
小自噬的研究进展*
2012-11-13 07:57陈艾,童煜,毛萌△
中国病理生理杂志 2012年5期
陈 艾, 童 煜, 毛 萌△
(1四川大学华西第二医院儿科,四川成都610041;2泸州医学院附属医院儿科,四川泸州646000)
自噬(autophagy)是继坏死和凋亡后发现的第3种细胞死亡形式,与疾病的发生息息相关,是目前细胞领域研究的热点之一。自噬是指细胞在自噬相关基因(autophagy-related gene,Atg)的调控下利用溶酶体/液泡降解自身受损的细胞器和大分子物质的过程,在进化过程中有高度的保守性。本文就目前小自噬(microautophagy)的研究近况作一综述。
1 自噬的背景
真菌中与哺乳动物细胞的溶酶体相当的细胞器称为液泡(vacuole)。根据底物进入溶酶体/液泡途径的不同可将自噬分为3种类型[1-2]:大自噬(macroautophagy)、小自噬和分子伴侣介导的自噬(chaperone-mediated autophagy,CMA)。自噬的主要功能是保证细胞对底物的再利用,维持细胞的相对稳态[3]。
大自噬即通常所指的自噬,是最主要的自噬形式,其特点是自噬体(autophagosome)的形成[4]。首先,来源不明的游离双层膜结构包裹胞液或受损的细胞器,形成自噬体。然后,溶酶体/液泡与自噬体融合形成自噬溶酶体,将其内容物降解成氨基酸、游离脂肪酸等为细胞重建提供原料,并在机体处于饥饿、营养缺乏等条件下提供能量。
小自噬现象最早由de Duve和Wattiaux在1966年发现,而其概念直到1983年才被明确[5]。酵母中的小自噬研究比较广泛和深入,而哺乳动物细胞的小自噬研究较少。一般认为,小自噬分为非选择性小自噬(nonselective microautophagy,NSM)和选择性小自噬(selective microautophagy,SM)[6],前者是指液泡非特异性吞噬蛋白小微粒或细胞器成分,表现为液泡膜受底物触发后发生管状内陷,底物从内陷处被吸入形成小泡[7]。SM的吞噬对象是特定的,如线粒体、细胞核、过氧化物酶体等,表现为哺乳动物的溶酶体膜或植物/真菌等生物的液泡膜与被吞噬底物直接融合,溶酶体膜/液泡膜先向外出现臂膀一样的突起,将特异性底物揽入其中,再逐渐形成多个内陷的小泡,之后小泡完整从膜上的自噬管顶端脱落至膜内被降解[3]。小自噬的特点是膜直接形变包裹底物[8]。
CMA为胞浆内蛋白底物结合到分子伴侣后再被转运到溶酶体腔中,分子伴侣主要为热休克同源蛋白70(70 kD heat- shock cognate protein,Hsc70)。Hsc70可识别5个氨基酸(赖-苯丙-谷-精-谷酰胺)固定排列的5肽序列(Lys-Phe-Glu-Arg-Gln,KFERQ),并可与含有此序列的蛋白质发生相互作用。KFERQ是可溶性蛋白,Hsc70与之结合后,再结合至溶酶体膜上的溶酶体相关膜蛋白2(lysosome-associated membrane protein 2,LAMP -2),转运到溶酶体内降解[9]。
然而最新研究表明,Hsc70-KFERQ-LAMP-2开启的通路只是分子伴侣介导的一个类型。Hsc70也可以选择性地与转运所需的内体(蛋白质)分选复合体(endosomal sorting complex required for transport,ESCRT)结合,开启一条类似小自噬的途径,这条途径被命名为内体小自噬(endosomal microautophagy,eMI)[10]。
2 小自噬概述
