
4种化学调控剂对烟草幼苗耐弱光性及光合特性的影响
2018-04-08 01:53杨利云王丽特徐照丽杨双龙
西北植物学报 2018年1期
杨利云,王丽特,徐照丽,杨双龙,何 彬,龚 明*
(1 云南师范大学 生命科学学院,生物能源持续开发利用教育部工程研究中心,云南省生物质能与环境生物技术重点实验室,昆明650500;2 云南省烟草农业科学研究院,昆明650031;3 云南省烟草烟叶公司,昆明650218)
光环境作为一个极其重要的生态因素,可通过光强、光质、光周期三个途径影响植物的地理分布、生长发育及物质代谢等过程,植物可以通过长期进化过程中形成的光感受和应答系统,调节光形态建成、生理代谢途径、基因表达水平等方式,以适应周围光环境的变化[1]。近年来,随着全球气候变暖的趋势,常常出现气候异常,局部地区长时间阴雨天气,造成植物生长在弱光环境下。此外,在一些设施农业中,弱光胁迫已成为影响作物生长发育、产量及品质的一个重要限制因素。前人研究表明,不同光照强度对植物的地理分布、形态建成、生长发育及物质代谢等均有显著的影响,相同的光照强度对不同植物或处于不同生长时期相同植物的影响也不同[2-3]。
烟草(NicotianatabacumL.)作为一种重要的经济作物和科研模式植物被广泛的种植和深入研究。我们研究组近年来的研究结果表明,作为一种喜光植物,烟草的生长对周围光环境的变化较为敏感,不同的光质、光强及光照时间对烟草植株的生长发育及物质代谢均有显著影响,进而影响烟叶的产量和品质[4-10]。前人研究也表明,不同光照强度对烟草的种子萌发、生长发育、生理生化过程、物质代谢途径、烟叶的产量和品质[11-13]有显著的影响。在实际生产中,大棚或温室育苗及长期阴雨天气等因素容易引起弱光环境,导致烟株的生长发育减缓,生育期延长、干物质积累减少、植株生长纤弱、叶片变薄、各种生理生化及物质代谢过程受到影响,进而影响烟叶的产量和品质[14]。因此,通过各种农艺措施来提高烟草植株对弱光胁迫的耐受性有重要的理论和实践意义。
钙是植物生长发育过程中所必需的一种大量元素,细胞外Ca2+不仅可以稳定和保护细胞质膜结构和功能,同时还对植物体内其它重要生理生化代谢过程起到良性作用;此外,Ca2+作为植物细胞中最重要的第二信使,参与植物对许多逆境信号的转导,当植物生长遭遇各种逆境胁迫时,胞质内Ca2+在时间、空间及浓度变化上呈现特异性变化,即诱导产生钙信号,然后再经下游的钙结合蛋白进行感受和转导,进而引起胞内一系列生理生化反应,从而对各种逆境胁迫进行响应,提高植物的抗逆性[15]。赤霉素(GAs)是植物中一类重要的四环二萜类化合物,目前已知有136种天然赤霉素广泛分布于细菌、真菌、藻类及植物中,但仅GA1、 GA3、GA4、GA7有生理活性,它们在植物整个生长发育过程中发挥着重要的调节作用;同时,GAs还作为一类信号分子,在植物应对逆境胁迫的响应过程中发挥着重要的调控功能[16]。水杨酸(SA)是植物中广泛存在的一种简单的酚类化合物,其在植物生热、开花、侧芽萌发、性别分化、成熟衰老、系统抗性的获得等生长发育等过程中发挥着重要的作用,同时SA也以一种信号分子的形式参与植物对各类逆境胁迫的响应[17]。多效唑(PP333)又名氯丁唑,是一种高效、低毒的植物生长调节剂,具有抑制茎枝伸长,促进茎围增加、植株分蘖、成花和坐果、延缓植物生长和衰老,进而提高植物抗逆性的功能[18]。
