
不同生境条件茶园大型土壤动物群落结构与季节动态
2017-07-21 04:30李金林欧丽丽黄贞辉黄丽芳魏少娜许佳敏邢树文
韩山师范学院学报 2017年3期
李金林,欧丽丽,黄贞辉,黄丽芳,魏少娜,许佳敏,邢树文
(韩山师范学院食品工程与生物科技学院,广东潮州521041)
不同生境条件茶园大型土壤动物群落结构与季节动态
李金林,欧丽丽,黄贞辉,黄丽芳,魏少娜,许佳敏,邢树文*
(韩山师范学院食品工程与生物科技学院,广东潮州521041)
对凤凰山和坪溪镇茶区不同生境条件的茶园大型土壤动物进行调查,分析生态茶园与非生态茶园大型土壤动物群落与营养类群组成及其动态差异,探讨了影响两种茶园大型土壤动物群落结构及动态变化的主要因素.结果表明:生态茶园捕获大型土壤动物22目69科1835只,占土壤动物总捕获量的60.34%;非生态茶园捕获大型土壤动物19目53科1 206只,占土壤动物总捕获量的39.66%.生态茶园各类群的个体数均高于非生态茶园.夏季、秋季和冬季生态茶园土壤动物的优势类群比例低于非生态茶园,常见类群高于非生态茶园.茶园土壤动物的数量、种类、Shannon指数和Simpson指数均表现为秋季高,春季次之,冬季低.生态茶园捕食性动物是主要功能类群,占总捕获量的38.53%;非生态茶园腐食性动物为主要功能类群,占总捕获量的41.29%.生态茶园夏季和秋季的各功能类群的数量、种类、Shannon指数和丰富度指数均高于非生态茶园,春季和冬季捕食性动物的数量和种类及多样性指数高于非生态茶园,植食性和杂食性动物均低于非生态茶园.RDA排序结果表明,生态茶园与非生态茶园土壤动物之间存在明显的分布梯度,土壤动物的分布受土壤环境因子(全N、全K、土壤有机质、土壤含水量、土壤温度和茶园植被量)的影响.因此,茶园周边环境条件是影响茶园土壤动物群落结构的重要因素,气候因子是影响土壤动物季节动态分布的主要因素.
生态茶园;非生态茶园;大型土壤动物;群落;季节动态;功能类群
茶叶种植是潮州市的主要农业经济产业,随着茶叶价格不断提高,茶农为追求经济收入大面积地开荒植茶,对茶园周边的生态环境造成了严重的破坏,严重影响茶园生态环境及土壤微生态环境.土壤是陆地生态系统中物质循环和能量交换的主要场所,而土壤动物直接或间接地反映了土壤环境的变化[1].土壤的理化性质、化学性质、农业技术措施以及土地利用类型等对土壤动物群落组成及分布有一定的影响[2-6].而且土壤动物的活动范围小、迁移能力弱,它们与环境间具有相对稳定的关系[7-9].因此,土壤动物被看作是评价土壤质量变化的敏感性指示生物,物种的组成及其群落结构会随着环境的变化而改变[10-11].研究茶园土壤动物群落组成及功能团结构及其动态对建立生态茶园、保护茶园生态环境意义重大.勾影波[12]等报道了茶园土壤动物群落结构的多样性,申燕[13]对茶园土壤动物群落结构特征及影响因素进行了系统研究.关于不同生境条件茶园土壤动物群落多样性及季节动态的研究少见报道.本文选择地形和规模较为典型的广东省潮州市凤凰山区生态茶园和浮滨镇的生境破坏型茶园作为研究对象,探讨不同茶园大型土壤动物群落结构特征及其季节动态变化,探讨茶园周边生境条件、土壤生态环境对茶园大型土壤动物群落结构的影响,揭示不同茶园大型土壤动物组成与土壤环境的关系,旨在为生态茶园建设提供有价值的参考.
1 研究地区与方法
1.1 样地概况
凤凰山位于广东潮州市潮安区凤凰镇(116°22′~116°49′E、23°26′~24°00′N),地势自西北向东南倾斜,西、北、东边境多山地,属莲花山系,峰峦迭翠,绵延成串的山地构成了粤东南丘陵,阻挡寒潮南侵,故常年气温较高,夏长冬暖,春秋相连.年温差和日较差均小,年平均温度21-22℃.
生态型茶园:茶园位于潮安区凤凰镇叫水坑村,海拔850 m,茶园面积约1 000亩.茶园周围是毛竹(Phylostachysheterocycla(Carr.)Mitford cv.Pubescens)及阔叶混交林、灌木,茶园伴生革命菜(GynuracrepidioidesBenth)、芒萁(Dicranopterisdichotoma(Thunb.)Bernh.)、野牡丹(Melastomacan⁃didumD.Don)、霍香蓟(Ageratumconyzoides L.)等多种植物.茶园为梯田式造田,茶树行距2.0 m,间距40 cm.每年春季4月人工采春茶1次,秋后整枝和人工除草,平均每两年施有机肥1次,茶园不施用农药与除草剂.
非生态型茶园:茶园位于广东饶平县浮滨镇,海拔380 m,面积6万余亩.该区域的丘陵山坡地带均开发为茶园,茶园周边自然生境遭到严重破坏,茶园连成片,形成单一茶园.茶园伴生极少量鸡矢藤(Paederiascandens(Lour.)Merr.)及空心莲子草(Alternantheraphiloxeroides(Mart.)Griseb)等植被.梯田式茶园,茶树行距1.5 m.每年采茶4次以上,施肥2次以上并使用低毒农药.
两种茶园生境的基本概况如表1:

表1 两种茶园生境的基本概况
1.2 样地设置与标本采集
于2015年春季(5月2~8日)、夏季(8月8~16日)、秋季(10月25日~11月5日)和冬季(1月19~25日)4个季节对生态茶园和非生态茶园进行取样调查,在叫水坑村选取5块生态茶园,在坪溪镇选取5块非生态茶园作为研究样地,每块样地设5个样方作为取样点,每个样方为50 cm×50 cm.采用手捡法收集其中的大型土壤动物.同一样地同一土层的标本合并保存在80%酒精中,带回实验室在体视显微镜下进行分类鉴定.利用《中国土壤动物检索图鉴》[14-17]等,在体视镜下对土壤动物进行鉴定和分类统计,一般鉴定到科,统计土壤动物的类群和数量.
