
辽河源不同龄组油松天然次生林生物量及空间分配特征
2014-09-19 10:59赵金龙王泺鑫韩海荣康峰峰宋小帅于晓文赵伟红
生态学报 2014年23期
赵金龙,王泺鑫,韩海荣,康峰峰,宋小帅,于晓文,赵伟红
(北京林业大学林学院, 北京 100083)
生物量是指单位面积上所有生物有机体的干重。森林植物群落生物量和生产力是森林生态系统的功能指标,是物质循环和能量流动等方面研究的基础[1],也是研究全球气候变化和碳平衡的基础[2]。在20世纪70年代国际生物学计划(IBP)实施期间,国外[3]就已开始了不同类型森林生物量的研究。国内的冯宗炜[4]、马钦彦[5]等在20世纪80年代采用相对生长法,分别对马尾松和油松生物量进行了研究。此后,一些研究者[6- 7]开展了全国尺度森林植被生物量的研究。目前更多的研究则是集中于落叶松[8- 10]、马尾松[11]、油松[12- 13]、山杨[14]、栎[15- 17]等单个树种的生物量及生产力分配方面。以上研究工作极大地推动了我国森林生物量研究的发展。然而从林分起源上来看,目前的研究对象主要还是人工林,以多个龄组天然林及其次生林生物量及空间分配特征的研究较少。在当前原生林大幅减少,次生林大面积增加的背景下,研究油松次生林的生物量及分配格局具有重要的意义。
油松是中国暖温带落叶林重要的森林树种,其分布范围横跨中国14个省市,地理范围为东经103°20′ —124°45′,北纬31°13′ —43°33′,垂直分布范围为海拔400—1000 m[18]。本试验以4个龄组的油松天然次生林为研究对象,通过计算其乔、灌、草及凋落物层的生物量,揭示林分生物量在时空间尺度上的分配特征,旨在为油松次生林该领域的研究提供科学依据,同时为研究地区该森林类型的可持续经营提供基础。
1 材料与方法
1.1 自然概况
研究地点选在河北省平泉县辽河源自然保护区大窝铺林场(118°22′—118°37′E,41°01′—41°21′N)。该地区处于暖温带向寒温带过渡地带,属于半湿润半干旱大陆性季风型山地气候,年均气温7.3℃。年平均降雨量540 mm,年平均蒸发量1800 mm,湿润度为0.4。土壤主要是棕壤土和褐土,土层深厚。
森林植被主要是油松(P.tabulaeformis)天然林,分布于950—1350 m的阳坡、半阳坡及半阴坡山地,其主要伴生种为辽东栎(Quercusliaotungensis)、蒙古栎(Quercusmongolica)、山杨(Populusdavidiana)、白桦(Betulaplatyphylla)、蒙椴(Tiliamongolica)、山杏(Armeniacasibirica)、大果榆(Ulmusmacrocarpa)等。 灌木层有锦带花(Weigelaflorida)、胡枝子(Lespedezabicolor)、榛(Corylusheterophylla)、毛榛(Corylusmandshurica)、三裂绣线菊(Spiraeatrilobata)、小叶鼠李(Rhamnusparvifolia)、照山白(Rhododendronmicranthum)、迎红杜鹃(Rhododendronmucronulatum)、大花溲疏(Deutziagrandiflora)等。草本层发育较好,以细叶薹草(Carexrigescens)为主,其它组成种主要是野青茅(Deyeuxiapyramidalis)、银背风毛菊(Saussureanivea)、石竹(Dianthuschinensis)、玉竹(Polygonatumodoratum)、花旗竿(Dontostemondentatus)、华北蓝盆花(Scabiosatschiliensis)、歪头菜(Viciaunijuga)、委陵菜(Potentillachinensis)、斑叶兰(Goodyeraschlechtendaliana)等。