利用各种细胞信号分子、植物激素及生长调节剂对植物正常生长及逆境胁迫环境下生长发育进行调控具有广泛的应用前景及意义,国内外学者对此已进行了很多的研究及报道。已有学者分别研究了Ca2+、GAs、SA和PP333预处理对一些植物在低温、高温、干旱、盐胁迫等逆境下生长发育及生理特性的影响[19-21]。最近,我们研究组也对这4种化学调控剂预处理对烟草幼苗耐冷性的影响进行了报道[22]。弱光胁迫也是植物生长发育过程中经常遭遇到的一种非生物逆境胁迫,但目前有关这4种化学调控剂预处理对增强植物耐弱光性的效应还未见报道,特别是4种化学调控剂中,哪一种调控剂、多高的浓度处理对植物的耐弱光性综合效果最佳,目前也还不清楚。因此,本研究通过对烟草幼苗喷施不同浓度的4种化学调控剂进行预处理,然后于人工气候室内模拟弱光环境处理,并对弱光环境下烟草幼苗的生长、伤害指标及叶片光合性能进行分析,并应用隶属函数法对这4种化学调控剂预处理对改善烟苗品质及缓解弱光胁迫的效应进行综合评价,以期为促进烟草及其它作物壮苗及耐弱光性提供指导和借鉴。
1 材料和方法
1.1 材料及处理
试验以烟草品种‘云烟97’为材料,种子由云南省玉溪中烟种子有限公司提供,按GB/T25241.1-2010方法在温室内培育烟苗。待种子萌发,烟苗长至3叶1心时,分别于温室育苗池内对每个处理的烟苗喷施不同浓度的CaCl2、GA3、SA、PP333,以蒸馏水为对照,共9个处理(表1)。每个处理共计336株烟苗,每隔2 d 喷施1次,共喷施4次,以全株淋湿、药液欲滴为标准,对照喷施相同体积的蒸馏水。喷施完第4次后,继续置于育苗池内生长2 d,然后将烟苗移入人工气候室中育苗池内进行弱光(光照强度:80 μmol·m-2·s-1)处理,光周期14 h /10 h,相对湿度70%,CO2浓度400 μl/L,变温生长,日均温23.5 ℃[23],分别于弱光处理0、3、6 d取样测定相关指标。
1.2 测试指标与方法
1.2.1生长指标分别于弱光处理的0、3、6 d选取每个处理中长势均匀的烟苗30株,分为3组,每组10株,洗净并自然风干,用吸水纸吸取表面残留水滴,用电子天平分别对每个处理烟株的整株、地上部及地下部分鲜重进行测定,然后于烘箱内杀青后烘至恒重并分别测定其干重,并计算相对生长速率和干物质积累速率[22]。
相对生长速率(RGR)=(lnW2-lnW1)/(t2-t1)
干物质积累速率=(t2天后的干物质量-t1天后的干物质量)/(t2-t1)
式中,W1、W2分别表示在时间t1和t2时的干重。
1.2.2生理生化指标分别于弱光处理时0、3、6 d时,选取不同处理的烟苗自下向上第6片烟叶为材料,混匀称量并置于液氮中保存,采用硫代巴比妥酸(TCA)法测定不同处理烟叶内的丙二醛(MDA)含量[24]。按王学奎[25]的方法测定烟叶叶绿素含量、根系活力(TTC法)和脯氨酸(酸性茚三酮比色法)含量,采用相对电导法测定电解质渗透率[26]。
1.2.3光合参数根据柯学等[4]的方法,使用LI-6400XT(Li-COR, USA)于弱光处理0、3、6 d时分别测定不同处理烟苗自下向上第6片烟叶的净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr)。
1.3 数据处理
数据来源于3个重复小区,结果采用平均值 ± 标准差(mean±sd)表示,利用SPSS 15.0 进行单因素方差分析,差异显著时用LSD法进行多重比较,P<0.05表示显著性差异,使用Sigmplot 12.0 作图。
另外,利用隶属函数法[27]对4种化学调控剂预处理烟草幼苗生长发育(0 d)及弱光胁迫(6 d)时的耐弱光性进行综合评价,每个处理各项指标的具体隶属函数值计算公式为:
Xu=(X-Xmin)/(Xmax-Xmin)(1)
Xu=1-(X-Xmin)/(Xmax-Xmin)(2)
式中,X为不同处理某一生理指标的测定值,Xmax、Xmin分别为所有处理中该指标的最大值和最小值,若所测指标与植物的耐弱光性呈正相关则采用(1)式计算隶属值,负相关则用(2)式计算。最后把每个处理各项指标隶属函数值累加,取平均值,平均值越大,耐弱光性越强,反之,耐弱光性越弱。