植物生物量:将样方内的植物采集带回实验室鉴定,然后称重记录,再烘干称干重.
动物生物量:将鉴定后的动物烘干,称干重.
1.3 土壤理化指标测定
采用鲁如坤[18]对土壤理化性质的测定方法,以烘干法测定土壤含水量,采用重铬酸钾容量法-外加热法测定土壤有机质含量,采用半微量开氏法测定土壤全氮,采用氢氧化钠熔融火焰光度法测定土壤全钾[19].
1.4 数据处理
土壤动物的丰度按以下标准划分:根据原始捕获量占捕获总量的百分比来划分各类群数量等级,即个体数量大于捕获总量的10.0%以上者为优势类群,占1.0%~10.0%者为常见类群,不足1.0%者为稀有类群.
土壤动物物种多样性指数按如下公式进行计算[19]:
(1)Shannon-Wiener多样性指数H′=-∑Pi1nPi
(3)Pielou均匀度指数J=H′/lnS
(4)Margalef丰富度指数SR=(S-1)/lnN
式中,Pi为第i类群或物种的个体数量占土壤动物总数的比率,S为土壤动物群落所有动物类群数,ni为第i物种的个体数,N为土壤动物群落全部类群的个体总数,Hmax为最大多样性指数.
(5)群落相似性分析
采用Jaccard系数[20]见式:q=C/(A+B-C)
式中C为2个群落的共有类群数,A和B分别为2个群落中各自的土壤动物类群数,0<q<0.25为极不相似,0.25≤q<0.5为中等不相似,0.5≤q<0.75为中等相似,0.75≤q<1.0为极为相似.
数据运用Excel(2010版)进行统计处理,采用SPSS16.0软件对数据进行分析,采用单因素方差分析(One-way ANOVA)比较分析各项土壤理化性质及各指数的差异性.
2 结果与分析
2.1 生态茶园与非生态茶园大型土壤动物的优势度及季节动态
本次调查的生态茶园四个季节共获得大型土壤动物22目69科1 835只,占土壤动物总捕获量的60.34%;非生态茶园四季共获得大型土壤动物19目53科1 206只,占土壤动物总捕获量的39.66%.生态茶园土壤动物优势类群的个体数高于非生态茶园,而生态茶园土壤动物优势类群的相对丰度(25.89%)却低于非生态茶园(31.67%)(表2).生态茶园各季节土壤动物常见类群和稀有类群的个体数所占比例均高于非生态茶园(表3),春季的常见类群除外.生态茶园土壤动物的优势类群为寡毛纲和蚁科,非生态茶园的优势类群为寡毛纲和线蚓科,生态茶园土壤动物常见类群有20类,占该茶园土壤动物个体总数的58.31%,稀有类群有46类,占该茶园土壤动物个体总数的15.80%;非生态茶园常见类群有19类,占该茶园土壤动物个体总数的57.38%,稀有类群32类,占该茶园土壤动物个体总数的10.95%.两种茶园共有常见类群有13类,共有的稀有类群有20类.
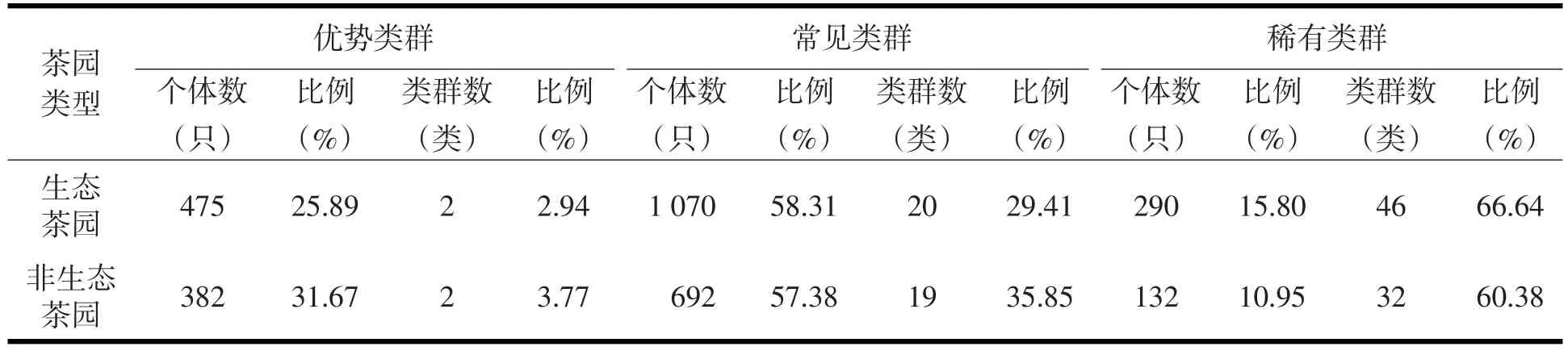
表2 不同茶园土壤动物优势度组成
生态茶园春季土壤动物的优势类群为寡毛纲(Oligcchacte)和蚁科(Formicidae),夏季和冬季为寡毛纲,秋季为蚁科.非生态茶园中,春季与生态茶园相同,夏季为线蚓科(Enchytraeidae)、寡毛纲和蜚蠊科(Blattidae),秋季和冬季为线蚓科、寡毛纲.春季生态茶园土壤动物优势类群的个体丰度高于非生态茶园,生态茶园类群数的相对度低于非生态茶园;夏季、秋季和冬季土壤动物优势类群个体丰度与类群数的相对丰度均低于非生态茶园(表3).
生态茶园春季土壤动物常见类群的个体数相对丰度低于非生态茶园,类群数相对丰度低于非生态茶园;夏季、秋季和冬季的常见类群个体丰度均高于非生态茶园,类群数的比例两者高低不同.生态茶园的常见类群有丝尾螋科(Diplatyidae)、歩甲科(Carabidae)、狼蛛科(Lycosidae)、跳虫科(Po⁃duridae)、蜚蠊科(Blattidae)、地蜈蚣目(Geophilomorpha)、石蜈蚣科(Lithobiidae)、光盔蛛科(Liocranidae)、管巢蛛科(Clubionidae)9类,非生态茶园共有的常见类群有丝尾螋科、歩甲科、狼蛛科和叶甲科(Chrysomelidae)4类.生态茶园常见类群的个体丰度是秋、冬季高于春、夏季;非生态茶园春、秋和冬季较高,夏季较低(表3).两个茶园常见类群的个体数差异大,相对丰度生态茶园高于非生态茶园(表2).在四个季节中生态茶园稀有类群的个体数、类群比例均高于非生态茶园.两茶园稀有类群的个体数和类群比例显示为秋季高,冬季低.
