
樟子松子代测定林高碳汇家系选择研究
2016-09-13 02:41张含国张明远张吉利王千雪于宏影
西南林业大学学报 2016年1期
刘 灵 张含国 张明远 张吉利 王千雪 于宏影
(1.中国林业科学院林业新技术所,北京 100091;2.国家林业局哈尔滨林业机械研究所,黑龙江 哈尔滨 150086;3.林木遗传育种国家重点实验室(东北林业大学),黑龙江 哈尔滨 150040)
樟子松子代测定林高碳汇家系选择研究
刘灵1,2张含国3张明远2张吉利2王千雪2于宏影2
(1.中国林业科学院林业新技术所,北京 100091;2.国家林业局哈尔滨林业机械研究所,黑龙江 哈尔滨 150086;3.林木遗传育种国家重点实验室(东北林业大学),黑龙江 哈尔滨 150040)
以黑龙江省齐齐哈尔市龙江县错海实验林场32年生樟子松自由授粉家系子代测定林为试验材料,测定并分析其生长性状、含碳率及树干碳储量的家系遗传变异,估算各性状遗传增益,筛选含碳率高、树干碳储量大的优良家系。结果表明:树干材积、树干生物量和树干碳储量家系间存在丰富的遗传变异,变异系数分别为31.421%、31.998%和31.903%;木材基本密度和含碳率遗传变异较小,变异系数分别为2.725% 和1.868%。相关分析结果表明,含碳率与树高、胸径、林木树干材积、树干生物量及树干碳储量之间均存在微弱的负相关关系;树干碳储量与树高、胸径、树干材积及树干生物量成极显著正相关。家系K70和A64入选为优良家系,2个家系的平均树干碳储量为19.903 kg,比家系平均树干碳储量高出36.89%。
樟子松;子代测定林;生长性状;含碳率;碳储量:遗传变异
樟子松(Pinussylvestrisvar.mongolica)是欧洲赤松的一个变种,其天然分布于我国大兴安岭及呼伦贝尔草原的红花尔基沙地,材性性状表现良好,树干通直,具有速生、抗寒、抗旱耐贫瘠等优良特性,适合生长于沙地,起到防风固沙等作用。因此,我国北方半干旱风沙地区选择樟子松营造防风固沙林、水土保持林、防护林和用材林[1-4]。
国内的林木良种选育工作始于20世纪50年代,樟子松良种选育主要集中在优树选择、建立母树林及种子园的建立及管理、杂交育种和地理变异规律的探讨[5-7]。为了解樟子松的地理变异规律,国内学者开展了大量的种源试验[8-9],结果表明樟子松不同地理种源种子的品质存在显著差异,各材性性状间也存在极显著差异,证明种源选择有效。