2 结果与分析
2.1 化学调控剂预处理对弱光胁迫下烟草幼苗生长的影响
首先,在弱光胁迫期内,各处理烟苗株高均随处理时间延长呈逐渐增加的趋势;在弱光胁迫0、3和6 d时,P1和P2预处理烟苗株高均比CK显著降低,其余处理烟苗株高均比CK不同程度增加,且除S2外大多达到显著水平;各处理间相比较,各时期始终以G1、G2显著较高,C2和S1次之,C1和S2较低,P1和P2最低。其中,弱光胁迫处理6 d时, G1、G2、C2、S1处理株高分别比CK显著增加87.2%、65%、32.9%和32.5%,C1、S2预处理与CK相近,而P1和P2预处理则分别比CK显著降低42.8%和34.6%(图1,A)。
同时,弱光胁迫0 d时,仅C1、S2和P2预处理烟苗单株干重比CK显著增加,其余处理与CK无显著差异;而弱光胁迫6 d后,各处理烟苗单株干重均比弱光胁迫前(0 d)不同程度降低,但各浓度化学调控剂预处理烟苗均比同期CK显著增加,增幅在20.4%~40.1%之间,但仍以S2和P2处理较高(图1,B)。此外,经弱光胁迫处理6 d后,各浓度化学调控剂预处理及对照烟苗的相对生长速率及干物质积累速率均有所降低且为负值;但与CK相比,4种化学调控剂预处理均显著减缓了弱光胁迫下烟苗相对生长速率和干物质积累速率的降低幅度,并均以S1、P1处理效果最为明显(图1,C、D)。以上结果表明,弱光胁迫致使烟叶的光合作用同化量低于烟苗整体(根、茎、叶)呼吸作用所消耗的物质量,显著抑制了烟苗正常生长,而4种化学调控剂预处理可有效缓解弱光胁迫对烟苗生长的抑制。
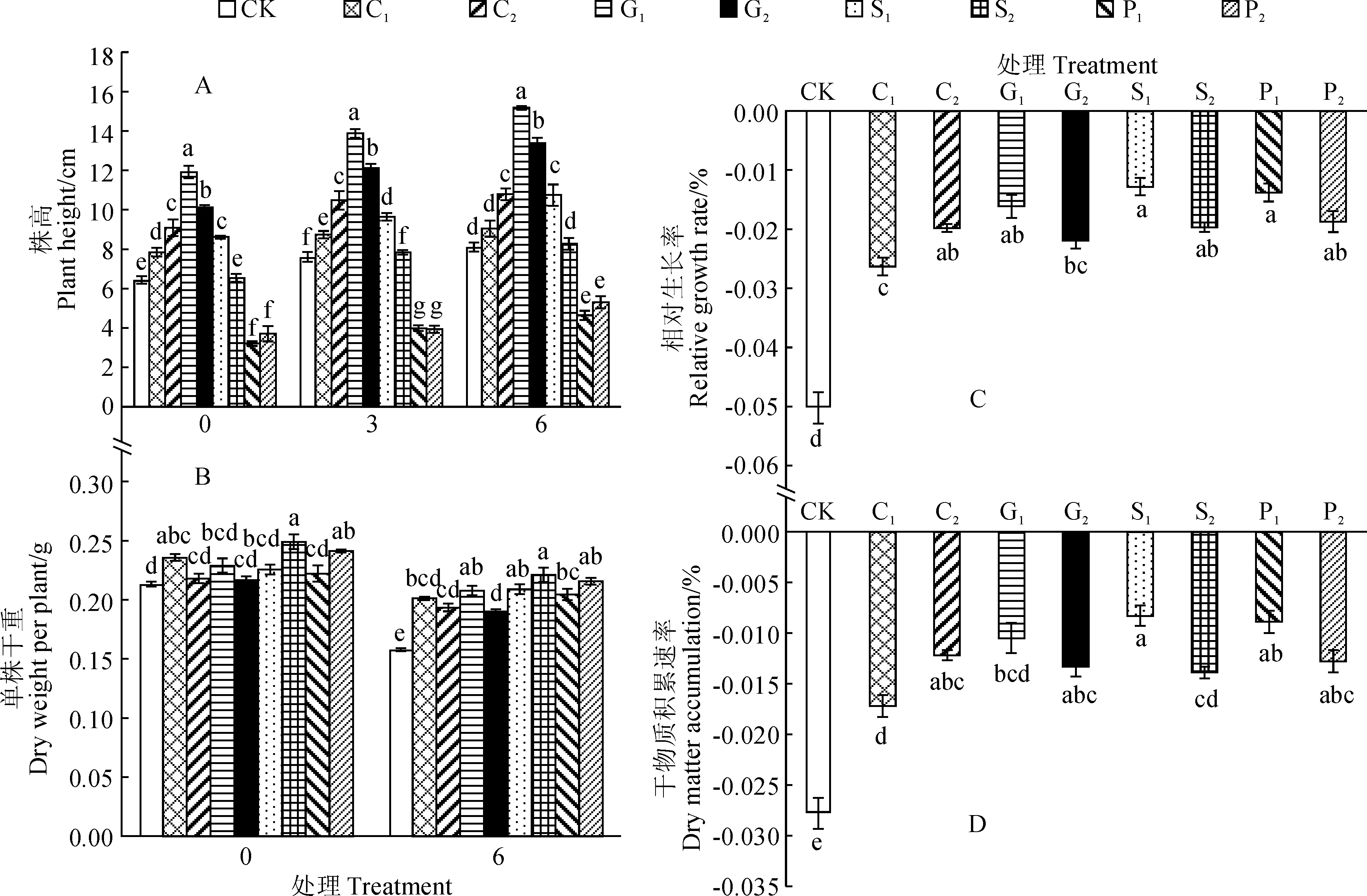
处理代码同表1;同期不同小写字母表示处理间差异显著(P<0.05);下同图1 弱光胁迫下4种化学调控剂预处理烟草幼苗生长的变化Treatment codes in figures were same as in Table 1; The different normal letters within the same period indicates significant difference (P<0.05) among treatments; The same as belowFig.1 The growth changes of tobacco seedlings pretreated with four chemical regulators under low light stress

图2 弱光胁迫下4种化学调控剂预处理烟草幼苗根系活力及总叶绿素含量的变化Fig.2 The root vigor and total chlorophyll content of tobacco seedlings pretreated with four chemical regulators under low light stress
2.2 化学调控剂预处理对弱光胁迫下烟草幼苗根系活力及叶绿素含量的影响
随着弱光胁迫时间的延长,各处理烟苗根系活力呈逐渐降低的趋势,而其烟叶总叶绿素含量则呈逐渐升高的趋势(图2,A、B)。弱光胁迫0~6 d时,各浓度化学调控剂预处理烟苗根系活力均比同期CK不同程度增加,其中C1、S1、S2及P1处理增幅在各时期均达到显著水平,它们在弱光胁迫处理6 d时分别比CK显著增加47.8%、131.4%、62%和95%(图2,A)。同时,各浓度化学调控剂预处理烟苗叶片叶绿素含量在弱光胁迫0~6 d期间均比同期CK显著增加,它们在预处理6 d时的增幅在37.4%~77.3%之间,且S1处理含量在各时期始终处于显著较高水平(图2,B)。以上结果说明4种化学调控剂预处理可有效缓解弱光胁迫下烟苗根系活力的降低程度并提高弱光胁迫下烟叶内总叶绿素含量,进而提高烟苗的耐弱光性。