1.2 研究方法
1.2.1 样地设计
2012年7— 9月,根据国家林业局《森林资源规划设计调查主要技术规定》(2003)和《国家森林资源连续清查技术规定》(2003)中对油松天然林的龄组划分标准(油松天然林幼龄林 ≤ 30a、中龄林31—50a、近熟林51—60a、成熟林61—80a),选择林相整齐、具有代表性的油松天然次生林地段作为固定标准地。在上述试验林内共设计18块 (幼龄林4块、中龄林6块、近熟林5块、成熟林3块)面积为600 m2(20 m×30 m)的固定样地。各林分详细资料见表1。

表1 油松天然次生林标准地概况
1.2.2 生物量测定
目前获得森林中乔木层生物量的方法主要有皆伐法、平均木法和回归估计法(相对生长法) 3种。鉴于本次研究对象是保护区的油松天然林,因此采用回归估计法测定其生物量。在计算区域尺度森林或林分生物量方面,方精云[19- 20]认为主要方法有3类:平均生物量法[21]、平均换算因子法[22- 24]和换算因子连续函数法[25];张志等[26]则认为主要有平均生物量法(MBM)、生物量扩展因子法 (BEFM)[27]、连续生物量扩展因子法 (CBEFM) 和IPCC法等方法。论文采用平均生物量法计算了4个龄组油松天然次生林林分生物量。
(1)乔木层
对样地内油松每木检尺,测量胸径、树高、株数、枝下高等测树因子。然后计算出各龄组林木的平均胸径,以2 cm为一个径级,选取在平均胸径附近的3株立木作为平均标准木进行树干解析,4个龄组共计12株标准木(多次向当地林业局申请才得以在实验区采伐)。所有标准木均采用 “分层切割法” 进行研究,即在树干1.3 m处分段,以上按每2 m (树高h>15 m) 或者1 m (树高h<15 m) 长度分段,直到对应的梢头不足2 m或者1 m (森林生态系统长期定位观测方法 (LY/T 1952—2011))。在标准木干基部和每段分层处均截取一个5 cm厚的圆盘,带回实验室将每个圆盘的树皮与树干木质分离,并称取各自鲜重,利用Lintab 5年轮分析仪测定其年轮宽度并进行定年。实测每段“分层”内油松干材、干皮、树枝、叶及球果(摘除每一层内所有的树叶和球果,分干球果和湿球果) 的鲜重,并对每棵标准木的以上组分分别取样。采用全挖法挖取根系,对根桩、粗根 (>5 cm)、大根 (2—5 cm)、细根 (1—2 cm) 和小细根 (<1 cm) 分类处理,称取鲜重并分别取样。
(2)林下植被及凋落物层
①灌木层 在每块标准地 (20 m×30 m) 外选取与林内植物分布特征类似的地段,按品字形布置5个2 m×2 m 小样方,记录灌木种名、株数、盖度等信息后,采用全部收获法分别测定所有灌木 (不分种类) 叶、茎及根系的总鲜重,并分别取样;
②草本层 在每块灌木小样方 (2 m × 2 m) 的左下角分别设置1块 1 m×1 m的小样方,记录下种名、株数、盖度等资料后,同样采用全部收获法分地上和地下2部分称取鲜重并取样;
③凋落物层 在每块标准地外设置1块1 m×1 m的小样方,按照 (森林生态系统长期定位观测方法 (LY/T 1952—2011)) 中对未分解层、半分解层和全分解层的划分标准对现存凋落物分别称取鲜重并取样。
将测定鲜重后的以上取样样品放入85℃烘箱中烘干至恒重 (间隔1h称重质量无变化),然后测定各样品干重、计算含水率。根据样品干重与湿重比推算各组分的生物量,最后将各龄组计算出的乔、灌、草及凋落物平均生物量累加得到对应的林分生物量。
1.2.3 计算公式
(1)乔木生物量计算方程