在森林碳汇研究方面,国外研究人员已取得较大进展,主要表现在对不同地区、不同树种、不同林龄的碳储量及碳密度的估算[10-13]。日本,俄罗斯和美国在森林碳储量研究领域取得的成果比较突出[14-16]。日本学者对Sugi和Hinkoi 2个地区的森林碳储量进行估算[17],其总碳储量分别为346.4×106Mg和139.2×106Mg,碳密度分别为76.81 Mg/hm2和58.01 Mg/hm2,远高于日本西南部地区碳储量。美国估算国内森林生态系统碳储量为36.7 Pg,其中土壤中碳储量占50%,活立木占33%,其次为枯枝落叶层及地下根部[18]。国内对森林碳储量的研究开始于20世纪70年代,大都是采用生物量与含碳率系数乘积的研究方法。基于不同地区、不同树种及不同测定方法(蓄积量法、样地清算法及遥感法等),国内外学者也提出适应不同方法的估算模型[19-21]。
基于森林植被生物量及碳储量方法估算碳密度及碳储量时,需要根据所估算的树种选取合适的含碳率,国内外学者多选用国际公认的森林生态系统含碳率平均值进行计算(0.45~0.50)[22]。显然,这样的估算结果不够精确,不能准确地反映森林的固碳能力。因此,有学者直接测定不同树种和相同树种不同器官的含碳率,分析讨论不同树种及相同树种不同器官间含碳率的差异。Thomas S C等[23]在对长白山地区的不同树种、不同器官的含碳率进行估算测定得出,不同树种间的含碳率存在显著差异,数值范围48.4%~51.0%,不稳定碳的含量也不可以忽略不计,平均值为2.2%,在不同树种间存在明显的差异。研究表明,针叶树种的含碳率为(50.8%±0.1%),要高于阔叶树种(49.5%±0.2%)[24]。
国内外对于碳汇树种的研究已经较为成熟,但是研究多数以树种、林分、器官为对象。本研究采取以家系为研究对象,直接测定试验材料不同器官的含碳率,更准确的估算出试验材料的碳储量,以进行樟子松优良家系选择,提出培育技术模式,为提高林分的碳储量提供优良材料。
1 研究方法
1.1研究对象
研究地设于黑龙江省齐齐哈尔市龙江县错海林场,林场海拔400 m,地处东经122°51′,北纬47°27′。年降水量350~450 mm,年蒸发量1 500 mm,年平均气温3.4 ℃,无霜期129 d。研究地土层深厚、疏松,肥力中等,为暗棕壤,透水性良好,pH 6.8。在研究地以樟子松优树自由授粉子代测定实验林,子代测定林于1981年7月定植,株行距2.0 m×2.0 m,采用完全随机区组设计,4株小区,6次重复,共21个处理,分别为A10和、A15、A17、A4和、A50、A64、A8、CK、K1、K14、K16、K17、K2、K20、K27、K32、K46、K48、K49、K60、K70,总面积共2 352 m2。2011年11月测量试验林的21个家系284样株的树高、胸径。
1.2取样方法
2011年11月对错海子代测定试验林前5次重复进行木材取样,在胸高处同一方向用直径5 mm的生长锥取得由树皮至髓心的完整无疵木芯,每个小区随机选取2株树作为样本,每个家系5次重复共10个试样,共测定210个试样。