2.3 化学调控剂预处理对弱光胁迫下烟草幼苗电解质渗透率、MDA及脯氨酸含量的影响
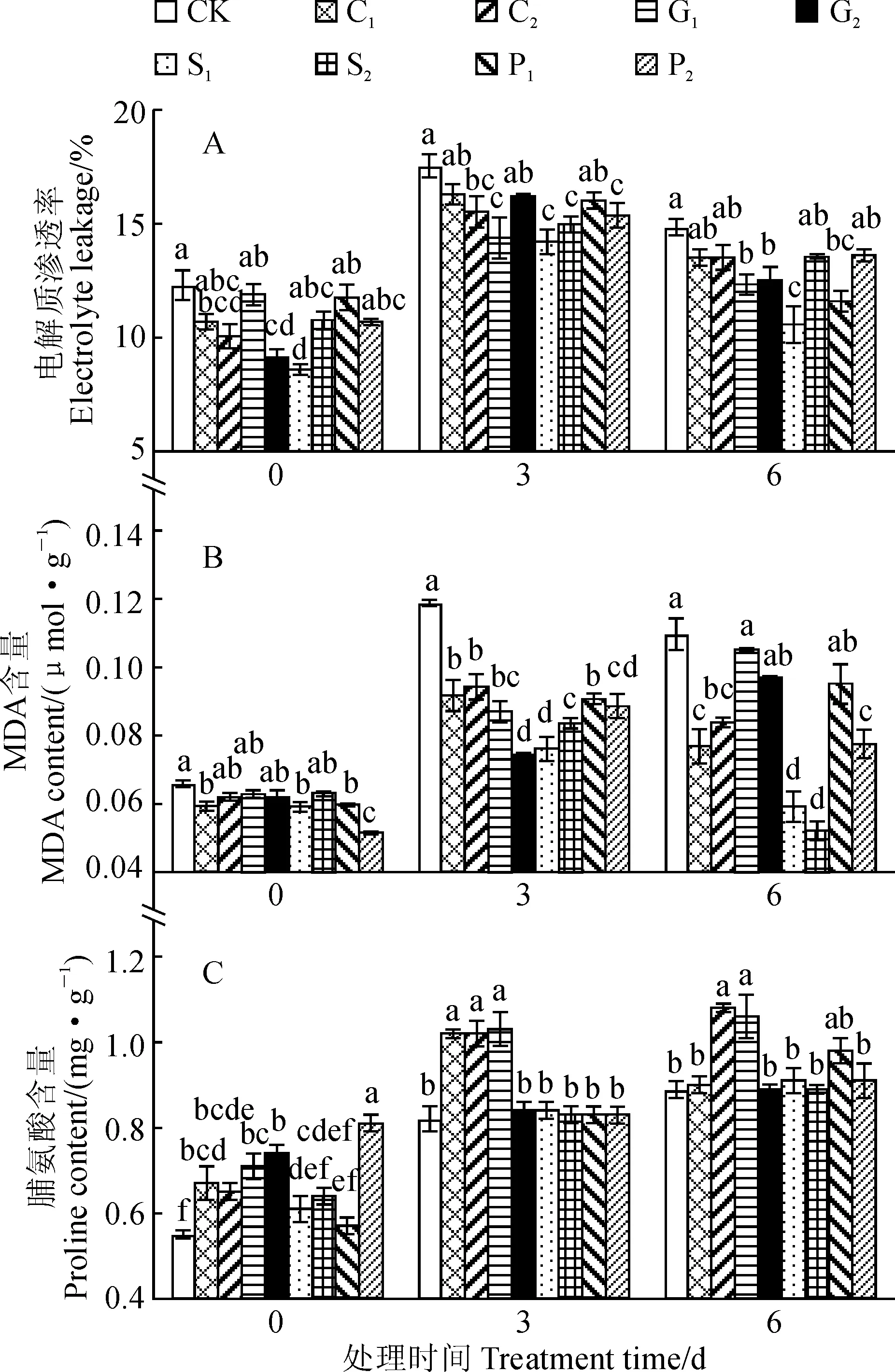
图3 弱光胁迫下4种化学调控剂预处理烟草幼苗电解质渗透率、MDA及脯氨酸含量的变化Fig.3 The electrolyte leakage, MDA and proline contents of tobacco seedlings pretreated with four chemical regulators under low light stress
图3显示,随着弱光胁迫时间的延长,各处理烟草幼苗叶片电解质渗透率、MDA及脯氨酸含量均呈现出先升后降的变化趋势,均在胁迫3 d时达到最大值;同时,与同期CK 相比,各浓度化学调控剂预处理幼苗电解质渗透率和MDA含量均不同程度降低,而它们的脯氨酸含量均不同程度增加。其中,弱光胁迫0 d的C2、G2、S1处理和弱光胁迫3~6 d的G1、S1处理烟苗叶片电解质渗透率均比同期CK显著降低(图3,A);弱光胁迫0 d时C1、S1、P1、P2处理,胁迫3 d时所有处理以及胁迫6 d时C1、C2、S1、S2、P2处理烟草叶片内MDA含量均显著低于同期CK(图3,B);弱光胁迫0 d时C1、C2及G1、G2、P2处理,以及弱光胁迫3~6 d时C2和G1预处理烟叶内脯氨酸的含量均比CK显著增加(图3,C)。以上结果说明4种化学调控剂预处理可有效缓解弱光胁迫下烟叶内电解质渗透率的增加、降低烟叶内MDA水平,减小弱光胁迫对烟苗细胞膜的损伤,并在一定程度上促进烟叶内脯氨酸积累,进而提高烟苗的耐弱光性。
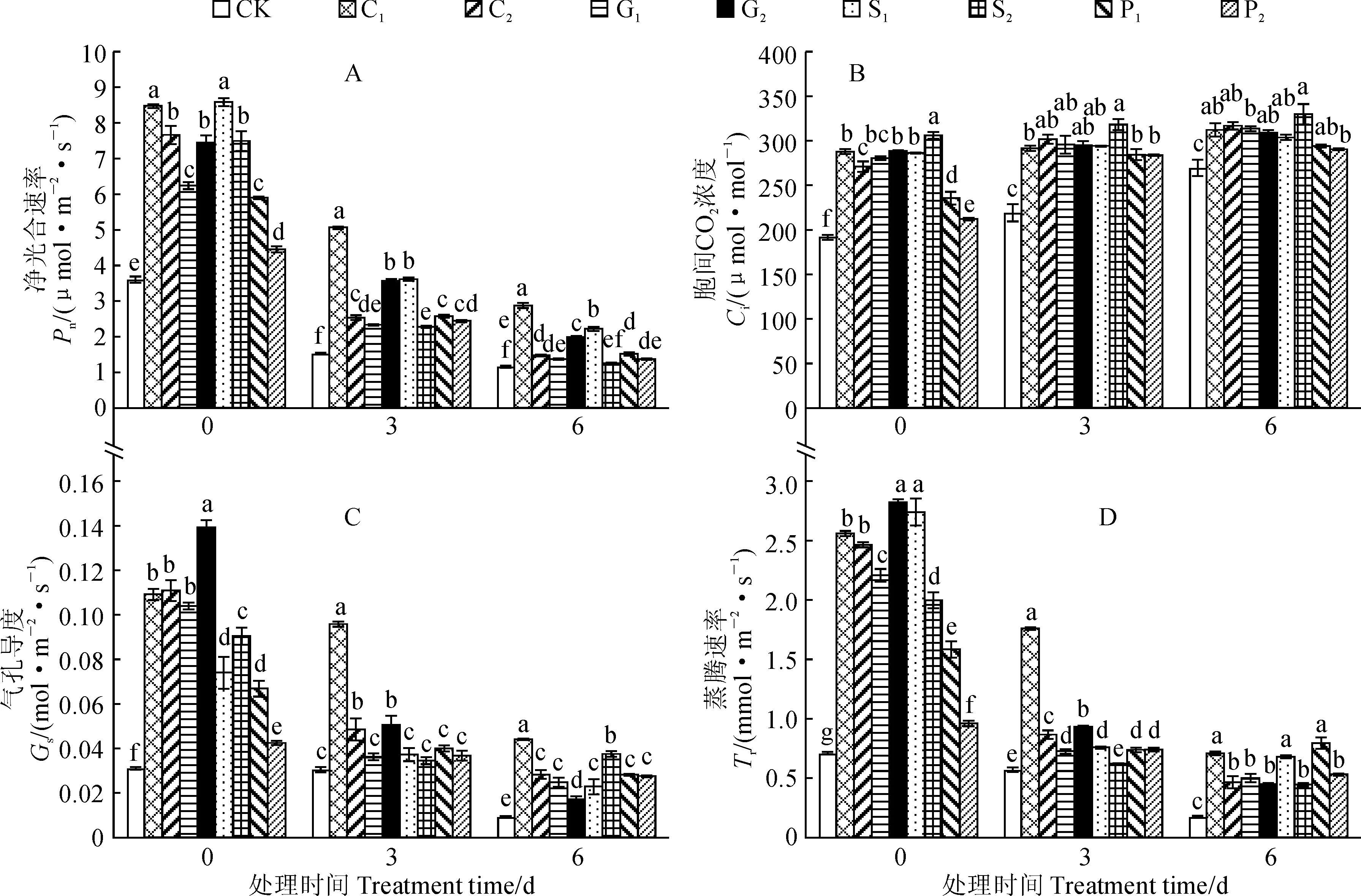
图4 弱光胁迫下4种化学调控剂预处理烟草幼苗叶片光合参数的变化Fig.4 The photosynthetic characteristics of tobacco seedlings pretreated with four chemical regulators under low light stress
2.4 化学调控剂预处理对弱光胁迫下烟草幼苗叶片光合性能的影响
图4显示,随着弱光胁迫时间的延长,各处理烟叶的净光合速率(Pn)、气孔导度(Gs)及蒸腾速率(Tr)均逐渐明显降低,而其胞间CO2浓度(Ci)却逐渐增加;与对照相比较,在弱光胁迫0~6 d内,烟苗叶片的Pn、Gs、Ci及Tr整体都不同程度增加,且增幅大多达到显著水平(P<0.05)。综合来看,弱光胁迫明显抑制了各处理烟苗叶片光合作用的正常进行,而不同化学调控剂预处理均可有效缓解弱光胁迫对光合作用的抑制。
2.5 化学调控剂预处理对弱光胁迫下烟草幼苗耐弱光性影响的综合评价
采用隶属函数法分别对4种化学调控剂喷施后及弱光胁迫6 d后烟草幼苗的6个主要生理指标进行全面、综合的评价,比较不同处理对烟草幼苗耐弱光性影响的大小。隶属函数分析结果(表2)表明,弱光胁迫前(0 d),4种化学调控剂处理对烟苗生长发育的促进效果表现为S1> S2> C1> C2> P2> G2> P1>G1,综合来看SA(S1、S2)和CaCl2(C1、C2)处理效果要优于GA3(G1、G2)和PP2333(P1、P2)处理;而在弱光胁迫6 d后,4种化学调控剂处理对烟草幼苗耐弱光性促进效果为 S1> C1> S2> P1> P2> G2> G1>C2,也是以SA和CaCl2处理的效果最佳(表2)。因此,化学调控剂SA和CaCl2在促进烟草幼苗生长和耐弱光性方面的表现比GA3和PP2333更佳。
3 讨 论