2.1 小自噬的相关基因及信号通路 Atg在各类自噬中发挥作用,不再累述。小自噬的特异基因为“Apg/Aut”通路以及两类Atg7依赖的泛素样结合系统(ubiquitin-like conjugation,Ublc)参与了饥饿诱发的非选择性小自噬或者葡萄糖诱发的选择性小自噬[11]。在第1个Ublc系统中,Atg8在E1样酶Atg7和E2样酶Atg3作用下,经过半胱氨酸水解酶的酯化,才能与膜脂质的磷脂酰乙醇胺(phosphatidylethanolamine,PE)集合。在第2个Ublc系统,E1样酶Atg7和E2样酶Atg10吸引Atg12吸附Atg5,这个Atg12-Atg5二聚体与Atg16寡聚体化,才能促使Atg8-PE复合物形成[12]。而Atg8-PE复合物在小泡形成和融合过程中起关键作用。其它特异性基因见后。
2.2 小自噬的功能 小自噬的溶酶体/液泡直接摄取底物并且与其膜融合并内吞的过程弥补了大自噬释放自噬体时丢失的大量膜成分。因此,小自噬是大自噬必要的补充,小自噬产生的小泡(vesicles)在功能上相当于大自噬的自噬体。小自噬通过小泡内吞的膜成分正好补充大自噬以及CMA形成自噬体释放的膜成分,这种平衡维持了膜的稳态[13]。但是,这些膜成分在3种自噬之间是如何流通和转换的,目前尚不清楚。
3 小自噬的分类
3.1 eMI 最近,Sahu等[10]鉴定出一种由 ESCRT及Hsc70共同参与,将可溶性胞质蛋白转运到内体的降解过程。在该研究中,阻断大自噬过程,可溶性蛋白仍然可以进入内体;若同时敲除CMA膜受体LAMP-2A,也不影响内体摄取蛋白。通过CMA通路的底物蛋白在摄取之前是完全折叠的,必须先经解折叠才能被摄取。而实验中,溶酶体中的折叠蛋白数量没有降低,证明有一条非大自噬也非CMA的自噬通路,于是Sahu等命名这条通路为eMI。
虽然Hsc70没有任何标准区域与脂质蛋白结合,但是其C端的AA526-539中有一簇可通过静电作用与磷脂酸结合,其它两簇与疏水的氨基酸区域结合,有助于将未解折叠的可溶性脂质摄入次级内体后降解。这种结合是选择性的,有的底物不经Hsc70也能通过eMI途径被降解[14]。
eMI发生依赖于胞内多泡体(multivesicular bodies,MVBs)的形成,依赖ESCRT I-III系统的激活及其与底物交联。ESCRT-I和ESCRT-II是可溶解于细胞浆的异聚体,ESCRT-III是可溶解于细胞浆的单体,最终3种复合物均被募集到溶酶体内膜形成不能溶解的矩阵,才能形成MVBs需要的微环境[15]。
ESCRT-I的亚基包括 VPS23、VPS28和 VPS37等,这些亚基被定植于初级内体膜的VPS27募集在一起,并与膜内磷酸化磷脂酰肌醇结合,此过程中VPS23与VPS27产生交联,激发ESCRT-I与泛素系统联合,激活自噬调控[16]。ESCRT-I与初级内体结合后才能募集包含VPS22和VPS25的 ESCRT-II形成MVBs通路。MVBs通路开始形成后,已经转位在内体上的ESCRT-II再吸附胞质中的ESCRT-III,至少有6种ESCRT-III亚基存在于酵母中,而在人类至少有10种。3种ESCRT复合物相互作用,使底物聚集于内体膜,同时诱导后者向腔内弯曲出芽,最终脱芽产生胞内小囊泡,使底物通过MVBs途径进入内体被降解。内吞过程执行的后期,ESCRT-III必须从ESCRT复合物上解离以便循环利用,解离过程由VPS4催化[17]。
以上实验均提示,小自噬与CMA途径可能共用一些调节因子。在Sahu等的研究基础上,Santambrogio等[18]总结了eMI与CMA途径的主要异同是:eMI途径的底物为可溶性物质,摄入前需为折叠状,需与磷脂酰丝氨酸结合,不经LAMP-2A识别,不需GTP供能,最终与次级内体(late endosome)融合后被分解并包裹成外体(exosome),分泌至溶酶体被降解。
CMA途径的底物摄入前必须解链成单体,在LAMP-2A的引导下,消耗ATP及GTP,最终在溶酶体内被降解。其共同点为:2条途径对底物均为直接摄取,都需要KFERQ样序列及Hsc70的识别,均需消耗ATP酶,见图1、表1。