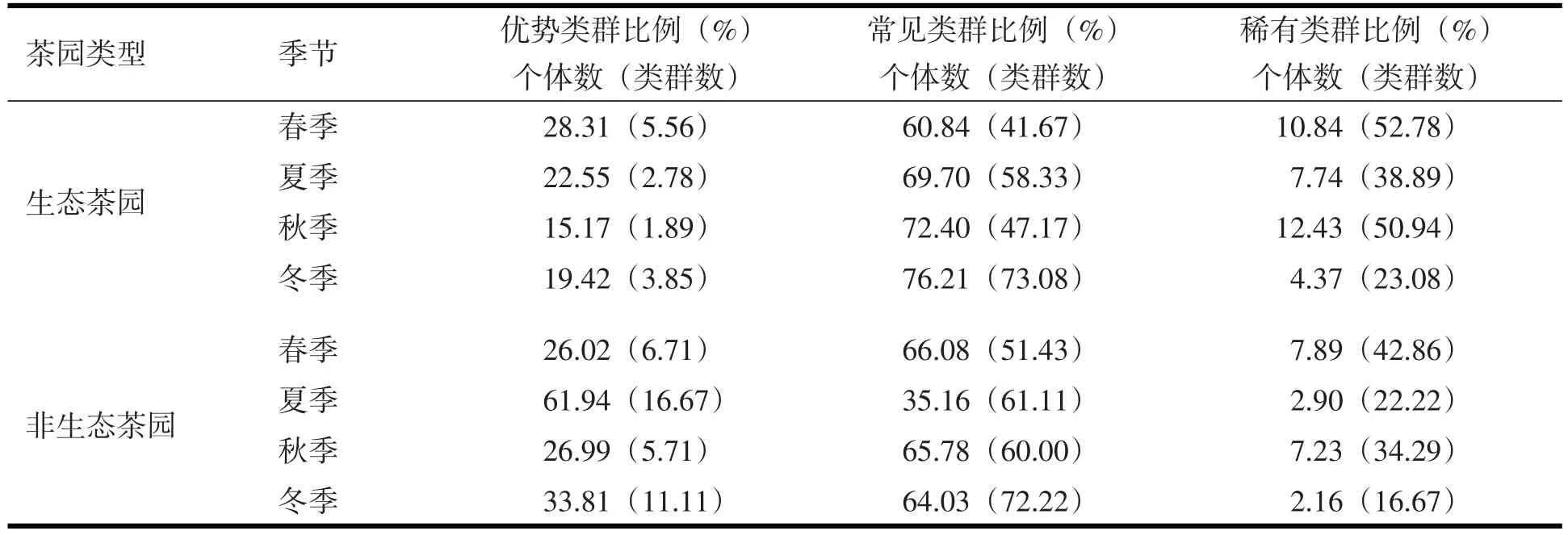
表3 不同茶园土壤动物优势度动态
2.2 生态茶园和非生态茶园大型土壤动物多样性及季节动态
对两类茶园土壤动物群落组成及结构特征分析表明,生态茶园大型土壤动物的个体数、类群数、多样性指数、优势性指数和丰富度指数均高于非生态茶园(图1).表明茶园及其周边生境对大型土壤动物的群落多样性具有重大影响.
生态茶园土壤动物数量、种类和丰富度指数在不同季节表现出一定的差异,冬季显著低于春季、夏季和秋季,秋季显著高于春季、夏季和冬季(图1-Ⅰ、图1-Ⅱ、图1-Ⅵ).多样性指数表现为秋季显著高于春、夏和冬季(图1-Ⅲ);优势性指数秋季显著高于冬季,且秋季和冬季均与春季和夏季无显著性差异(图1-Ⅴ);各个季节的均匀度指数间均无显著差异(图1-Ⅳ).
非生态茶园土壤动物个体数是冬季显著低于春季、夏季和秋季,且秋季最高(图1-Ⅰ);物种数、多样性指数、优势性指数和丰富度指数,春、秋季显著高于夏、冬季(图1-Ⅱ、图1-Ⅲ、图1-Ⅴ、图1-Ⅵ);均匀度指数,夏季显著低于秋季、冬季,而春季、秋季和冬季无显著性差异(图1-Ⅳ).
综合分析两个茶园大型土壤动物群落组成及多样性季节变化,两茶园土壤动物数量、种类、Shannon指数均表现为秋季>春季>夏季>冬季,且非生态茶园上述各指数均低于生态茶园.两茶园的丰富度指数均在冬季最低,而生态茶园的最高值出现在秋季,非生态茶园出现在春季,且生态茶园四个季节的丰度指数均高于非生态茶园.

图1 不同茶园大型土壤动物多样性的季节动态
2.3 生态茶园和非生态茶园大型土壤动物功能类群组成及季节动态
不同生境下的茶园,不同功能的土壤动物表现出不同的群落组成特征.依据土壤动物大致的食性特征,将土壤动物分为植食性、捕食性、杂食性和腐食性4个功能类群.
从表4看出,生态茶园捕食性动物的个体数占总捕获量的38.53%,是其主要功能类群;非生态茶园腐食性动物个体数占总捕获量的41.29%,是其主要功能类群.植食性动物在两茶园中所占的比例最小,分别是9.75%、10.78%.除均匀度指数外,生态茶园捕食性土壤动物的各指数在四个食性中最高;除个体数外,非生态茶园植食性动物的各生态指数在四个功能团中最高.
分析两个茶园不同功能团,结果表明,植食性和捕食性土壤动物中,除均匀度指数外,生态茶园的个体数、类群数、多样性指数、优势性指数和丰富度指数高于非生态茶园;杂食性动物中,除个体数外,生态茶园其余各指数均低于非生态茶园;而生态茶园腐食性动物各指数均高于非生态茶园.