烟草作为一种喜光作物,其种子萌发、植株生长发育、烟叶内物质代谢、烟叶的产量、品质及风味等均受到光照强度的广泛调控[28-30]。前人研究证实,与正常光照强度下生长的烟草相比,弱光环境下生长的烟草株高和叶片总叶绿素含量显著增加,而相对生长速率、干物质积累速率、单株干重和根系活力及叶片光合性能则显著降低;此外,弱光环境引起烟草植株内各种代谢途径紊乱失衡,加速活性氧的爆发和积累,加剧细胞膜损伤,使烟草叶片的电解质渗透率和MDA含量升高[31-36]。本研究结果也发现,弱光(80 μmol·m-2·s-1)胁迫6 d后,烟草幼苗株高增加,而单株干重、相对生长速率、干物质积累速率、根系活力降低,同时烟叶内电解质渗透率和MDA含量显著增加。这表明弱光环境使烟草植株的同化作用降低,表现为对烟草植株生长的抑制;同时弱光环境加速了烟草植株的细胞膜损伤,若烟草长期生长于较低光强下,将进一步加速烟草植株的衰老和死亡,严重影响烟草的产量及品质。此外,本研究还发现,与喷施蒸馏水相比,4种化学调控剂预处理提高了弱光胁迫前烟草幼苗的单株干重、根系活力、烟叶总叶绿素、脯氨酸含量及光合性能,提高了烟苗素质。经隶属函数综合评价分析,4种化学调控剂处理对弱光胁迫前烟草幼苗的壮苗综合效果为S1>S2>C1>C2>P2>G2>P1>G1;整体上,SA(S1、S2)和CaCl2(C1、C2)处理对改善烟苗品质的效果优于GA(G1、G2)和PP333(P1、P2)处理。4种化学调控剂预处理均在一定程度上提高了烟苗的整体素质,这为烟苗后期缓解弱光胁迫提供了一个稳定的物质基础和前提。
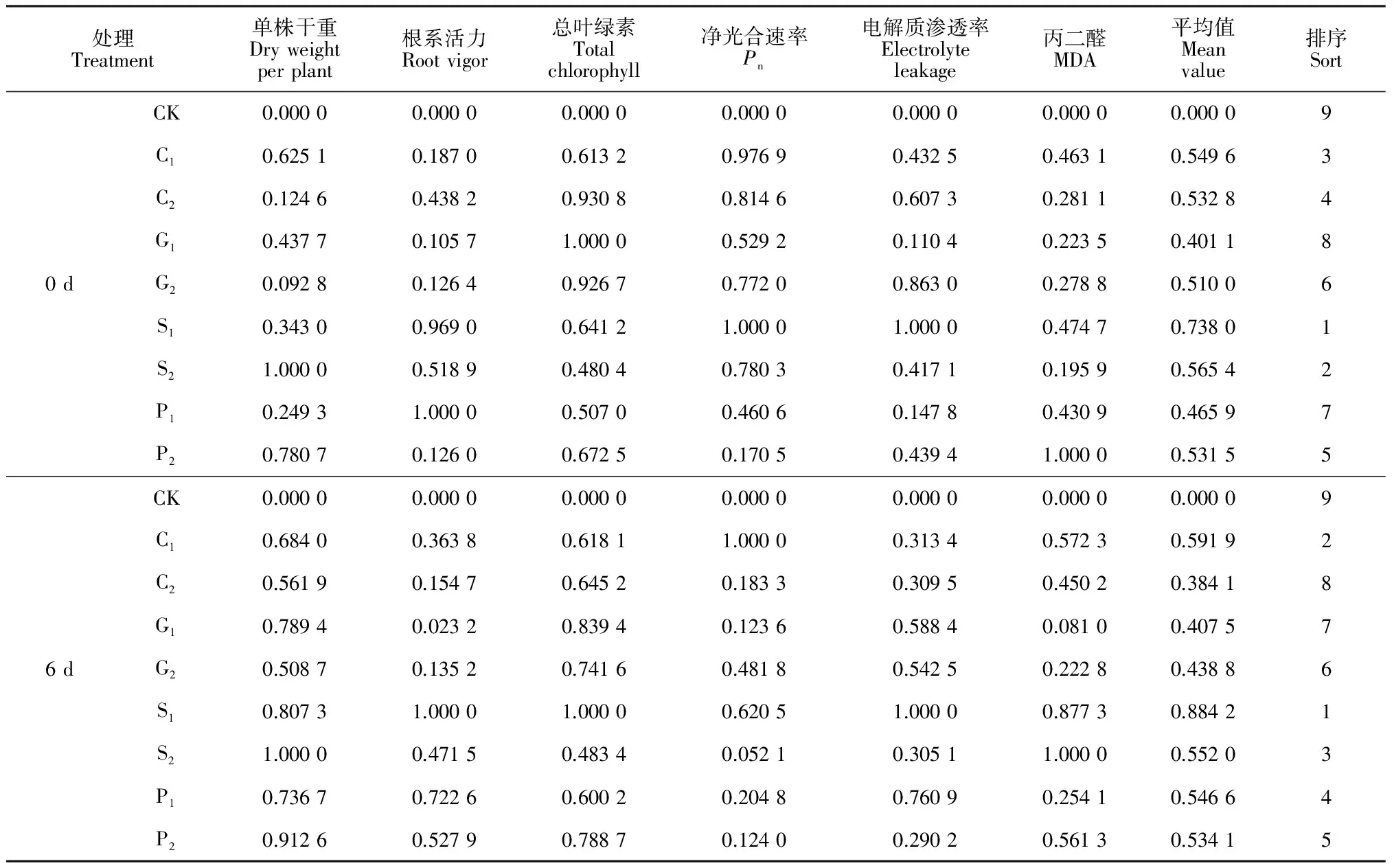
表2 4种化学调控剂预处理对烟草幼苗耐弱光性的综合评价
干物质的积累是植物生长发育的物质前提和基础,前人研究证实,在弱光环境下烟草植株的单株干重、相对生长速率、干物质积累速率、根系活力降低[35-36],本研究中也观察到了相同的结果。虽然经弱光胁迫6 d后,烟苗单株干重与0 d时相比降低,但经4种化学调控剂预处理均显著延缓了弱光胁迫下烟苗相对生长速率和干物质积累速率的降低程度,并使烟苗单株干重显著高于对照,进而增强了烟苗在弱光胁迫下的耐弱光性。经隶属函数分析表明,4种化学调控剂预处理对烟草幼苗单株干重的促进作用表现为S2>P2>S1>G1>P1>C1>C2>G2>CK,并以2.0 mmol/L SA(S2)处理效果最佳,这与我们之前有关4种化学调控剂预处理促进低温胁迫下烟苗的干物质积累的研究结果相似[22]。
在各类逆境胁迫下,植物体内各种代谢途径紊乱失衡,活性氧爆发,造成氧化胁迫及细胞膜损伤,引起细胞内电解质渗漏,膜脂过氧化作用加剧,表现为电解质渗透率和MDA含量的升高,二者含量越高表明膜损伤程度越大[37]。本研究中,弱光胁迫6 d后烟苗叶片电解质渗透率及MDA含量与0 d时相比均显著增加,表明弱光胁迫加剧了细胞膜的过氧化损伤程度,质膜结构遭到破坏。但经4种化学调控剂预处理后,弱光下烟苗的电解质渗透率及MDA含量与CK相比均显著下降,说明4种化学调控剂处理可有效缓解弱光胁迫对烟苗叶片的膜脂过氧化作用,对细胞膜具有保护和修护功能,并以SA(1.0 和2.0 mmol/L)的处理效果最佳。这与王丽特等[22]的研究结果一致。
叶绿素是植物光合作用过程中光能转换和传递的物质基础,反映了植物生长的内在动力,在一定范围内,植物体内的叶绿素含量升高,植物的光合性能也相应增加,促进植物的生长[38]。本研究中,弱光胁迫6 d 后,烟苗叶片内的总叶绿素含量显著增加,且经4种化学调控剂预处理后,其总叶绿素含量显著高于CK,经隶属函数分析,4种化学调控剂处理对烟叶总叶绿素提高的效果为S1> G1> P2> G2> C2> C1> P1> S2> CK,这在一定程度上促进弱光胁迫下烟苗叶片光合性能的提高,进而增强烟苗的耐弱光性,其中1.0 mmol/L SA(S1)的处理效果最好。