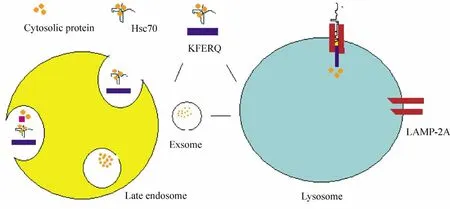
Figure 1.The differences between endosomal microautophagy(eMI)and CMA.图1 内体小自噬与分子伴侣小自噬形成的区别

表1 内体小自噬与分子伴侣小自噬的区别[18]Table 1.The differences between endosomal microautophagy(eMI)and CMA
3.2 非选择性小自噬 NSM的发生需要ATP酶以及跨液泡的质子梯度;摄取过程需要GTP酶,跨膜电位以及脂肪和膜成分的持续补充。其调控因子除了一些自噬相关蛋白外,还依赖于雷帕霉素靶蛋白(target of rapamycin,TOR)和EGO蛋白复合体(exit from rapamycin-induced growth)的结合,后者由Ego1、Gtr2 及 Ego3 组成[19]。
另一类也在NSM中发生作用,即液泡转运伴侣(vacuolar transporter chaperone,VTC)复合物。当饥饿、缺氧等应激出现时,VTC不均一的分布于液泡膜,富集于内陷的自噬微管膜上。VTC复合物包含3个C端的螺旋状结构:体积较小的VTC1p完全种植于膜内,VTC2p、VTC3p/VTC4p复合物形成一个较大的N端朝向细胞质。VTC是自噬微管的主要成分,在小泡从微管上剪切下来时发挥关键作用。如果VTC变异,自噬微管仍可以形成,但是最终形成的吞噬小泡却难以被剪切下来[20]。
3.3 酵母研究中发现的选择性小自噬
3.3.1 选择性小自噬的特点 自噬微管的形成是选择性小自噬区别于其它自噬的最大特点:(1)小自噬开始时,溶酶体/液泡膜上一些脂质从一侧向表面出芽样突起,进而在一些转膜蛋白协助下,更多的脂质或含有脂质样结构的蛋白富集,驱动并参与出芽的过程,此过程需要具有GTP酶活性的Vps1调控;(2)脂质蛋白集落在伸延的同时收缩卷曲,开始朝胞内内陷,向内弯曲形成类圆形、有一定直径的小管样结构,称为“自噬管”(autophagic tube);(3)同时,自噬管形成初期出芽处的蛋白扭结,成为识别力很强的瓶颈,正是这瓶颈样结构将自噬管区别于一般的膜内陷;(4)沿着自噬管,细胞内膜蛋白迅速松解后向管的顶端排列,这些同源性蛋白以拓扑样方式编构,在ATP功能下逐渐形成小泡,开始小自噬的选择性吞噬活动[7]。
3.3.2 过氧化物酶体的小自噬(micropexophagy)过氧化物酶体(peroxisome)为真核细胞的一种细胞器。同样,液泡朝过氧化物酶体伸出突起,这种突起被命名为液泡分离膜(vacuolar sequestering membrane,VSM),经常又被间隔成许多小隔膜[21]。在VSM与过氧化物酶体融合时,帽子形状的双层膜结构,称为过氧化物酶体小自噬特异性膜突起(micropexophagy- specific membrane apparatus,MIPA),协助VSM进行吞噬。MIPA是包裹过氧化物酶体的小泡与液泡接触时双层膜处形成的扁平状小泡,形成于被吞噬小泡的尖端,帮助形成过氧化物酶体自噬体。过氧化物酶体小自噬过程分成3个阶段:(1)液泡与过氧化物酶体接近并排,接收成熟过氧化物酶体发出的信号;(2)回应信号,液泡出现膜的内陷,凹陷处的边缘形成并伸出手臂一样的VSM结构,在MIPA协助下将过氧化物酶体拥入并内吞;(3)在吞噬以后,过氧化物酶体被解离产生氨基酸和脂肪以供细胞循环再利用。
MIPA的形成与上述过程的实施需要自噬核心基因的完全调控,以及过氧化物酶体特异性自噬基因,如 Atg11、Atg20、Atg24、Atg25、Atg26、Atg28 和Atg30 的参与[5]。