2.3.1 生态茶园大型土壤动物功能类群组成及多样性
个体数分析,植食性动物类群秋季显著高于夏季和冬季(P<0.05),与春季无显著性差异;杂食性动物类群秋季显著高于春季、夏季和冬季(P<0.05);捕食性和腐食性动物类群冬季显著低于春季、夏季和秋季(P<0.05)(图2-Ⅰ).
类群数分析,植食性土壤动物类群秋季显著高于春季、夏季和冬季(P<0.05);捕食性动物类群冬季显著低于春季、夏季和秋季(P<0.05),秋季显著高于春季和冬季,与夏季无显著性差异;杂食性与腐食性动物类群秋季显著高于冬季(P<0.05),与春季和夏季无显著性差异(图2-Ⅱ).
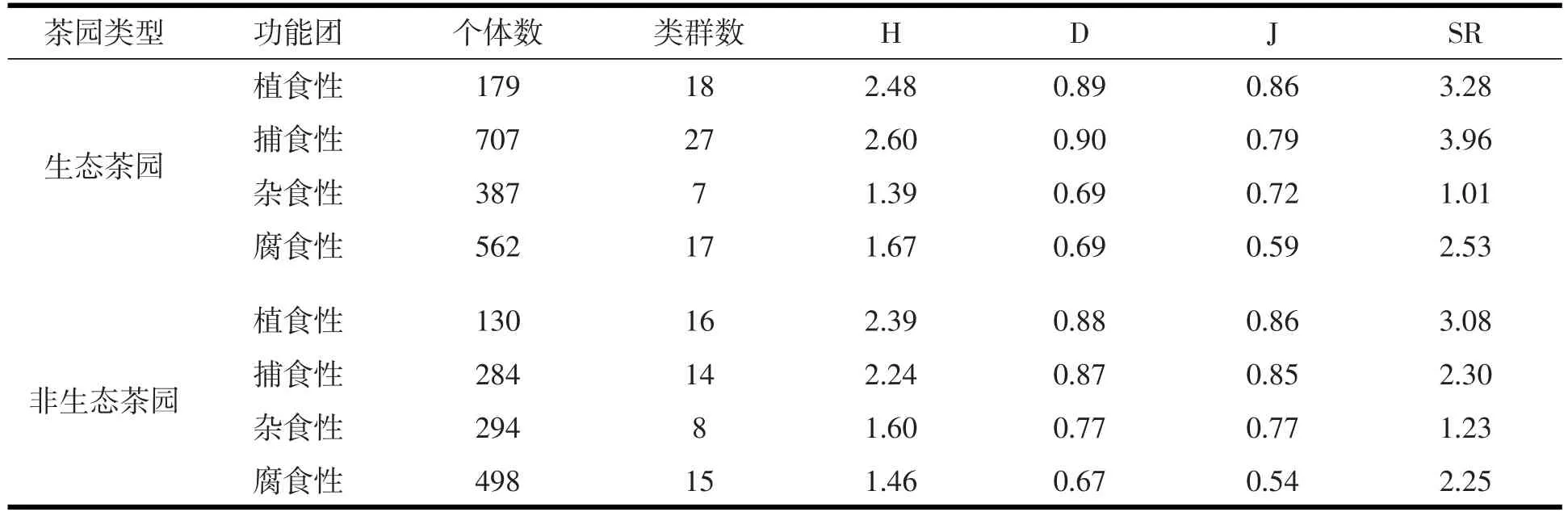
表4 不同茶园大型土壤动物功能类群多样性指数
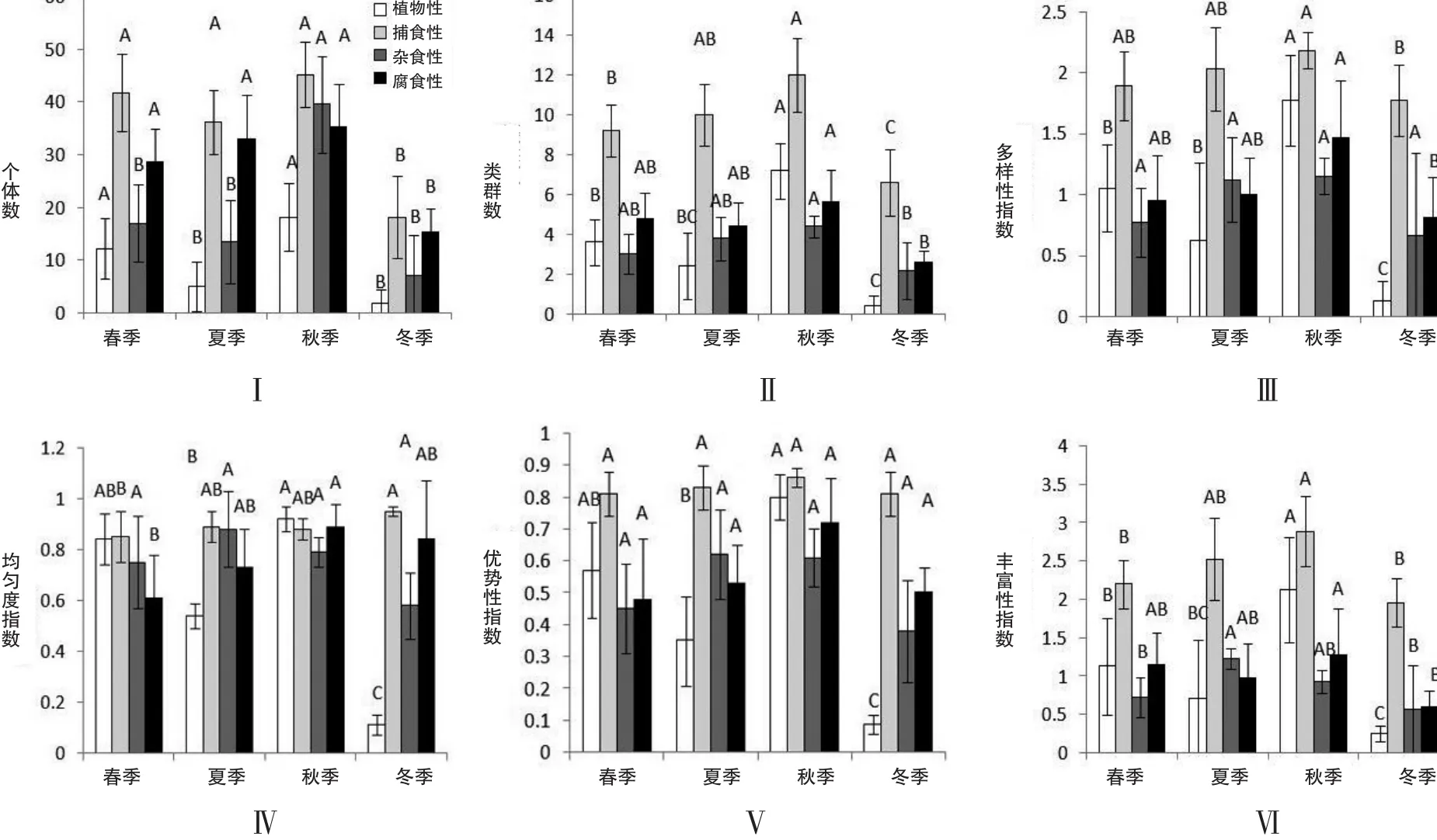
图2 生态茶园大型土壤动物功能类群季节动态
多样性指数分析,植食性动物类群冬季显著低于春季、夏季和秋季,秋季显著高于春季、夏季和冬季(P<0.05);捕食性和腐食性土壤动物类群秋季显著高于冬季(P<0.05),与春季和夏季无显著性差异;杂食性动物类群在四个季节无显著性差异(图2-Ⅲ).