绿色植物通过将光能转化为化学能,同时将无机物转变为有机物贮存在植物体内,是地球上所有生命现象的能量和物质基础[23]。随着光照强度的减弱,烟草植株的光合性能降低,同化作用减弱,干物质积累减少,生长受到抑制[35]。本研究中,经弱光胁迫6 d后,烟苗叶片的光合性能降低,但经4种化学调控剂预处理后,烟苗叶片的光合性能均显著提高且大于CK,在一定程度上促进了烟叶的光合作用,延缓了弱光下烟苗干物质积累的降低,并以3.0 mmol/L CaCl2(C1)处理的效果最佳,1.0 mmol/L SA(S1)处理的次之。前人研究表明,外源施用CaCl2可显著提高弱光环境下黄瓜幼苗叶片的光合酶基因表达水平及光合酶活性,增强弱光下黄瓜幼苗的光合性能,提高叶片叶绿素及脯氨酸含量及抗氧化水平,增强黄瓜幼苗在弱光胁迫环境下的适应性[39-41]。类似的研究中也证实,SA预处理可显著提高弱光下黄瓜幼苗叶片的抗氧化性、提高光合酶活性及基因表达水平,增强黄瓜幼苗的光合性能[42-44]。这很可能也是本研究中CaCl2和SA预处理提高弱光下烟苗叶绿素含量,增强抗氧化水平从而降低膜损伤并提高烟叶光合性能,促进同化作用,进而提高烟苗耐弱光性的主要原因之一。目前针对GA和PP333处理对植物耐弱光性的影响还没有相关的报道,在未来的研究中还需要深入探究。
大量研究也证实,外源施用CaCl2、GA3、SA及PP3334种化学调控剂可通过提高植物在正常生长环境和各类非生物逆境胁迫(干旱、高温、低温、盐胁迫等)下的抗氧化水平,降低植物在逆境胁迫下的氧化损伤,维持细胞正常的运转及功能,提高植物的光合性能,增强植物的渗透调节能力,进而促进植物的生长和提高植物的抗逆性[45-48],这很可能也是本研究中4种化学调控剂预处理提高烟苗耐弱光性的作用机制之一。
整体来看,与喷施蒸馏水(CK)相比,4种化学调控剂预处理均可在一定程度上提高烟苗的单株干重、根系活力、烟叶总叶绿素含量及光合性能,并降低烟叶MDA含量及电解质渗透率,改善烟苗的素质,其壮苗效果从大到小依次为:S1> S2> C1> C2> P2> G2> P1> G1。而在弱光胁迫6 d后,与CK相比,4种化学调控剂预处理在一定程度上提高了弱光胁迫下烟苗的单株干重、根系活力、烟叶总叶绿素及其光合性能,同时降低了烟苗电解质渗透率、MDA水平,延缓了烟苗相对生长速率及干物质积累速率的降低,进而增强了弱光胁迫下烟苗的耐弱光性,各处理对缓解烟草幼苗弱光胁迫的效应从强到弱分别为:S1> C1> S2> P1> P2> G2> G1> C2。综合来看,以1.0 mmol/L SA预处理效果最佳,这与低浓度的SA预处理具有提各种非生物逆境胁迫下植物细胞的抗氧化水平,维持细胞结构完整及正常运转,加强胁迫信号转导及响应过程,进而提高植物应对各类非生物逆境胁迫的适应性和抗逆性有密切关系[49]。
参考文献:
[1]FRANKLIN K A. Light and temperature signal crosstalk in plant development[J].CurrentOpinionPlantBiology, 2009,12(1): 63-68.
[3]FAN X X, XU Z G, LIU X Y,etal. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light[J].ScientiaHorticulturae, 2013,153(153): 50-55.
[4]柯学, 李军营, 李向阳,等. 不同光质对烟草叶片生长及光合作用的影响[J]. 植物生理学报, 2011,47(5): 512-520.