1.3数据测定及分析
采用饱和含水率法测定木材基本密度,利用德国耶拿专家型总有机碳/总氮分析仪(multi N/C3100 TOC)测定含碳率。
干材生物量及碳储量的计算:
C=B×Cc
B=Vρj
本文通过构建均衡移动模型来模拟目标价格变化对中国大豆市场均衡产生的效果,进而分析对政府财政支出和经济福利变化的影响,分析结论归纳如下:
V=(h+3)g1.3f
式中:C为碳储量;B为生物量;Cc为含碳率;V为树干材积;f为樟子松平均试验形数,f=0.41;h为树高;g1.3为胸高处断面积;ρj为木材密度。
家系的生长性状及各项其他指标的分析均采用小区平均数分析,采用SPSS 18.0软件进行性状方差分析,生长性状相关分析,进而估算家系遗传力和性状间遗传相关,采用Ducan法进行各性状的差异显著性检验以筛选最优家系。
2 结果与分析
2.1生长和材性的性状遗传变异分析
樟子松各家系生长性状及材性性状测定表明:樟子松树干材积、树干生物量及树干碳储量家系间遗传变异较大,这为速生及高碳汇优良家系的选择提供了丰富的变异基础。基本密度与含碳率家系间变异系数相对较小,结果见表1。

表1 生长性状遗传变异分析
由表1可知,树干材积家系变异系数为31.421%,置信区间为0.102~0.114 m3。树干材积家系内也存在丰富的遗传变异,变异最大的家系为A64,变异系数为39.891%,置信区间为0.111~0.167 m3。变异最小的家系A4和变异系数为8.615%,置信区间变异范围是0.053~0.109 m3。家系的最大变异系数是最小的3.6倍,变异系数最大的家系大于家系平均值26.956%。
树干生物量的家系平均变异系数31.998%,置信区间是30.161~33.899 kg。树干生物量家系内也存在较大的遗传差异,其中家系K32的变异系数最大,为40.979%,置信区间是26.511~42.596 kg。家系A4和的变异系数最小,为8.464%,置信区间是15.604~32.688 kg。家系的最大变异系数是最小家系的3.8倍,家系最大的变异系数大于家系平均28.067%。
树干碳储量的家系平均变异系数31.903%,置信区间是13.095~14.711 kg。樟子松树干碳储量家系内也存在较大的遗传变异,其中家系K32的变异系数最大,为40.935%,置信区间是11.679~19.085 kg。家系A4和的变异系数最小,为8.156%,置信区间变异范围是6.815~14.222 kg。家系的最大变异系数是最小家系的4倍。家系最大的变异系数大于家系平均28.311%。