均匀度指数分析,植食性动物类群冬季显著低于春季、夏季和秋季;捕食性动物类群冬季显著高于春季,夏、秋季间无显著性差异;杂食性动物类群四个季节均无显著性差异;腐食性动物类群的秋季显著高于春季(P<0.05),与夏季和冬季无显著性差异(图2-Ⅳ).
优势性指数分析,植食性土壤动物类群的优势性指数冬季显著低于春季、夏季和秋季(P<0.05);杂食性动物类群在四个季节中无显著性差异;捕食性动物和腐食性动物类群在四个季节中无显著性差异(图2-Ⅴ).
丰富度指数分析,植食性动物类群秋季显著高于春季、夏季和冬季;捕食性动物类群秋季显著高于春季和冬季,与夏季无显著性差异;杂食性动物类群夏季显著高于春季和冬季;腐食性动物类群秋季显著高于冬季,与春季和夏季无显著性差异(图2-Ⅵ).
2.3.2 非生态茶园大型土壤动物功能类群的组成
个体数分析,植食性和杂食性土壤动物类群的个体数秋季显著高于冬季(P<0.05),与春季和夏季无显著性差异;捕食性动物类群中冬季显著低于春季和秋季(P<0.05);腐食性动物类群冬季显著低于夏季和秋季(P<0.05)图3-Ⅰ).
类群数分析,植食性动物的类群数冬季显著低于春季、夏季和秋季(P<0.05);捕食性动物类群冬季显著低于春季和秋季(P<0.05),与夏季无显著性差异;杂食性动物类群在四个季节中均无显著性差异;腐食性动物类群冬季显著低于春季和秋季(P<0.05),与夏季无显著性差异(图3-Ⅱ).
多样性指数分析,植食性动物类群冬季显著低于春季、夏季和秋季(P<0.05),春季、夏季和秋季无显著性差异;捕食性和腐食性土壤动物类群秋季显著高于冬季(P<0.05),与春季无显著性差异;杂食性动物类群四季无显著性差异(图3-Ⅲ).
均匀度指数分析,植食性动物类群表现为冬季显著低于春季和秋季(P<0.05),与夏季无显著性差异;捕食性动物类群春季显著低于夏季和秋季(P<0.05);杂食性和腐食性动物类群在四个季节中均无显著性差异(图3-Ⅳ).
优势性指数分析,植食性土壤动物类群冬季显著低于春季、夏季和秋季(P<0.05);杂食性动物类群在四个季节中无显著性差异;捕食性动物类群秋季显著高于冬季(P<0.05),与春季和夏季无显著性差异;腐食性土壤动物类群冬季显著低于春季和秋季,与夏季无显著性差异(图3-Ⅴ).
丰富度指数分析,植食性动物类群冬季显著低于春季、夏季和秋季(P<0.05),春、秋季间无显著性差异;捕食性和杂食性动物类群在四个季节中均无显著性差异;腐食性夏季和冬季都显著低于春季和冬季(P<0.05)(图3-Ⅵ).
综合分析两个茶园的多样性指数表明,生态茶园除了冬季植食性动物类群外,其余食性的动物类群在四个季节中的个体数均高于非生态茶园.生态茶园四个食性动物类群在夏季和秋季中的类群数、多样性指数和丰富度指数均高于非生态茶园,而在春季,除捕食性动物类群外,均低于非生态茶园;在冬季,植食性和杂食性动物类群均低于非生态茶园,其余两个食性反之.两茶园优势性指数在春季、秋季和冬季与类群数一致,而在夏季,除植食性动物类群外,生态茶园其余食性类群均高于非生态茶园.