KE X, LI J Y, LI X Y,etal. Effects of different light quality on growth and photosynthesis of tobacco (NicotianatabacumL.) leaves[J].PlantPhysiologyJournal, 2011,47 (5): 512-520.
[5]柯学, 李军营, 徐超华, 等. 不同光质对烟草叶片组织结构及Rubisco羧化酶活性和rbc、rca基因表达的影响[J]. 植物生理学报, 2012,48(3): 251-259.
KE X, LI J Y, XU C H,etal. Effects of different light quality on anatomical structure, carboxylase activity of ribulose 1,5-Biphosphate carboxylase oxygenase and expression ofrbcandrcagenes in tobacco (NicotianatobacumL.) leaves[J].PlantPhysiologyJournal, 2012,48(3): 251-259.
[6]文锦芬, 柯学, 徐超华, 等. 光质对烟草叶片生长发育过程中抗氧化系统的影响[J]. 西北植物学报, 2011,31(9): 1 799-1 804.
WEN J F, KE X, XU C H,etal. Effects of light quality on antioxidant defense system during growth and development of tobacco leaves[J].ActaBotanicaBoreali-OccidentaliaSinica, 2011,31(9): 1 799-1 804.
[7]ZHAO J, XU C H, LI J Y,etal. Effects of different light qualities on activity and gene expression of caspase-like proteases in tobacco leaves[J].AgriculturalScienceandTechnology, 2012,13(2): 276-279.
[8]徐超华, 李军营, 崔明昆, 等. 延长光照时间对烟草叶片生长发育及光合特性的影响[J]. 西北植物学报, 2013,33(4): 763-770.
XU C H, LI J Y, CUI M K,etal. Effects of supplemental lighting on growth and photosynthesis of tobacco leaves[J].ActaBotanicaBoreali-OccidentaliaSinica, 2013,33(4): 763-770.
[9]杨利云, 李军营, 王丽特,等. 光环境对烟草生长及物质代谢的影响研究进展[J]. 基因组学与应用生物学, 2015,34(5): 1 114-1 128.
YANG L Y, LI J Y, WANG L T,etal. Research advances about effects of light environment on growth and material metabolism of tobacco (NicotianatabacumL.) plants[J],GenomicsandAppliedBiology, 2015,34(5): 1 114-1 128.