根据油松各标准木生物量实测数据,分别以D、H、DH、D2和D2H为自变量进行了多种回归模型拟合。结果表明:方程 (1) 或 (2) 拟合叶、枝、干、干皮和根生物量整体效果最好,其判定系数 (R2) 在0.775—0.975之间,F检验的相关性均达到显著 (P<0.01) 或极显著水平 (P<0.001) (表2)。而球果生物量则以D2为自变量的二次项方程 (3) 拟合最好,判定系数 (R2) 为0.887,F检验的相关性均达到显著(P=0.004)。
W=a(D2H)b
(1)
或
lnW=lna+bln(D2H)
(2)
W=b0-b1D2+b2(D2)2
(3)
式中,D为胸径 (m),H为树高 (m),W为生物量 (t),V为蓄积量 (m3),a、b、b0、b1和b2均为常数。
本研究还采用Brown和Lugo[28]及方精云[7]提出的利用生物量与蓄积量间存在线性关系估测地上森林生物量的方程 (公式(4)) 进行了油松标准木生物量的研究,同样表现出较好的拟合效果(表2)。
W=a·V+b
(4)
(5)
(6)
式中,W为生物量 (t),V为标准木材积(m3),gi为第i区分段中央断面积 (m2),li为第i区分段长度 (m),gn为梢头木底断面积 (m2),l′为梢头木长度 (m),n为总区分段数。M为林分的蓄积量 (m3/hm2),ni为第i级中标准木株数,k为分级级数 (i=1, 2,...,k),Gi为第i级的胸高断面积 (m2/hm2),Vij和gij分别是为第i级中第j株标准木的材积 (m3) 和胸高断面积 (m2)。
(2)林分生物量的计算公式
计算出不同龄组油松天然次生林乔木层、灌木层、草本层及枯落物层4个层次生物量,累加后按样地取平均值,得到对应龄组下的林分生物量:
(7)
式中,Wi为第i个龄组林分生物量 (t/hm2),wTij、wSij、wHij和wLij分别为第i个龄组下第j块样地乔木层、灌木层、草本层和凋落物层的生物量 (t/hm2),i为龄组 (i= {1,2,3,4};分别对应幼龄林、中龄林、近熟林和成熟林),n为各龄组油松林样地数量。
1.2.4 统计分析
文中所有统计分析采用spss 18.0软件包,作图采用制图软件Sigmaplot 10.0。
2 结果与分析
2.1 异速生长方程
以D2H和V为自变量与油松叶、枝、干、干皮及根各组分生物量建立的2种异速生长方程均能表现出较好的拟合效果(表2),判定系数(R2)均较高。通过对方程F检验结果的比较分析,本研究采用方程1计算乔木层以下各组分的生物量。而球果生物量则通过方程W=0.006-0.486D2+8.996(D2)2(公式3)计算。

表2 计算乔木层各组分生物量的2种异速生长方程比较
2.2 乔木层生物量及分配
2.2.1 乔木层各组分生物量及分配
从表3可以看出,油松幼龄林林分密度与其他龄组有显著差异,幼龄林密度最高 (1967株/hm2),近熟林最低 (747株/hm2)。乔木层生物量成熟林 (373.128 t/hm2)>近熟林 (224.991 t/hm2)>中龄林 (187.750 t/hm2)>幼龄林 (119.169 t/hm2)。每公顷生物量、样地生物量和单木生物量的大小均随林龄的增大而增大。
乔木层地上部分生物量最大的组分是干(45.749—231.660 t/hm2),所占比例大小为:中龄林 (66.27%)>近熟林 (64.35%)>成熟林 (62.09%)>幼龄林 (38.39%)。生物量最小的是球果 (0.959—3.710 t/hm2),大小顺序是成熟林 (1.02%)>幼龄林 (0.92%)>近熟林 (0.76%)>中龄林(0.51%)(表4)。

表3 乔木层生物量
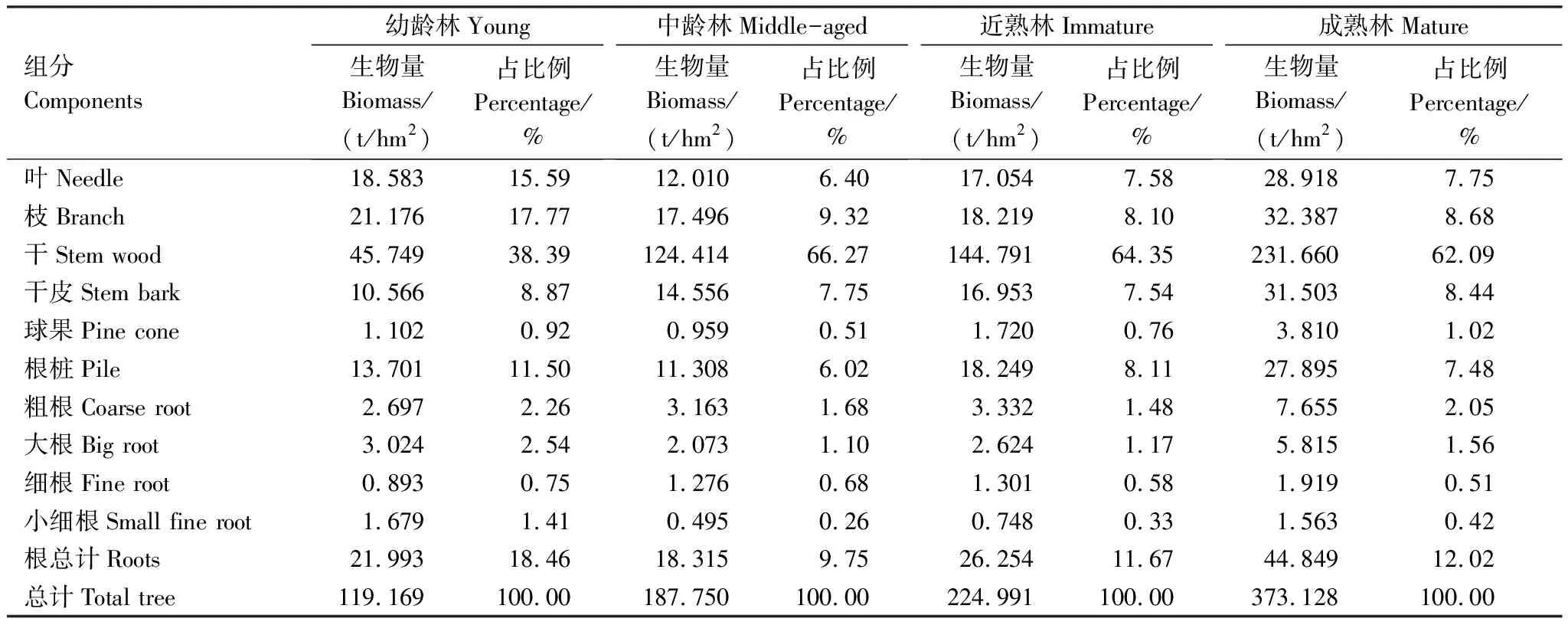