图3 非生态茶园大型土壤动物功能类群季节动态
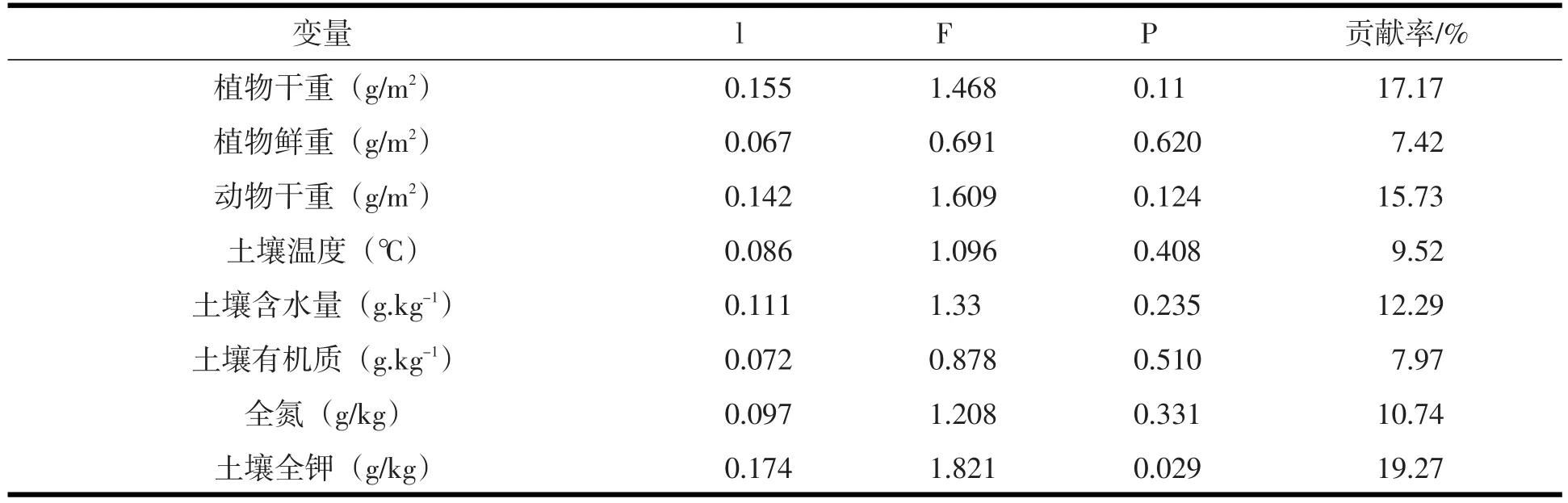
表5 环境因子对地面节肢动物群落变化的独立贡献
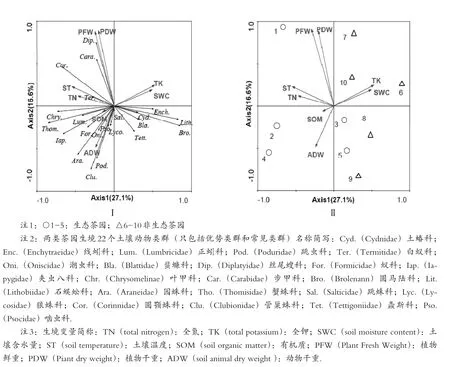
图4 生态茶园与非生态茶园土壤动物群落与2个环境变量关系的RDA二维排序图
3 生态茶园和非生态茶园大型土壤动物群落与环境因子的关系
图4 是对生态茶园和非生态茶园土壤动物群落的RDA排序分析.图4-Ⅱ可以看出所有非生态茶园在轴2的右侧,说明生态茶园和非生态茶园土壤动物群落存在明显的分布梯度.RDA排序图直观地反映了土壤动物群落与土壤环境因子的关系以及不同因子对动物类群分布的影响.图4-Ⅰ排序轴1的贡献率为27.1%,主要反映了全氮、全钾、土壤含水量、土壤温度对土壤动物群落分布的影响,其中,全钾和土壤含水量与轴1呈正相关,二者贡献率为31.56%(表5).全氮和土壤温度呈负相关.图4-Ⅰ排序轴2的贡献率为16.66%,主要反映植物鲜重、植物干重和动物干重对土壤动物群落分布的影响,轴2与茶园植物鲜重、植物干重呈正相关,与动物干重和土壤有机质呈负相关.RDA排序结果表明,两类茶园与土壤环境因子总贡献率为48.5%的土壤动物群落和环境因子的关系.
从排序图还可以看出,主要的动物类群分布明显受环境因子影响.丝尾螋科、步甲科受茶园植物量的影响,白蚁科和圆颚蛛科受土壤温度和全氮的影响;叶甲科、蟹蛛科、正蚓科、夹虫八科、蚁科、潮虫科、园蛛科、跳虫科、管巢蛛科、啮虫科、狼蛛科和跳蛛科受土壤有机质和动物干重的影响.

表6 生态茶园和非生态茶园群落相似性季节分析
4 生态茶园与非生态茶园群落相似性分析
相似性分析表明,生态茶园土壤动物群落夏季和秋季的相似性指数为0.51,属于中等相似,其它季节相似性指数在0.35~0.47之间,均表现为中等不相似.而非生态茶园各季节土壤动物群落的相似性指数在0.26~0.40之间,均为中等不相似,且低于生态茶园的相似性指数(夏冬季除外).
5 讨论
5.1 生态茶园与非生态茶园大型土壤动物的群落组成及多样性季节动态
生态茶园周围有毛竹、阔叶混交林和灌木,且伴生多种植物,植被相对茂盛,样地凋落物成分较复杂且较厚,大量的动物活跃在土壤表层,通过粉碎和分解的作用,将动植物的残体分解或转化为可被它们利用的物质,使大型土壤动物获得营养渠道更加广泛,具有更复杂的食物链.这种复杂的生境为大多数土壤节肢动物提供了捕食和生存的空间,吸引了诸如蜘蛛、甲虫和蜈蚣类等游猎性天敌,因此,生态茶园土壤动物多样性高,物种数多,个体数多,尤其是常见类群的丰度较高,为天敌提供充足的食物,使得生态茶园土壤动物群落较稳定,自然控制能力较强.而非生态茶园伴生植物少,周边生境遭到严重破坏,单一的植被使其土壤动物群落物种数量和个体数量均减少,多样性降低,结构不稳定.茶园类型不同,其环境、海拔高度、降雨量等存在很大差异,因此,温度、湿度和茶园及周边微环境是影响茶园土壤动物群落多样性季节变化的主要原因.两种茶园均受外界自然环境及气候因子的影响,导致同一类茶园土壤动物群落在不同季节的相似性较低.其中,非生态茶园植被组成单一,干扰程度大,土壤动物多样性较低,土地裸露面积较多,相对更易受季节性干旱和低温的影响,季节间群落特征波动较大,相似度指数相对比生态茶园的较低.
两茶园的常见类群和稀有类群均在秋季最高,冬季最低.这都受植被组成特征、环境的湿度和温度等多种因素的影响[21].在四个季节中,两茶园秋季的含水量低于春季和夏季,但高于冬季.冬季含水量不足将影响土壤动物的生存,春季和夏季含水量过高也会影响其生存.四个季节土壤动物的优势类群均为蚯蚓类,季节的变化对其影响不大.秋季凋落物层较厚,通过微生物的作用,形成腐殖质返还给土壤,为土壤动物提供丰富的营养来源.而且秋季温度和降雨量都比较适合土壤动物的生存和活动,会使其各种食性动物的个体数和类群数增加,冬季最低.