樟子松家系子代测定林木材基本密度家系变异系数为2.725%,置信区间是0.295~0.298 g/cm3。含碳率的家系变异系数为1.868%,置信区间是43.331%~43.638%。
对各性状进行方差分析,结果表明,家系间树高和材积性状均差异极显著,家系遗传力分别为49.33%和45.92%,受中等强度遗传控制,说明生长性状进行家系水平的改良具有一定的潜力。各生长性状的方差分析及遗传参数估算见表2。树高、材积、树干生物量、碳储量多重比较结果见表3。
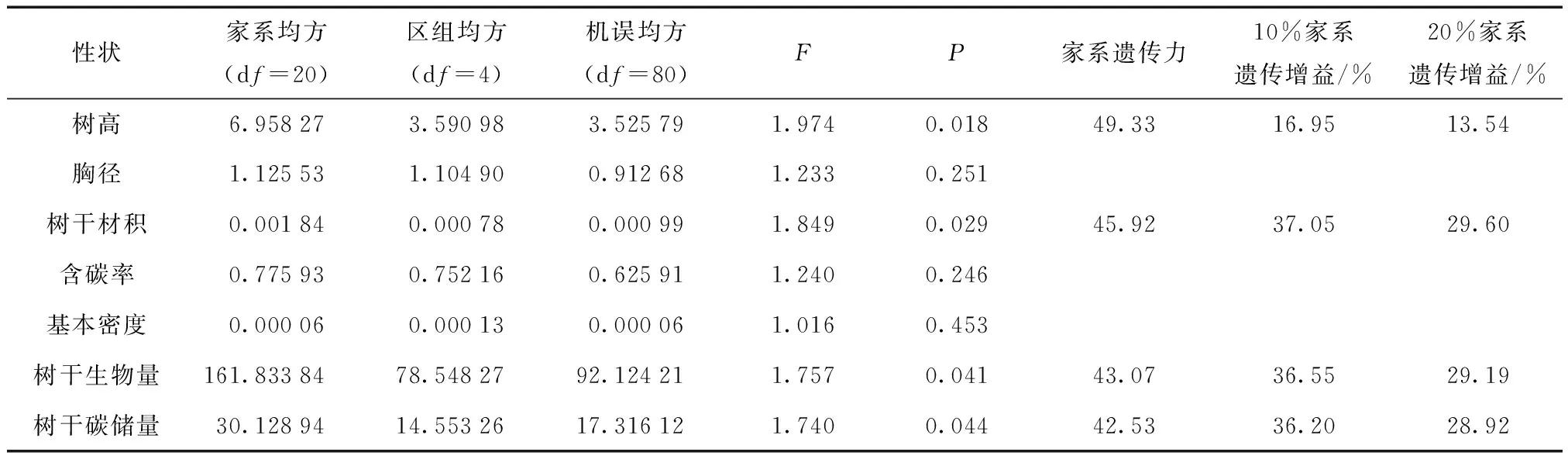
表2 生长性状的方差分析及遗传参数估算
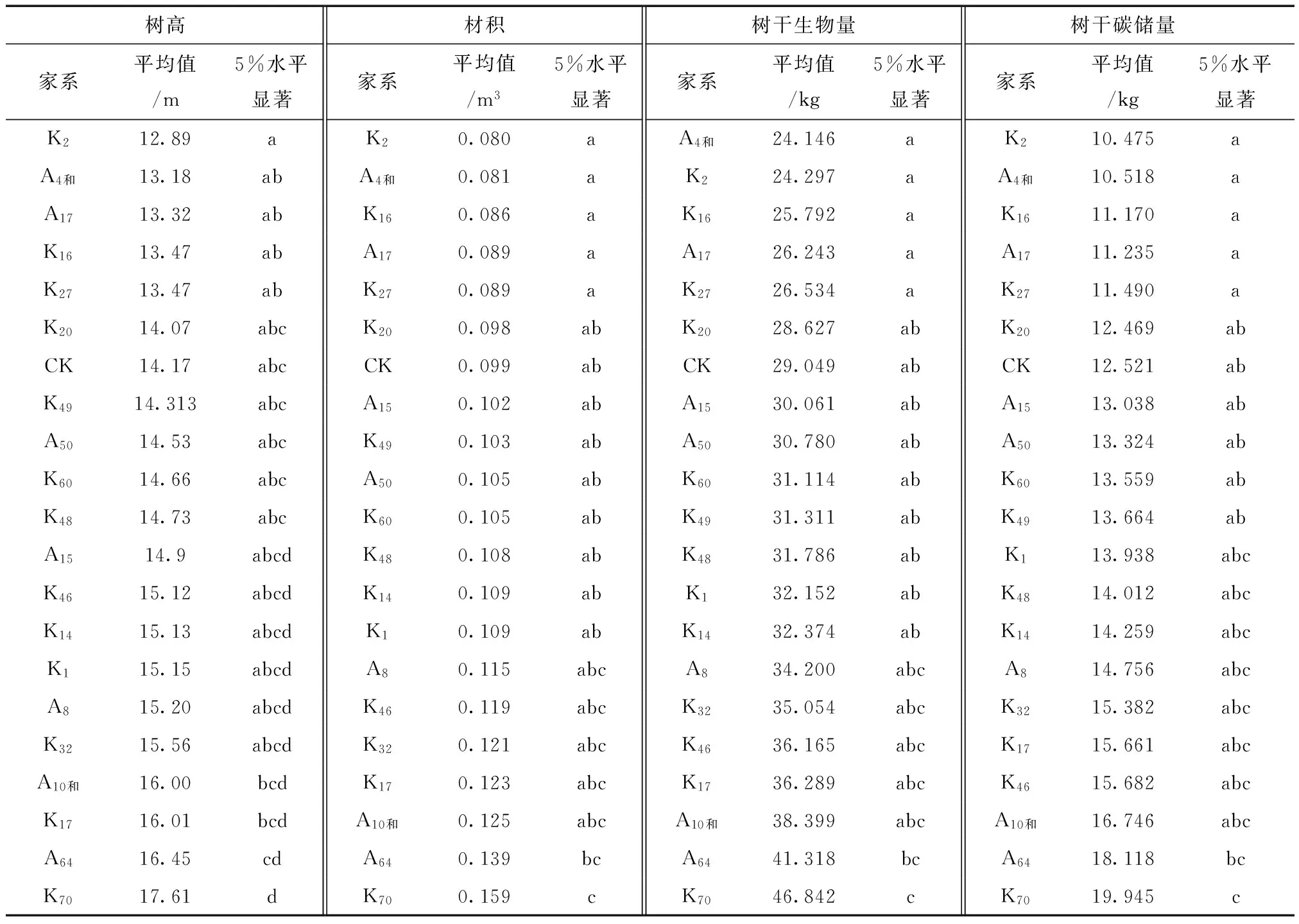
表3 树高、材积多重比较
由表3可知,树高较高的家系是K70和A64,两者树高平均值比胸径最小的家系K2高32.12%,比家系树高平均值高出15.39%。材积较大的是家系K70和A64,两者材积平均值比材积最小的家系K2高86.25%,比家系材积平均值高37.96%。入选率为10%时,优良家系树高和材积的遗传增益分别为16.95%和37.05%。入选率为20%时,优良家系树树高和材积的遗传增益分别为13.54%和29.60%。
家系间树干生物量,树干碳储量差异显著。家系遗传力分别是43.07%和42.53%,受中等强度的遗传控制。多重比较结果显示树干生物量较大的家系是K70和A64,比树干生物量最小的家系A4和高出82.56%,比树干生物量家系平均值高出37.76%。树干碳储量最大的家系是K70和A64,比树干碳储量最小的家系K2高出81.68%,比树干碳储量家系平均值高出36.89%。入选率为10%时,优良家系树干生物量和树干碳储量的遗传增益分别为36.55%和36.20%。入选率为20%时,优良家系树干生物量和树干碳储量的遗传增益分别为29.19%和28.92%。
各性状间相关关系见表4。

表4 各性状间相关关系
注:**表示1%水平显著,*表示5%水平显著。
由表4可知,树高与胸径、林木材积、树干生物量和树干碳储量成极显著正相关,相关系数为0.452、0.970、0.962和0.960。胸径与林木材积、树干生物量和树干碳储量成极显著正相关,相关系数为0.608、0.607和0.608。木材含碳率与树高、胸径、林木材积、树干生物量及树干碳储量之间均存在微弱的负相关关系。木材基本密度与树高、胸径、林木材积、树干生物量与树干碳储量存在极微弱的正相关关系。在对木材基本密度及含碳率选择时可独立进行。树干碳储量与树高、胸径、材积、和树干生物量呈极显著正相关。在对林木碳储量进行选择时,可以对其他几项进行联合选择。
3 结论与讨论