表4 乔木层各组分生物量及分配

图1 乔木层各组分生物量结构特征
在林分水平上,各龄组油松叶、枝、干皮、球果及不同径级根系等组分的生物量分布均相对集中,特别是中龄林和近熟林离散度较低;异常值 (干)随林龄的增大而增大,而且离散程度有递增的趋势 (图1)。各龄组林分地下生物量占总生物量比例依次是幼龄林 (18.46%)>成熟林 (12.02%)>近熟林 (11.67%)>中龄林 (9.75%)。地上生物量与地下生物量之间的Person相关系数高达0.934 (解析木) 和0.874 (样地),均呈显著线性相关(图2)。

图2 地上生物量与地下生物量的关系
在地下各组分生物量大小比较中,幼龄林表现为:细根(0.893 t/hm2)<小细根 (1.679 t/hm2)<粗根 (2.697 t/hm2)<大根 (3.024 t/hm2)<根桩 (13.701 t/hm2);而其他龄组均表现为:根桩>粗根>大根>细根>小细根。根桩生物量在各龄组根系生物量中所占百分比均大于60%,大小顺序是近熟林>幼龄林>成熟林>中龄林(表5)。

表5 根系组分生物量及分配
2.2.2 乔木层各组分生物量的空间结构
生物量的空间结构是指植物各组分生物量在空间上的配置状况,它在很大程度上反映了植物对光能、水分和养分的利用效率[29]。本研究采用选取的回归方程,结合各龄组解析木生物量在空间结构上的垂直分布特征,对标准地中林木各组分生物量的空间结构进行了研究(图3)。结果表明:各龄组油松林各“层次”内干和干皮的生物量随树高的增加均表现出递减的趋势,而枝、叶及球果生物量则是先增加到一个峰值后才递减。从幼龄林至成熟林,其他组分生物量的和分别在5.6、14.6、12.6 m和16.6 m处超过了干的生物量,其中幼龄林的枝叶较为茂盛,在所有龄组中所占比例最大。叶的生物量在空间结构上存在差异,幼龄林在地上3.6 m开始分布,中龄林和近熟林均在离地面4.6 m处,而成熟林则高达6.6 m。球果的生物量在各“层次”组分生物量中占比例较小,随树干高度增加有增大的趋势,幼龄林球果高度分布最低 (4.6 m)。在林分中上层,各组分生物量大小表现为干>枝>叶>干皮>球果。
2.3 林下植被层生物量及分配
灌木层生物量大小随林龄的增大而增大,表现为:成熟林(0.861 t/hm2)>近熟林 (0.790 t/hm2)>中龄林 (0.559 t/hm2)>幼龄林 (0.401 t/hm2),成熟林的生物量是幼龄林的2.14倍。灌木各组分生物量随林龄的增大而增加,且均表现为根>茎>叶。草本层在各龄组油松天然林下均生长较好,在林下植被层生物量中占主导地位(表6)。草本层生物量的大小顺序为:幼龄林 (3.058 t/hm2)>近熟林 (2.017 t/hm2)>中龄林(1.220 t/hm2)>成熟林 (1.181 t/hm2)。草本层各组分生物量所占比例差异显著,近熟林地上部分生物量仅占20.92%,而幼龄林则高达31.20%。

表6 林下植被层生物量及分配

图3 乔木层地上部分各组分生物量的空间结构特征
2.4 凋落物层生物量及分配
凋落物在森林中起到固持水分、保育土壤等重要作用,每年林地都能积累大量来自林冠层的枯枝、落叶、花果及死树皮等类型的凋落物。由表7可以看出,各龄组油松林林下凋落物各组分厚度及生物量均表现为未分解层>半分解层>全分解层。凋落物生物量大小关系为:成熟林 (22.623 t/hm2)>近熟林 (14.390 t/hm2)>中龄林 (14.272 t/hm2)>幼龄林 (10.265 t/hm2)。
2.5 林分生物量及分配
