRF基因的表达受到了IAA的诱导调控,GRF基因的表达又影响IAA的生物合成。综上,miR396-GRF模块参与了植物激素的调节过程,但大多缺少直接联系的作用证据,还需进一步开展深入的研究。
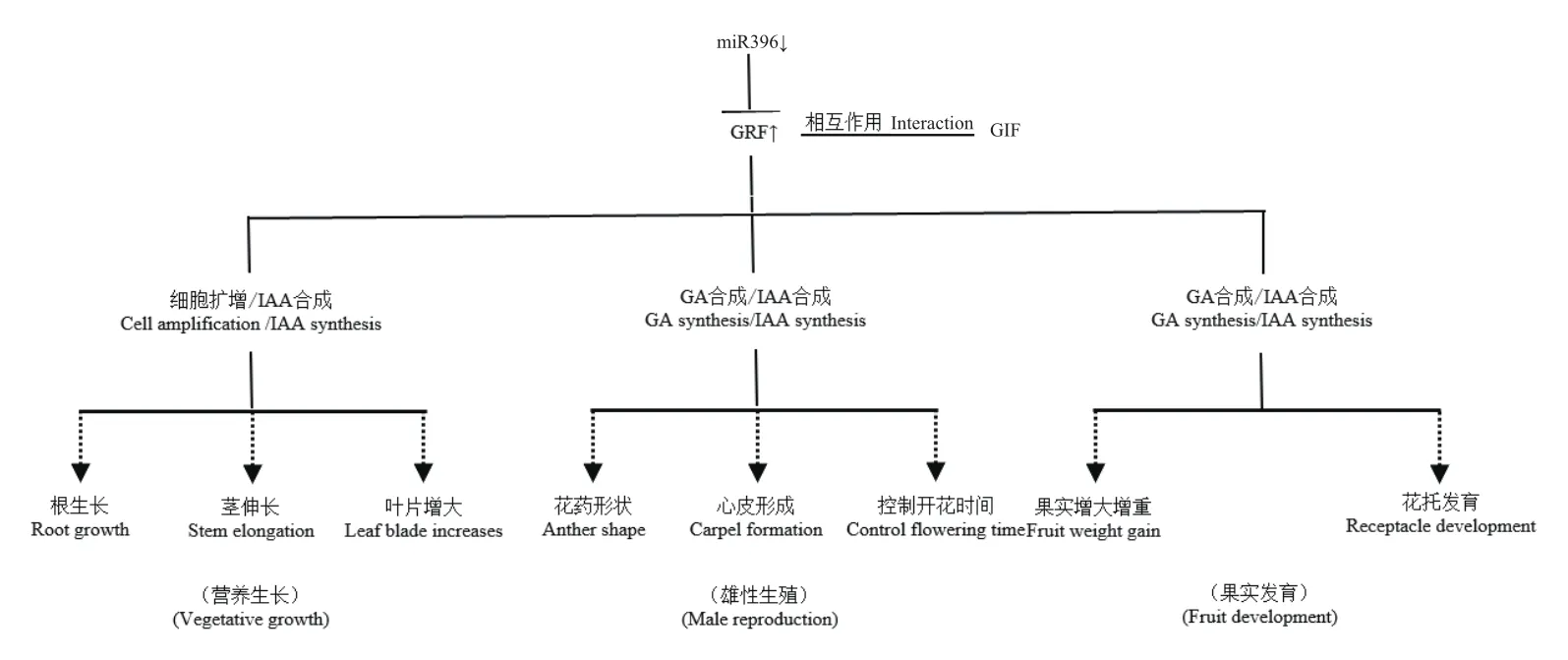
图4 miR396-GRF模块调控植物生长发育的过程Fig.4 Process of miR396-GRF module regulating plant growth and development
3.4 miR396-GRF模块调控植物逆境胁迫响应的机制研究
植物由于其固着的生活方式而不得不连续暴露于可能存在各种逆境胁迫的环境之中。为应对环境胁迫,植物进化出了各种复杂的防御机制,包括信号传感和转导途径、调控下游胁迫反应相关功能基因的表达等,从而通过气孔关闭、细胞生长、光合作用抑制、可溶性物质积累等各种生物过程,保护植物免受损害并增加生存机会。有关miR396-GRF模块遗传转化材料的转录组及Go富集分析发现,除了与细胞分裂有关的基因外,还存在很多与表达调控、激素合成、抗氧化还原反应等各种与逆境胁迫响应有关基因的差异表达[33]。因此说明,miR396-GRF模块在植物响应逆境胁迫的过程中,也发挥着重要的调控作用。
根据目前研究获得的数据表明,miR396-GRF模块在植物中响应逆境胁迫的潜在信号通路大致为:在逆境胁迫下,miR396-GRF模块被诱导转录后,首先控制蛋白激酶和转录因子等调节蛋白;然后这些调节蛋白激活或抑制下游逆境响应相关的功能基因,进而响应逆境胁迫过程(图5)。例如,Yuan等[33]通过RT-PCR分析发现,在miR396的转基因匍匐翦股颖中,WRKY、MADS和AP2等调节作用基因被下调表达,进而下游Na+逆转运蛋白AsSOS1的作用被激活,从而影响了植株的耐盐胁迫响应能力。Vall等[46]研究发现,GRFs的下调表达将导致控制活性氧(ROS)产生的乙烯反应因子ERF1基因的上调,从而阻断乙烯反应,降低抗氧化酶活性。Kim等[47]的研究揭示,拟南芥AtGRF7基因可直接作用到与脱水反应有关的DREB2A基因的启动子区,并抑制其表达,从而导致atgrf7突变体在高盐与干旱条件下的耐受性增强。Fracasso等[31]研究结果显示,MIM396番茄植株中,GRF基因的上调表达可刺激表皮细胞增殖,导致气孔密度增加,气孔变小;并可促进茉莉酸(JA)和γ-氨基丁酸(4-aminobutyric acid,GABA)的积累,从而调节气孔关闭和维系光合作用过程,改变植物的逆境适应能力。总之,关于miR396-GRF模块的研究有很多报道,但是其在植物中响应胁迫的调控网络在很大程度上仍然是未知的,也需进一步开展深入的研究工作。
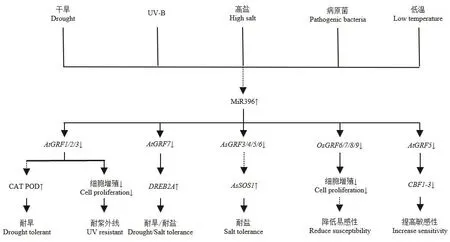
图5 miR396-GRF调控模块在胁迫响应过程中的可能调控途径Fig.5 Feasible regulation approach of miR396-GRF module in the process of stress response