1) 樟子松自由授粉子代测定林家系试验中,树干生物量及树干碳储量遗传变异较大。木材基本密度及含碳率遗传变异较小。其中,树高、材积、树干生物量和树干碳储量差异显著。家系遗传力分别为49.33%、45.92%、43.07%和42.52%,属于中等遗传。
2) 对各性状进行方差分析,树高、林木材积、树干生物量和树干碳储量差异显著。K70和A64在树高、林木材积、树干生物量和树干碳储量等方面均表现最优异,作为优良家系被选入,2个家系的树干碳储量分别为19.945、18.118 kg,2个家系的平均树干碳储量为19.903 kg,比家系平均树干碳储量高出36.89%。以现有试验林造林密度为标准,用家系K70造林,林分碳储量可以达到28.0×103kg/hm2,比现有林分碳储量高2.36倍,固碳效果会得到显著提高。
刘文线[25]分析了29年生樟子松半同胞子代测定林树高、胸径及材积的变异,结果表明树高的家系变异系数为9.80%,胸径的家变异系数为 18.85%,材积的变异系数42.41%。本试验得出的家系间胸径变异较小,树高变异稍大,可能受定植密度的影响,材积的变异与本试验基本相符。王全波等[26]对樟子松5个天然种源子代苗期的高及地径进行生长变异分析,结果表明,家系间苗木地径生长存在丰富的变异,变异系数为21.99%~33.17%,家系间苗木高生长存在丰富的遗传变异,变异系数为29.63%~44.11%。可以看出樟子松在苗期阶段变异较大。
周志春等[27]在1998年测定马尾松(Pinusmassoniana)木材性状时得到结论,可以用胸高处任一半径方向的木芯样品,以代表树干均值。段喜华等[28]在1997年测定长白落叶松(Larixgmelini)株内变异时采用非破坏性取样方法,并得到相同结论,在基本密度及管胞长度方面,胸高处任一方位半径的样品测定值,可代表树干平均值。贾庆彬等[29]在2012年长白落叶松含碳率遗传变异及高固碳种源和家系研究一文中指出,利用1.3 m处圆盘含碳率或木芯样品含碳率推算树干平均含碳率均可行。基于大量试验结论的基础上,本试验采用的取样方法是木芯取样法,属于非破坏性取样。
通过性状相关分析可以了解各性状间及指标的连带关系,为多性状联合选择提供理论依据。综合分析显示,树干碳储量占地上部分碳储量的比例达到80%左右,且树干碳储量与生物量,树干材积成极显著正相关,说明生长性状和生物量优异对林木的固碳能力有积极促进作用,说明可以用树干碳储量代表地上部分碳储量进行选优,可以节省大量的工作量。这与张超等[30]在日本落叶松(Larixkaempferi)碳储量家系遗传变异及优良家系选择中的研究一致。
[1]朱教君,康宏樟,许美玲,等.外生菌根真菌对科尔沁沙地樟子松人工林衰退的影响[J]. 应用生态学报, 2007, 18(12):2693-2698.