3.3.3 核的小自噬 (micronucleophagy,piecemeal microautophagy of the nucleus,PMN) PMN 于 2003年被Roberts等[22]发现,迄今也仅在酵母研究中被描述。指酵母的部分核膜和核质内陷进入液泡并降解的过程。PMN小泡的包膜有3层,最外层来自于液泡膜,内2层来源于核被膜。细胞核通过核泡连接(nucleus-vacuole junction,NVJ)与酵母的液泡接触。在氮气或二氧化碳缺乏时,或用雷帕霉素诱导后,NVJ即可产生。
NVJ是一种搭扣样结构,发生于液泡膜蛋白Vac8和最外层的核膜蛋白NVJ1之间。在营养缺乏时,NVJ1逐渐与脂肪代谢的2个酶:氧酮结合蛋白OSH1和辅酶A还原酶TSC13结合。TSC13为PMN形成所必须的因子之一。OSH1与NVJ1的结合抑制了丝氨酸与质膜的反应,从而使NVJ内陷形成,释放装满胞核物质的小泡到液泡,形成PMN过程[23]。
Dawaliby等[24]研究表明,NVJ在液泡表面形成的屏障在对蛋白开放的同时,偶尔会排斥V-ATP酶,由于PMN过程需要V-ATP酶激活,所以排斥V-ATP酶的屏障只能通过NVJ蛋白家族的成员和其它电化学成分代谢产生的电位差来释放小泡,比如OSH1以及TSC13等。所以,NVJ的形成包含2个独立的步骤:NVJ1和VAC8等富集于核泡两层膜接触融合的部位;同时电化学梯度和酶代谢促使液泡表面形成弥漫的屏障,经过如下5个步骤,最终完成PMN过程,如图2所示[25]。同样,PMN也需要核心基因的调控,如 Atg1~Atg10、Atg12~Atg16、Atg18、Atg22等。同时,免疫荧光检测发现PMN的形成需要特异的Atg蛋白,如 Atg11和Atg17。

Figure 2.Prodecure of piecemeal microautophagy of the nucleus.图2 核的小自噬形成过程[25]
3.3.4 线粒体小自噬(micromitophagy) 线粒体的吞噬主要通过小自噬途径[6]。当氧化应激等情况出现时,线粒体会出现渗透性转移,此过程中,线粒体出现去极化、膨胀等损伤,最终会被降解[26]。不同于其它类别的小自噬,经过荧光以及电镜的观察,线粒体小自噬分为2种类型:一种类型依赖于Uth1,一种部分定植于线粒体外膜的蛋白,液泡接收到线粒体的信号后慢慢与后者并列,吸引线粒体堆积在其周围,然后两者的膜融合产生连接,这种吞噬较精准,只吞噬线粒体而几乎不会包含其它胞质成分。第2种发现于敲除Uth1的野生型酵母,液泡可直接吞噬线粒体以及其周围的胞质碎片[27]。其特异调控基因为Atg32。
3.3.5 哺乳动物细胞的小自噬 Glaumann将分离的大鼠肝细胞溶酶体与Percoll颗粒一起孵化,发现Percoll颗粒被溶酶体吞噬,其过程类似酵母中的MIPA。由于溶酶体膜也出现杯子样突起去包裹Percoll颗粒,哺乳动物细胞的这类小自噬被称为溶酶体包裹化(lysosomal wrapping mechanism,LWM)。LWM发生于各种细胞,如大鼠组织细胞,经辐射后的大鼠肾脏巨噬细胞等。LWM发生时,初级或次级溶酶体会出现变形,由球形拉长,再伸出手臂一样的突起。目前认为LWM的特点是:(1)在摄取、吞噬和降解过程中需要消耗ATP;(2)LWM的发生依靠肌纤维的存在;(3)LWM发生时有蛋白合成[28]。
4 小自噬与疾病
目前已有许多研究证实,小自噬过程出现障碍与许多疾病相关。如神经退行性病变的阿尔茨海默病,以快速进行性痴呆、肌阵挛和特征性脑电图表现为特点的克-雅病(Creutzfeldt-Jakob disease),以进行性慢性小脑性共济失调为表现的格斯特曼-施特劳斯纳(Gerstmann-Straussler-Scheinker,GSS)病,膨胀的MVBs在GSS病人的组织中大量存在。多囊肾、心肌衰弱等也与小自噬有关。心肌糖原沉积病(Pompe病)也和糖原不能被液泡正常摄取有关[29]。小自噬与各类疾病的关系引起人们普遍关注,研究也正逐渐深入。