林分生物量的准确测定是合理估算森林生态系统碳贮量及分配格局的基础,对全球碳循环的研究具有重要意义。在本研究中,成熟林 (397.793 t/hm2)>近熟林 (242.188 t/hm2)>中龄林 (203.801 t/hm2)>幼龄林 (132.894 t/hm2),成熟林的生物量是幼龄林的近3倍。在各龄组林分不同层次生物量的比较中,均表现为:乔木层>凋落物层>草本层>灌木层。乔木层、灌木层和凋落物层生物量随林龄的增大而增加,而草本层生物量则呈S型曲线递减(表8)。

表8 油松天然次生林林分生物量及分配
3 结论与讨论
(1)林分生物量大小顺序是成熟林>近熟林>中龄林>幼龄林。其中,乔木层生物量成熟林 (373.128 t/hm2)>近熟林 (224.991 t/hm2)>中龄林 (187.750 t/hm2)>幼龄林 (119.169 t/hm2),表现出生物量随林分年龄的增大而增大,这种趋势与前人的研究结果[30- 33]一致。在具体龄组生物量大小的比较中,幼龄林生物量与孙巧玉等[34]的研究结果 (97.22—122.20 t/hm2) 吻合,而中龄林则与程小琴等[13](38a油松) 的研究结果一致。但从各龄组生物量来看,则比Cao等[33]对北京地区油松生物量的研究结果要高。这可能与本研究区域相对较好的外界环境有关。
(2)乔木层地上部分各组分生物量的研究中,中龄林和成熟林大小顺序与多种森林类型的研究结果[9- 17]一致,表现为:干>根>枝>干皮>叶>球果;而幼龄林和近熟林则是干>根>枝>叶>干皮>球果,这与Noh等[31]对赤松和 Li 等[35]对白皮松的研究结果一致。虽然干在乔木组分中占据较大比重,但是干生物量占乔木层生物量的比例在不同龄组间差异却非常显著[32]。在本研究中,此比例的大小关系是中龄林 (66.27%)>近熟林 (64.35%)>成熟林 (62.09%)>幼龄林 (38.39%)。幼龄林叶生物量之所以大于干皮生物量是因为林木处于快速生长期,光合器官相对发达,近熟林则是林分密度过低,枝叶茂盛所致。
地下部分根系生物量大小顺序是成熟林>近熟林>幼龄林>中龄林,幼龄林生物量大于中龄林也是因为中龄林林分密度相对较低。本研究根组分生物量大小顺序的结果与赵金龙等[11]对针叶和阔叶树的研究结果一致,表现为:根桩>粗根>大根>细根>小细根。
根冠比被认为是一个指示林木累积生物量及其分配方式的重要指标[36],在整个林分水平上呈线性关系。本研究此线性方程的平均斜率为0.145,比其他温带松类森林[37- 39]的值略小。
(3)林下植被及凋落物层的总生物量在整个林分生物量中所占比例较小,幼龄林占10.33%、中龄林占7.88%、近熟林占7.10%,成熟林占6.20%。林下植被层生物量在本研究的4个龄组间为1.779—3.459 t/hm2,和Li等[32]在红松 (1.61—3.76 t/hm2) 和Cao等[33]在油松 (0.87—3.55 t/hm2) 林下植被生物量的研究结果一致。这可能与森林经理、林冠结构、土壤条件等有关,进而通过光照、营养及水热条件影响林下植被生长[30]。
凋落物层生物量随林分年龄的增大而增大,大小关系表现为:成熟林 (22.623 t/hm2)>近熟林 (14.390 t/hm2)>中龄林(14.272 t/hm2)>幼龄林 (10.265 t/hm2),与Li等[35]在白皮松次生林的研究结果一致。
致谢:感谢北京林业大学林学院陈锋、张彦雷给予的帮助。
:
[1] Luo J, Yang Z, Yang Q W. A study on the biomass and production of forest on the Gongga mountain. Acta Phytoecologica Sinica, 2000, 24(2): 191- 196.