[2]徐化成,李长喜,唐谦.樟子松、常白松、赤松的引种研究[J].林业科学研究,1991,4(1):44-49.
[3]Zhu J J, Fan Z P, Zeng D H, et al. Comparison of stand structure and growth between artificial and natural forests of Pinus sylvestiris var. mongolica on sandy land[J]. Journal of Forestry Research, 2003, 14(2):103-111.
[4]底国旗, 沈海龙.内蒙古大兴安岭林区樟子松天然分布与资源状况[J].东北林业大学学报, 2006,34(1) :101-103.
[5]陈瑶, 朱万才.樟子松人工林胸径生长规律的研究[J].林业科技情报,2010, 42(2):26-27.
[6]肖杰, 李广玉, 赵勇,等.樟子松种源家系苗木高生长变异规律研究[J].林业科技, 2004, 29(3):1-3.[7]张树根, 张泽宁, 聂向东.樟子松遗传改良研究现状及育种策略[J].林业调查规划,2008, 33(5): 116-118.[8]刘桂丰, 褚延广, 时玉龙,等.17年生帽儿山地区樟子松种源试验[J].东北林业大学学报, 2003, 31(4):1-3.
[9]赵欣欣, 王丽, 王永长,等.樟子松的良种选育及建园方法[J].中国西部科技, 2011, 10(35):34.
[10]闫德仁, 闫婷.内蒙古森林碳储量估算及其变化特征[J].林业资源管理, 2010, 6(3):31-33.
[11]关芳, 高畅, 任汉屏.基于森林蓄积估算鹤立林业局的森林生物量碳储量[J].林业勘察设计, 2010(3):11-12.
[12]李峰, 刘桂英, 王力刚.黑龙江省森林碳汇价值评价及碳汇潜力分析[J].防护林科技, 2011(1):87-88.
[13]焦燕, 胡海清.黑龙江省森林植被碳储量及动态变化[J].应用生态学报,2005, 12(16):2248-2252.
[14]Strohbach M W, Haase D. Above-ground carbon storage by urban trees in Leipzig, Germany: analysis of patterns in a European city[J]. Landscape & Urban Planning, 2012, 104(1):95-104.
[15]Fang S, Xue J, Tang L. Biomass production and carbon sequestration potential in poplar plantations with different management patterns.[J]. Journal of Environmental Management, 2007, 85(3):672-679.
[16]Nowak D J, Greenfield E J, Hoehn R E, et al. Carbon storage and sequestration by trees in urban and community areas of the United States[J]. Environmental Pollution, 2013, 178c(1):229-236.
[17]Fukuda M, Iehara T, Matsumoto M. Carbon stock estimates for sugi and hinoki forests in Japan[J]. Forest Ecology & Management, 2003, 184(3):1-16.[18]Schroeder P, Brown S, Mo J, et al. Biomass estimation for temperate broadleaf forests of the United States using inventory data[J]. Forest Science, 1997, 43(3):424-434.[19]刘萍,邓鉴锋,魏安世,等. 广州市森林生物量及碳储量的评估[J].西南林业大学学报,2015,35(4):62-65.
[20]Man Y S, Miah M D, Lee K H. Potential contribution of the forestry sector in Bangladesh to carbon sequestration[J]. Journal of Environmental Management, 2007, 82(2):260-276.
[21]Bert D, Danjon F. Carbon concentration variations in the roots, stem and crown of maturePinuspinaster(Ait.)[J]. Forest Ecology & Management, 2006, 222:279-295.
[22]马钦彦, 陈遐林, 王娟,等.华北主要森林类型建群种的含碳率分析[J].北京林业大学学报,2002, 24(5/6): 96-100.[23]Thomas S C, Malczewski G. Wood carbon content of tree species in Eastern China: Interspecific variability and the importance of the volatile fraction[J]. Journal of Environmental Management, 2007, 85(3):659-662.