5 结语
目前,小自噬的主要功能被认为是维持细胞器的正常大小,保持膜成分在各类型自噬转换流通中量的平衡,以利于营养剥夺中细胞的存活。根据摄取底物的不同,小自噬被分为许多亚类型,但其发生过程基本一致,即膜的融合、小泡的形成、底物被溶酶体或液泡内吞降解等步骤。目前国内外的研究集中在小自噬的形态学方面,在其分子机制及基因调控,与大自噬以及分子伴侣自噬之间功能的交互等方面尚需要作进一步的研究与总结。
[1] Shpilka T,Elazar Z.Shedding light on mammalian microautophagy[J].Dev Cell,2011,20(1):1 - 2.
[2] 杜海磊,邱伟华,杨卫平.细胞自噬与肿瘤[J].中国病理生理杂志,2010,26(2):401-404.
[3] Farre JC,Krick R,Subramani S,et al.Turnover of organelles by autophagy in yeast[J].Curr Opin Cell Biol,2009,21(4):522 -530.
[4] Mehrpour M,Esclatine A,Beau I,et al.Overview of macroautophagy regulation in mammalian cells[J].Cell Res,2010,20(7):748 -762.
[5] Mijaljica D,Prescott M,Devenish RJ.Microautophagy in mammalian cells:revisiting a 40-year-old conundrum[J].Autophagy,2011,7(7):673 -682.
[6] Kissova I,Salin B,Schaeffer J,et al.Selective and non-selective autophagic degradation of mitochondria in yeast[J].Autophagy,2007,3(4):329 -336.
[7] Uttenweiler A,Mayer A.Microautophagy in the yeast Saccharomyces cerevisiae[J].Methods Mol Biol,2008,445:245-259.
[8] Kunz JB,Schwarz H,Mayer A.Determination of four sequential stages during microautophagy in vitro[J].J Biol Chem,2004,279(11):9987 -9996.
[9] Orenstein SJ,Cuervo AM.Chaperone-mediated autophagy:molecular mechanisms and physiological relevance[J].Semin Cell Dev Biol,2010,21(7):719 -726.
[10] Sahu R,Kaushik S,Clement CC,et al.Microautophagy of cytosolic proteins by late endosomes[J].Dev Cell,2011,20(1):131 -139.
[11] Levine B,Klionsky DJ.Development by self-digestion:molecular mechanisms and biological functions of autophagy[J].Dev Cell,2004,6(4):463 - 477.