[2] Rizvi R H, Dhyani S K, Yadav R S, Singh R. Singh Biomass production and carbon stock of poplar agroforestry systems in Yamunanagar and Saharanpur districts of northwestern India. Current Science, 2011, 100(5): 736- 742.
[3] Leith H, Whittaker R H. Primary Productivity of the Biosphere. New York: Springer Verlag. 1975.
[4] Feng Z W, Chen C Y, Zhang J W, Wang K P, Zhao J L, Gao H. Determination of biomass ofPinusMassonianastand in Huitong county, Hunan province. Scientia Silvae Sinicae, 1982, 18(2): 127- 134.
[5] Ma Q Y. A study on the biomass of Chinese pine forests. Journal of Beijing Forestry University, 1989, 11(4): 1- 10.
[6] Wu G, Feng Z W. Study on the social characteristics and biomass of thepinustabulaeformisforest systems in China. Acta Ecologica Sinica, 1994, 14(4): 415- 422.
[7] Fang J Y, Liu G H, Xu S L. Biomass and net production of forest vegetation in China. Acta Ecologica Sinica, 1996, 16(5): 497- 508.
[8] Wang Y H, Zhou G S, Jiang Y L, Yang Z Y. Estimating biomass and NPP ofLarixforests using forest inventory data(FID). Acta Phytoecologica Sinica, 2001, 25(4): 420- 425.
[9] Sun Y J, Zhang J, Han A H, Wang X J, Wang X J. Biomass and carbon pool ofLarixgmeliniyoung and middle age forest in Xing′an Mountains Inner Mongolia. Acta Ecologica Sinica, 2007, 27(5): 1757- 1761.
[10] Ju W Z, Wang X J, Sun Y J. Age structure effects on stand biomass and carbon storage distribution ofLarixolgensisplantation. Acta Ecologica Sinica, 2011, 31(4): 1139- 1148.
[11] Zhao J L, Liang H W, Wen Y G, Lu L H, Ming A G, Su J M. Distribution pattern of biomass in the mixed uneven aged stands ofPinusmassonianaandCastanopsishystrix. Journal of Central South University of Forestry & Technology, 2011, 31(2): 60- 64.
[12] Chen T R, Feng J, Ma Q Y, Feng Z K, Zhang S Z. Linear compatible models of tree layer biomass ofPinustabulaeformis plantations in Xiaolong Mountains. Chinese Journal of Ecology, 2008, 27(3): 317- 322.
[13] Cheng X Q, Han H R, Kang F F. Biomass, carbon accumulation and its partitioning of aPinustabulaeformisplantation ecosystem in Shanxi Province, China. Chinese Journal of Ecology, 2012, 31(10): 2455- 2460.
[14] Shi S D, Zhao P W, Zhou M, Yang X D. Biomass and carbon storage of the secondary forest (Populusdavidiana) at different stand growing stages in southern Daxinganling temperature zone. Ecology and Environmental Sciences, 2012, 21(3): 428- 433.
[15] Liu Y C, Wu M Z, Guo Z M, Jiang Y X, Liu S R, Wang Z Y, Liu B D, Zhu X L. Biomass and net productivity ofQuercusvariabilisforest in Baotianman Natural Reserve. Chinese Journal of Applied Ecology, 1998, 9(6): 11- 16.
[16] Li Y Y, Fan H B, Lin D X, Su B Q, Liu C H, Sun X. Biomass and distribution of stands mixedPinusmassonianawith broad leaved species. Journal of Zhejiang Forestry College, 2004, 21(4): 388- 392.
[17] Wang J, Chen Y M, Cao Y, Zhou J Y, Hou L. Carbon concentration and carbon storage in different components of naturalQuercuswutaishanicaforest in Ziwuling of Loess Plateau, Northwest China. Chinese Journal of Ecology, 2012, 31(12): 3058- 3063.
[18] Zhang Z M, Chang S, Zhang Y Z. Research on biomass carbon density of different kinds ofPinustabulaeformisForests. Shanxi Forestry Science and Technology, 2012, 41(3): 11- 13.
[19] Fang J Y. Forest productivity in China and its response to global climate change. Acta Phytoecologica Sinica, 2000, 24(5): 513- 517.
[20] Fang J Y, Chen A P, Zhao S Q, Ci L J. Estimating biomass carbon china′s forests: Supplementary notes on report published in science (291: 2320- 2322) by Fang et al. Acta Phytoecologica Sinica, 2002, 26(2): 243- 249.