全年中气温和降雨量变化所引起的土壤温湿度的变化是影响大型土壤动物群落季节动态的主导因素[22].大量研究表明大型土壤动物具有明显的季节变化[23-26].生态茶园各个季节的大型土壤动物个体数、类群数、Shannon指数、Simpson指数、丰富度指数普遍高于非生态茶园,这与茶园的环境条件及管理方式有一定的联系.因为茶叶生产过程中的施肥、施用农药、植保、修剪、采摘等农事活动也会造成茶园土壤动物群落的个体数、物种数减少,多样性降低[5],进而影响土壤动物群落结构的稳定性.尤其是茶园土壤动物多样性的大小与人为采茶活动强度有较大相关性,采茶频率低,土壤环境相对稳定,采茶频率相对高,对土壤环境干扰强,使土壤动物多样性降低.生态茶园不施用农药和除草剂,每年采茶1次,平均每两年施有机肥1次,使土壤动物不受大的干扰.而非生态茶园施用农药高达10次以上,除草剂1~2次,每年采茶8次以上,施化肥2次,这些频繁的人为活动严重干扰土壤动物群落结构的稳定性.
5.2 生态茶园和非生态茶园大型土壤动物功能团组成及季节动态
茶园优越的生态环境为大型土壤动物直接提供食物,同时也培养了大量的土壤微生物,这些微生物被中小型动物所食.生态茶园在各个季节中捕食性动物是主要功能类群,而在非生态茶园中腐食性动物是其主要功能类群,这与茶园的环境与管理方式密切相关.生态茶园的植被较为丰富,物种数多,食物链比较复杂,因而捕食性动物较多.捕食性动物是土壤群落中的消费者,主要以小的动物和细菌为食物.生态茶园的优势类群中,步甲、蜘蛛类(狼蛛科、管巢蛛科、跳蛛科、球蛛科)和蜈蚣类数量居多.腐食性动物是作为分解者的角色,在生态系统物质转换和能量传递中起着重要的作用,而蚯蚓是茶园腐食性动物中活动能力最强的优势种,这一类动物主要是以有机物的碎屑为食物,主要起疏松土壤、粉碎动植物残体和提高土壤通气透水性等作用.而且蚯蚓作为优势类群已经比较适应该地区的环境,环境因素的变化对其的影响比起其他的食性的动物来说较小,因为人为的干扰导致非生态茶园土壤有机质的增加,为腐食性动物提供了生存条件.因此在非生态茶园中腐食性动物占据较高的优势.生态茶园良好的生态环境不仅为捕食性动物提供食物,也为植食性动物提供食物来源和栖息环境.
研究表明,影响土壤动物群落的因素很多,其中食物资源的多寡和环境条件的改变是影响其类群丰富度、多样性的主要因素[27].生态茶园的地表落叶较厚,通过微生物的作用,形成腐殖质返还给土壤,成为土壤动物的营养来源,而且凋落物又可以保住水分,对土壤肥力的恢复也有一定的好处.而且受到的人工干扰比非生态茶园的小,物种数较多,有较复杂的食物链,因此各种食性动物的个体数和物种数会增加.杂食性动物在茶园土壤生态系统中,其兼有多种功能群,即取食动物和植物,涉及的动物类群较多.虽然生态茶园杂食性动物各指数低于非生态茶园,但是生态茶园周边毛竹、阔叶混交林及灌木,二者之间的土壤动物有一定程度的流动,土壤生态系统的复杂性、稳定性和多样性对茶园土壤动物群落结构及稳定性具有很强的调节功能.
生态茶园海拔较高,冬季气温降低,干燥缺水及植被枯萎落叶等因素,导致两种茶园四个功能类群的动物数量均有所减少,但总体表现由于周边生境的影响,生态茶园各功能团在各季节中的个体数均高于非生态茶园(冬季植食性除外).生态茶园土壤动物的四个功能团类群,夏季和秋季的类群数、多样性指数和丰富度指数均高于非生态茶园;而春季的植食性动物、杂食性动物和腐食性动物类群的各指数均低于非生态茶园;冬季植食性和杂食性动物类群各指数均低于非生态茶园,捕食性和腐食性动物类群表现为生态茶园高于非生态茶园.
6 结论
季节气候是引起土壤动物群落多样性季节性动态变化的重要因素.广东属于亚热带地区,秋季温度和湿度均适宜土壤动物的生存,因此在该季节两个茶园土壤动物的个体数、物种数和多样性指数高.生态茶园夏季和秋季的四个功能团类群个体数和类群数高于非生态茶园.春季持续的降雨冲刷,对部分土壤动物生存的空间环境造成严重破坏,动物群落的个体数量和物种数会减少,其多样性指数低于夏、秋季节.茶园周边生境是维持茶园土壤动物群落结构稳定性和较高多样性的重要因子.生态茶园周边为竹林、阔叶林和灌木杂草等复杂的生境,为土壤动物的生存提供了良好的食物资源、繁殖与生存环境,从而导致生态茶园土壤动物个体数和物种数多,群落多样性高;而非生态茶园周边环境遭到严重破坏,食物资源较少,一部分土壤动物丧失了栖息环境,其繁殖和生存都受到很大影响,导致个体数和物种数减少,多样性降低,土壤动物群落结构遭到破坏而不稳定.因此,保护和建设良好的生态环境对茶树科学种植管理以及提高茶叶质量具有重要意义.
[1]周健民,石元亮.面向农业与环境的土壤科学[M].北京:科学出版社,2004:160-169.
[2]蒋海东,杨青,吕宪国.土壤动物在农业生态系统中的研究进展[J].土壤通报,2006,4:805-808.
[3]廖崇惠,李健雄,杨悦屏,等.海南尖峰岭热带林土壤动物群落——群落结构的季节变化及其气候因素[J].生态学报,2003,23(1):139-147.
[4]殷秀琴,张桂荣.森林凋落物与大型土壤动物相关关系的研究[J].应用生态学报,1993,4(2):167-173.
[5]王振中,张友梅,邢协加.土壤环境变化对土壤动物群落影响的研究[J].土壤学报,2002,39(6):892-898.
[6]朱永恒,濮励杰,赵春雨,等.我国土壤动物群落生态学研究综述[J].生态学杂志,2005,24(12):1477-1481.