[12] Sou YS,Waguri S,Iwata J,et al.The Atg8 conjugation system is indispensable for proper development of autophagic isolation membranes in mice[J].Mol Biol Cell,2008,19(11):4762 -4775.
[13] Muller O,Sattler T,Flotenmeyer M,et al.Autophagic tubes:vacuolar invaginations involved in lateral membrane sorting andinverse vesicle budding[J].JCell Biol,2000,151(3):519-528.
[14] Yeung T,Gilbert GE,Shi J,et al.Membrane phosphatidylserine regulates surface charge and protein localization[J].Science,2008,319(5860):210 -213.
[15] Fader CM,Sanchez D,Furlan M,et al.Induction of autophagy promotes fusion of multivesicular bodies with autophagic vacuoles in K562 cells[J].Traffic,2008,9(2):230-250.
[16] Tsunematsu T,Yamauchi E,Shibata H,et al.Distinct functions of human MVB12A and MVB12B in the ESCRT- I dependent on their posttranslational modifications[J].Biochem Biophys Res Commun,2010,399(2):232 -237.
[17] Fader CM,Colombo MI.Autophagy and multivesicular bodies:two closely related partners[J].Cell Death Differ,2009,16(1):70 -78.
[18] Santambrogio L,Cuervo AM.Chasing the elusive mammalian microautophagy[J].Autophagy,2011,7(6):652-654.
[19] Dubouloz F,Deloche O,Wanke V,et al.The TOR and EGO protein complexes orchestrate microautophagy in yeast[J].Mol Cell,2005,19(1):15 - 26.
[20] Uttenweiler A,Schwarz H,Neumann H,et al.The vacuolar transporter chaperone(VTC)complex is required for microautophagy[J].Mol Biol Cell,2007,18(1):166 -175.
[21] Farre JC,Subramani S.Peroxisome turnover by micropexophagy:an autophagy - related process[J].Trends Cell Biol,2004,14(9):515 -523.
[22] Roberts P,Moshitch - Moshkovitz S,Kvam E,et al.Piecemeal microautophagy of nucleus in Saccharomyces cerevisiae[J].Mol Biol Cell,2003,14(1):129 - 141.
[23] Kvam E,Goldfarb DS.Nucleus-vacuole junctions and piecemeal microautophagy of the nucleus in S.cerevisiae[J].Autophagy,2007,3(2):85 -92.
[24] Dawaliby R,Mayer A.Microautophagy of the nucleus coincides with a vacuolar diffusion barrier at nuclear-vacuolar junctions[J].Mol Biol Cell,2010,21(23):4173 - 4183.
[25] Kvam E,Goldfarb DS.Nucleus-vacuole junctions in yeast:anatomy of a membrane contact site[J].Biochem Soc Trans,2006,34(Pt 3):340 -342.
[26] Kraft C,Reggiori F,Peter M.Selective types of autophagy in yeast[J].Biochim Biophys Acta,2009,1793(9):1404-1412.
[27] Kissova I,Deffieu M,Manon S,et al.Uth1p is involved in the autophagic degradation of mitochondria[J].J Biol Chem,2004,279(37):39068 -39074.
[28] Marzella L,Ahlberg J,Glaumann H.In vitro uptake of particles by lysosomes[J].Exp Cell Res,1980,129(2):460-466.
[29] Takikita S,Myerowitz R,Zaal K,et al.Murine muscle cell models for Pompe disease and their use in studying therapeutic approaches[J].Mol Genet Metab,2009,96(4):208-217.
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
河南科技(2022年6期)2022-04-22
生命科学研究(2021年1期)2021-01-25
生物化工(2021年2期)2021-01-19
中国农业科技导报(2020年1期)2020-03-15
读与写(2019年35期)2019-11-05
中国真菌学杂志(2019年1期)2019-01-23
分析化学(2018年7期)2018-09-17
现代职业教育·高职高专(2018年7期)2018-05-14
江苏农业科学(2016年5期)2016-07-23