[21] Brown S, Lugo A E. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica, 1982, 14(3): 161- 187.
[22] Turner D P, Koepper G J, Harmon M E, Lee J J. A carbon budget for forests of the conterminous United State. Ecological Applications, 1995, 5(2): 421- 436.
[23] Alexeyev V, Birdsey R, Stakanov V, Korotkov I. Carbon in vegetation of Russian forests: methods to estimate storage and geographical distribution. Water, Air and Soil Pollution, 1995, 82(1/2): 271- 282.
[24] Fang J Y, Wang G G, Liu G H, Xu S L. Forest biomass of China: an estimate based on the biomass-volume relationship. Ecological Applications, 1998, 8(4): 1084- 1091.
[25] Fang J Y, Wang Z M. Forest biomass estimation at regional and global levels, with special reference to China′s forest biomass. Ecological Research, 2001, 16(3): 587- 592.
[26] Zhang Z, Tian X, Chen E X, He Q S. Review of methods on estimating forest above ground biomass. Journal of Beijing Forestry University, 2011, 33(5): 144- 150.
[27] Lehtonena A, Makipaa R, Heikkinen J, Sievänena R, Liskic J. Biomass expansion factors (BEFs) forScotspine,Norwayspruceandbirchaccording to stand age for boreal forests. Forest Ecology and Management, 2004, 188(1/3): 211- 224.
[28] Brown S, Lugo A E. Biomass of tropical forests: A new estimate based on forest volumes. Science, 1984, 223(4642): 1290- 1293.
[29] Fan H B, Li Y Y, Su B Q, Lin D X, Liu C H, Jiang Z K. Allocation pattern of biomass and productivity in the mixed uneven-aged stands of Masson′s pine and hardwood species. Acta Ecologica Sinica, 2006, 26(8): 2464- 2473.
[30] Peichl M, Arain M A. Above-and belowground ecosystem biomass and carbon pools in an age-sequence of temperate pine plantation forests. Agricultural and Forest Meteorology, 2006, 140(1): 51- 63.
[31] Noh N J, Son Y, Lee S K, Seo K W, Heo S J, Yi M J, Park P S, Kim R H, Son Y M, Lee, K H. Carbon and nitrogen storage in an age-sequence ofPinusdensiflorastands in Korea. Science China Life Sciences, 2010, 53(7): 822- 830.
[32] Li X, Yi M J, Son Y, Park P S, Lee K H, Son Y M, Kim R H, Jeong M J. Biomass and carbon storage in an age-sequence of Korean pine (Pinuskoraiensis) plantation forests in central Korea. Journal of Plant Biology, 2011, 54(1), 33- 42.
[33] Cao J, Wang X, Tian Y, Wen Z, Zha T. Pattern of carbon allocation across three different stages of stand development of a Chinese pine (Pinustabulaeformis) forest. Ecological Research, 2012, 27(5), 883- 892.
[34] Sun Q Y, Liu Y, Li G L, Zhang S, Xu F, Wang W W. Effect of slope position on above-ground biomass distribution pattern inPinustabulaeformisplantation. Journal of Central South University of Forestry & Technology, 2012, 32(9): 102- 105.
[35] Li C, Zha T, Liu J, Jia X. Carbon and nitrogen distribution across a chronosequence of secondarylacebarkpinein China. The Forestry Chronicle, 2013, 89(2), 192- 198.
[36] Litton C M, Ryan M G, Tinker D B, Knight D H. Belowground and aboveground biomass in youngpostfirelodgepolepineforests of contrasting tree density. Canadian Journal of Forest Research, 2003, 33(2), 351- 363.
[37] King J S, Giardina C P, Pregitzer K S, Friend A L. Biomass partitioning in red pine (Pinusresinosa) along a chronosequence in the Upper Peninsula of Michigan. Canadian Journal of Forest Research, 2006, 37(1), 93- 102.
[38] Tobin B, Nieuwenhuis M. Biomass expansion factors for Sitka spruce (Piceasitchensis(Bong.) Carr.) in Ireland. European Journal of Forest Research, 2007, 126(2): 189- 196.
[39] Peichl M, Arain M A. Allometry and partitioning of above-and belowground tree biomass in an age-sequence ofwhitepineforests. Forest Ecology and Management, 2007, 253(1): 68- 80.
参考文献:
[1] 罗辑, 杨忠, 杨清伟. 贡嘎山森林生物量和生产力的研究. 植物生态学报, 2000, 24(2): 191- 196.