[24]贾炜玮, 于爱民.樟子松人工林单木生物量模型研究[J].林业科技情报, 2008,40(2):1-2.
[25]刘文线. 樟子松半同胞子代遗传增益分析[J]. 林业勘查设计,2013(4):12-14.[26]王全波, 李朝毅, 宋纯彦,等. 樟子松5个天然种源子代苗期遗传变异分析[J]. 林业科技, 2007(3):7-8.
[27]周志春, 王章荣, 陈天华,等.马尾松木材性状株内变异与木材取样方法的探讨[J].南京林业大学学报, 1988(4):52-60.
[28]段喜华, 张含国, 潘本立,等.长白落叶松木材材性株内变异[J].东北林业大学学报, 1997,25(2):33-36.
[29]贾庆彬, 张含国, 王美玲,等.长白落叶松家系碳含量遗传变异与高固碳家系选择[J].东北林业大学学报, 2013, 41(5):28-32.
[30]张超, 张含国, 朱航勇等.日本落叶松碳储量家系遗传变异及优良家系选择研究[J].中南林业科技大学学报, 2011, 31(10):53-62.
(责任编辑曹龙)
Selection of High Carbon Storage Families fromPinussylvestrisvar.mongolicaProgeny Testing Forest
Liu Ling1,2,Zhang Hanguo3,Zhang Mingyuan2,Zhang Jili2,Wang Qianxue2,Yu Hongying2
(1. Research Institute for Forestry New Technology, Beijing 100091, China;2.Harbin Research Institute of Forestry Machinery, State Forestry Administration, Harbin Heilongjiang 150086, China;3.State Key Laboratory of Forest Genetics and Tree Breeding,Northeast Forestry University,Harbin Heilongjiang 150040,China)
The 32-years-oldPinussylvestrisvar.mongolicaopen-pollinated progeny testing forest were sampled as the research object, which set on the Longjiang Cuohai Experimental Station in Heilongjiang province, to analyze the family genetic variation of growth traits, carbon content and stem carbon storage, and estimate their genetic gain, then filter the superior families of high carbon storage and carbon density. The results showed that there were plentiful genetic variation in trunk volume, biomass and carbon storage and the variation coefficient was 31.421%, 31.998% and 31.903%, respectively. The genetic variations in wood basic density, carbon content was relatively small, and the variation coefficient was 2.725% and 1.868%. There was weak negative correlation between carbon content with the other traits. Trunk carbon storage was significantly positive correlated with tree height, diameter at breast height (DBH) volume and trunk biomass. Synthesize the above results, K70and A64had been chosen as the superior families. The mean value of carbon storage was 19.903 kg, which was higher than mean value of all families by 36.89%.
Pinussylvestrisvar.mongolica; progeny testing forest; growth traits; carbon density; carbon storage; genetic variation

2015-07-24
中国林业科学院林业新技术所基本科研业务费专项(CAFINT2015C15)资助;中国林业科学院基本科研业务费专项(CAFYBB2014QA027)资助。
张含国(1962—),男,教授,博士生导师。研究方向:落叶松、红松常规育种。Email:hanguozhang1@sina.com。
10.11929/j.issn.2095-1914.2016.01.010
S722.3
A
2095-1914(2016)01-0057-06
第1作者:刘灵(1988—),女,助理工程师。研究方向:樟子松常规育种。Email:liulingln@126.com。
猜你喜欢
广西林业科学(2022年3期)2022-07-08
海南医学(2020年1期)2020-01-18
电子制作(2019年24期)2019-02-23
浙江农林大学学报(2019年1期)2019-01-24
江苏农业科学(2018年14期)2018-08-08
中南林业科技大学学报(2017年5期)2017-12-20
中国康复理论与实践(2015年10期)2015-12-24
中南林业科技大学学报(2014年6期)2014-12-28
食品工业科技(2014年5期)2014-03-11
卷宗(2012年5期)2012-10-21