[7]吴东辉,胡克.大型土壤动物在鞍山市大孤山铁矿废弃地生态环境恢复与重建中的指示作用[J].吉林大学学报:地球科学版,2003,33(2):213-216.
[8]王移,卫伟,杨兴中,等.我国土壤动物与土壤环境要素相互关系研究进展[J].应用生态学报,2010,21(9):2441-2448.
[9]朱立安,魏秀国.土壤动物群落研究进展[J].生态科学,2007,26(3):269-273.
[10]庞学勇,刘庆,刘世全,等.人为干扰对川西亚高山针叶林土壤物理性质的影响[J].应用与环境生物学报,2002,8(6):583-587.
[11]李涛,刘苑秋,郭圣茂,等.瑞昌石灰岩红壤区退耕还林土壤动物群落特征[J].应用生态学报,2012,23(4):910-916.
[12]勾影波,高方清,冯婷婷.常熟虞山茶园土壤动物群落结构的多样性研究[J].中国农学通报,2012,28(26):201-205.
[13]申燕,郑子成,李廷轩.茶园土壤动物群落特征及其与土壤理化特性的关系[J].浙江大学学报:农业与生命科学版,2010,36(5):503-512.
[14]尹文英.中国亚热带土壤动物[M].北京:科学出版社,1992:83-56.
[15]尹文英.中国土壤动物[M].北京:科学出版社,2000.
[16]郑乐怡,归鸿.昆虫分类(上、下)[M].南京:南京师范大学出版社,1999:27-975.
[17]周尧.周尧昆虫图集[M].郑州:河南科学技术出版社,2002:30-544.
[18]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000.
[19]孙儒泳,李庆芬,牛翠娟.基础生态学[M].北京:高等教育出版社,2002:142-148.
[20]马克平,钱迎倩.生物多样性研究的原理与方法[M].北京:中国科学技术出社,1994:141-165.
[21]刘继亮,殷秀琴,邱丽丽.左家自然保护区大型土壤动物与土壤因子关系研究[J].土壤学报,2008,45(1):130-136.
[22]杨效东,唐建维.西双版纳不同演替状态热带次生林土壤节肢动物群落特征[J].应用生态学报,2004,15(6):988-994.
[23]张秀娟,苏永春,杨晨利.洞庭湖环湖丘岗生态系统大型土壤动物群落结构特征分析[J].华中农业大学学报,2005,24(6):633-637.
[24]王海霞,殷秀琴,周道玮.松嫩草原区农牧林复合系统大型土壤动物生态学研究[J].草业学报,2003,12(4):84-89.
[25]刘新民,刘永江,郭砺.内蒙古典型草原大型土壤动物群落动态及其在放牧下的变化[J].草地学报,1999,7(3):228-235.
[26]殷秀琴,吴东辉,韩晓梅.小兴安岭森林土壤动物群落多样性的研究[J].地理科学,2003,23(3):316-322.
[27]范月君,侯向阳,石红霄,等.气候变暖对草地生态系统碳循环的影响[J].草业学报,2012,21(3):294-302.
The Large Soil Animal Community and Seasonal Dynamics in Tea Plantations with Various Surrounding Habitats
LI Jin-lin,OU Li-li,HUANG Zhen-hui,HUANG Li-fang,WEI Shao-na,XU Jia-ming,XING Shu-wen*
(College of Food engineering and Biotechnology,Hanshan Normal University,Chaozhou,Guangdong,521041)
Plantations in Fenghuang mountain and Pingxi town were surveyed by using the ecological method,the composition and the dynamic differences of two types of soil animal community and nutrient group were analyzed.Moreover,we discussed the factors affecting the community structure and dynamic changes of soil animals in the two kinds of tea plantations.The results indicated that a total of 1 835 soil macrofauna individuals was captured and classified into 9 classes,22 orders and 69 families in the ecological tea plantation,accounting for 60.34%of the total catch of soil macrofauna.A total of 1 026 soil macrofauna individuals was captured and classified into 9 classes,19 orders and 53 families in the non-ecological tea plantation,accounting for 39.66%of the total catch of soil macrofauna.Individual number of every species in ecological tea plantation was higher than the ecological tea plantation.The dominant groups of the ecological tea plantation soil animal in summer,autumn and winter were lower than the non-ecological tea garden,and the common groups were higher than those of the non-ecological tea plantation.The number,variety,Shannon index and Simpson index of soil animal were higher in autumn and spring,low in winter.According to the analysis of soil animal in functional group diversity and composition,the predatory animals of ecological tea plantation were the main functional groups,which accounted for 38.53%of the total amount.The saprophagous soil fauna of the non-ecological tea plantation was the main functional groups,accounting for 41.29%of the total amount.In summer and autumn,the number,variety,Shannon index and richness index of each functional group in the ecological tea plantation were higher than the non-ecological tea plantation.The number,species and diversity index of predatory animals in spring and winter were higher than the non-ecological tea plantation,and herbivorous and omnivorous animals were lower than the non-ecological tea plantation.RDA sorting results showed that obvious distributing gradient existed between ecological and non-ecological tea gardens,which indicated the distribution of soil animals was affected by environmental factors,such as TN,TK,organic matter,moisture,temperature and vegetation of soil.To sum up,the surrounding environment of tea plantation is an important factor affecting the community structure of soil animals,and the major factor that affects the seasonal dynamic distribution of soil animals is climate.
ecological tea plantation;non-ecological tea plantation;soil macro-fauna;community;seasonal dynamic;functional group
Q 958.1
A
1007-6883(2017)03-0041-12
责任编辑周春娟
2016-12-16
广东大学生科技创新培育专项资金(项目编号:pdjh2016b0320);广东省教育厅特色创新项目(项目编号:2014KTSCX160);潮州市科技计划项目(项目编号:2014N09);广东省协同创新与平台环境建设专项资金项目(项目编号:2014A070713039).
李金林(1993-),女,广东揭阳人,韩山师范学院食品工程与生物科技学院2012级学生.邢树文为通讯作者.
猜你喜欢
当代水产(2021年10期)2022-01-12
生态学报(2021年3期)2021-03-31
心声歌刊(2021年6期)2021-02-16
江淮法治(2020年16期)2020-11-27
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
上海包装(2019年2期)2019-05-20
乡村地理(2018年1期)2018-07-06
百科知识(2018年7期)2018-04-17
广东农业科学(2017年5期)2017-08-29