[4] 冯宗炜, 陈楚莹, 张家武, 王开平, 赵吉录, 高虹. 湖南会同地区马尾松林生物量的测定. 林业科学, 1982, 18(2): 127- 134.
[5] 马钦彦. 中国油松生物量的研究. 北京林业大学学报, 1989, 11(4): 1- 10.
[6] 吴刚, 冯宗炜. 中国油松林群落特征及生物量的研究. 生态学报, 1994, 14(4): 415- 422.
[7] 方精云, 刘国华, 徐嵩龄. 我国森林植被的生物量和净生产量. 生态学报, 1996, 16(5): 497- 508.
[8] 王玉辉, 周广胜, 蒋延玲, 杨正宇. 基于森林资源清查资料的落叶松林生物量和净生长量估算模式. 植物生态学报, 2001, 25(4): 420- 425.
[9] 孙玉军, 张俊, 韩爱惠, 王雪军, 王新杰. 兴安落叶松(Larixgmelini)幼中龄林的生物量与碳汇功能. 生态学报, 2007, 27(5): 1757- 1761.
[10] 巨文珍, 王新杰, 孙玉军. 长白落叶松林龄序列上的生物量及碳储量分配规律. 生态学报, 2011, 31(4): 1139- 1148.
[11] 赵金龙, 梁宏温, 温远光, 卢立华, 明安刚, 苏建苗. 马尾松与红锥混交异龄林生物量分配格局. 中南林业科技大学学报, 2011, 31(2): 60- 64.
[12] 程堂仁, 冯菁, 马钦彦, 冯仲科, 张宋智. 小陇山油松林乔木层生物量相容性线性模型. 生态学杂志, 2008, 27(3): 317- 322.
[13] 程小琴, 韩海荣, 康峰峰. 山西油松人工林生态系统生物量_碳积累及其分布. 生态学杂志, 2012, 31(10): 2455- 2460.
[14] 史山丹, 赵鹏武, 周梅, 杨小丹. 大兴安岭南部温带山杨天然次生林不同生长阶段生物量及碳储量. 生态环境学报, 2012, (3): 428- 433.
[15] 刘玉萃, 吴明作, 郭宗民, 蒋有绪, 刘世荣, 王正用, 刘保东, 朱学凌. 宝天曼自然保护区栓皮栎林生物量和净生产力研究. 应用生态学报, 1998, 9(6): 11- 16.
[16] 李燕燕, 樊后保, 林德喜, 苏兵强, 刘春华, 孙新. 马尾松林混交阔叶树的生物量及其分布格局. 浙江林学院学报, 2004, 21(4): 388- 392.
[17] 王娟, 陈云明, 曹扬, 周建云, 侯磊. 子午岭辽东栎林不同组分碳含量与碳储量. 生态学杂志, 2012, 31(12): 3058- 3063.
[18] 张志明, 常帅, 张亚珍. 不同类型油松林生物量碳密度的研究. 山西林业科技, 2012, 41(3): 11- 13.
[19] 方精云. 中国森林生产力及其对全球气候变化的响应. 植物生态学报, 2000, 24(5): 513- 517.
[20] 方精云, 陈安平, 赵淑清, 慈龙骏. 中国森林生物量的估算: 对Fang等Science一文(Science, 2001, 291: 2320—2322)的若干说明. 植物生态学报, 2002, 26(2): 243- 249.
[26] 张志, 田昕, 陈尔学, 何祺胜. 森林地上生物量估测方法研究综述. 北京林业大学学报, 2011, 33(5): 144- 150.
[29] 樊后保, 李燕燕, 苏兵强, 林德喜, 刘春华, 蒋宗垲. 马尾松-阔叶树混交异龄林生物量与生产力分配格局. 生态学报, 2006, 26(8): 2464- 2473.
[34] 孙巧玉, 刘勇, 李国雷, 张硕, 许飞, 王巍伟. 坡位对油松人工林地上生物量分配格局的影响. 中南林业科技大学学报, 2012, 32(9): 102- 105.
猜你喜欢
江西农业大学学报(2022年1期)2022-02-04
浙江林业科技(2021年5期)2021-10-19
林业资源管理(2021年2期)2021-06-07
江西农业大学学报(2021年2期)2021-05-16
林业勘查设计(2021年2期)2021-04-08
土壤学报(2020年6期)2021-01-05
农民致富之友(2020年8期)2020-05-11
广东农业科学(2019年2期)2019-04-09
农业与技术(2018年16期)2018-11-28
现代农业科技(2017年10期)2017-07-12