4 结语
目前已有的研究结果表明,miR396-GRF模块不仅可以调控水稻及小麦等的株高、分蘖、穗数、籽粒大小及灌浆过程而影响作物产量[25-27]、提高油菜籽的出油率[29];还可改善番茄的抗旱能力[31],草坪草的耐盐能力[33]及水稻的耐寒能力[3]等,并影响大豆感染孢囊线虫[34]及水稻感染稻瘟病毒的过接证据。第四,miR396-GRF模块在影响植物生长发育方面的研究报道进展较快,但是关于其影响植物逆境胁迫响应的研究相对薄弱。第五,miR396-GRF模块调控作用的分子机制还很不清楚,缺乏调控途径之间的直接关联证据。第六,在大豆、玉米等重要物种中及同一物种中的不同基因型品种中,GRF基因可提升遗传转化效率的有效性正亟待实验证实,与基因编辑模块的联合应用更有待开发。
综上,miR396-GRF作为植物分子育种领域的候选基因,进行实际应用之前还需要很多的研究准备工作。建议加强miR396-GRF模块在标的物种中的全方面调控作用研究,深入阐明其调控作用的分子机制;并大力开发高效有益的miR396-GRF基因用于植物品种改良,快速推进GRF基因在遗传转化和基因编辑系统中的有效运用。在此,本文仅为进一步深入研究miR396-GRF途径及其开发应用整理思路和提供参考。程[35]。此外,GRF基因被证实能有效提升小麦等植物基因转化及外植体再生效率[39]。因此,miR396-GRF模块在提高作物产量、改良植物抗逆能力及提升植物遗传转化效率方面都展现出了诱人的应用前景。
然而,近年来miR396-GRF模块虽然得到了研究者们的更多关注,有关miR396-GRF模块生物学功能的研究报道也层出不穷,但在其研究发展的道路上和真正被开发应用之前,还存在很多值得注意的地方和需要解决的问题。例如,第一,miR396-GRF模块调控途径在植物界的不同物种中保守存在,在每个植物物种中,miR396和GRF家族基因的成员数都普遍较多,存在功能冗余情况。第二,miR396-GRF模块中的同一基因在不同物种中,或同一物种的不同家族基因成员之间,可能存在着显著的功能差异,甚至相反的影响。例如,TaGRF4 与ZmGRF10因缺少C端转录激活结构域,由此它们都抑制叶片的伸长生长[45,48];miR396转基因烟草抗旱能力增强[49],而miR396转基因拟南芥抗旱能力降低[50],这可能与物种之间的差异有关。第三,目前miR396-GRF模块基因的功能更多的是在模式植物中异源表达被证实,大多缺乏在标的物种中的直
猜你喜欢
做人与处世(2022年6期)2022-05-26
理财周刊(2022年4期)2022-04-30
疯狂英语·新阅版(2022年5期)2022-04-29
昆明医科大学学报(2022年1期)2022-02-28
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
红领巾·萌芽(2017年5期)2017-06-23
爆笑show(2016年7期)2017-02-09
少儿科学周刊·儿童版(2015年10期)2015-11-07
少儿科学周刊·儿童版(2015年1期)2015-07-07