[10]YANG L Y, WANG L T, MA J H,etal. Effects of light quality on growth and development, photosynthetic characteristics and content of carbohydrates in tobacco (NicotianatabacumL.) plants[J].Photosynthetica, 2017,55(3): 467-477.
[11]SEIBERT M, WETHERBEE P J, JOB D D. The effects of light intensity and spectral quality on growth and shoot initiation in tobacco callus[J].PlantPhysiology, 1975,56(1):130-139.
[12]MIYAKE C, HORIGUCHI S,etal. Effects of light intensity on cyclic electron flow around PSI and its relationship to non-photochemical quenching of Chl fluorescence in tobacco leaves[J].PlantandCellPhysiology, 2005,46(11): 1 819-1 830.
[13]WALTER A, NAGEL K A. Root growth reacts rapidly and more pronounced than shoot growth towards increasing light intensity in tobacco seedlings[J].PlantSignalBehavior, 2006,1(5): 225-226.
[14]杨兴友,崔树毅, 刘国顺,等.弱光环境对烟草生长、生理特性和品质的影响[J]. 中国生态农业学报, 2008,16(3): 635-639.
YANG X Y, CUI S Y, LIU G S,etal. Effect of weak light on the growth, physiology and quality of tobacco[J].ChineseJournalofEco-Agriculture, 2008,16(3): 635-639.
[15]HONGBO S, LIYE C, MINGAN S,etal. Bioengineering plant resistance to abiotic stresses by the global calcium signal system [J].BiotechnologyAdvances, 2008,26(6): 503-510.
[16]COLEBROOK E H, THOMAS S G, PHILLIPS A L,etal. The role of gibberellin signalling in plant responses to abiotic stress[J].JournalofExperimentalBiology, 2014,217: 67-75.
[17]SAHU. Salicylic acid: Role in plant physiology and stress tolerance[J],MolecularStressPhysiologyofPlants, 2013, 217-239.
[18]GHOSH S, PENTERMAN J N, MADEY E,etal. Paclobutrazol: Its physiological effects and biochemical functions during plant stress[J].PhysiologyandMolecularBiologyofPlants, 2003,9(2): 163-173.
[19]LURIE S, RONEN R, LIPSKER Z,etal. Effects of paclobutrazol and chilling temperatures on lipids, antioxidants and ATPase activity of plasma membrane isolated from green bell pepper fruits[J].PhysiologiaPlantarum, 1994,91(4): 593-598.
[20]KNIGHT H, TREWAVAS A J, KNIGHT M R. Calcium signalling inArabidopsisthalianaresponding to drought and salinity[J].PlantJournal, 1997,12(5): 1 067-1 078.
[21]WANG L J, FAN L,etal. Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves[J].BMCPlantBiology, 2010,10(1): 1-10.
[22]王丽特, 徐照丽, 杨利云,等. 4种化学调控剂对烟草幼苗耐冷性及其光合特性的效应研究[J]. 西北植物学报, 2015,35(4): 801-808.
WANG L T, XU Z L, YANG L Y,etal. Effects of four chemical regulators on chilling tolerance and photosynthetic characteristics of tobacco seedlings[J].ActaBotanicaBoreali-OccidentaliaSinica, 2015,35(4): 801-808.
[23]杨利云, 段胜智, 李军营, 等. 不同温度对烟草生长发育及光合作用的影响[J]. 西北植物学报, 2017,37(2): 330-338.
YANG L Y, DUAN S Z, LI J Y,etal. Effects of different temperatures on growth, development and photosynthesis of tobacco (NicotianatabacumL.) plants[J].BotanicaBoreali-OccidentaliaSinica, 2017,37(2): 330-338.
[24]赵世杰, 李德全. 植物生理学实验指导[M]. 北京: 科学出版社, 2004: 305-306.
[25]王学奎. 植物生理生化实验原理与技术[M]. 北京: 高等教育出版社, 2006,5: 134-136.
[26]贾开志, 陈贵林. 高温胁迫下不同茄子品种幼苗耐热性研究[J]. 生态学杂志, 2005,24(4): 398-401.
JIA K Z, CHEN G L. Tolerance of different eggplant varieties at seedling stage to high temperature stress[J].ChineseJournalofEcology, 2005,24(4): 398-401.
[27]郭郁频, 任永霞, 刘贵河, 等. 外源钙和赤霉素对干旱胁迫下苜蓿幼苗生理特性的影响[J]. 草业学报, 2015,24(7): 89-96.
GUO Y P, REN Y X, LIU G H,etal. Effects of calcium (CaCl2), GA3and complex liquid on physiological characteristics of alfalfa seedlings under drought[J].ActaPrataculturaeSinica, 2015,24(7): 89-96.
[28]ANDERSEN R, KASPERBAUER M J. Chemical composition of tobacco leaves altered by near ultraviolet and intensity of visible light[J],PlantPhysiology, 1973,51: 723-726.
[30]杨兴有, 刘国顺. 成熟期光强对烤烟理化特性和致香成分含量的影响[J]. 生态学报, 2007,27(8): 3 450-3 456.
YANG X Y, LIU G S. Effects of different light intensity during tobacco ripe stages on the quality of flue-cured tobacco[J].ActaEcologicaSinica, 2007,27(8): 3 450-3 456.
[31]杨兴有, 叶协锋, 刘国顺, 等. 光强对烟草幼苗形态和生理指标的影响[J]. 应用生态学报, 2007,18(11): 2 642-2 645.
YANG X Y, YE X F, LIU G S,etal. Effects of light intensity and physiological characteristics of tobacco seedlings[J].ChineseJournalofAppliedEcology, 2007,18(11): 2 642-2 645.
[32]刘国顺, 乔新荣, 王芳, 等. 弱光对烤烟干物质积累及矿质养分的影响[J]. 西南农业学报, 2008,21(1): 130-133.
LIU G S, QIAO X R, WANG F,etal. Effects of low light on dry matter and mineral of flue-cured tobacco[J].SouthwestChinaJournalofAgriculturalSciences, 2008,21(1): 130-133.
[33]刘典三, 刘国顺, 贾芳芳, 等. 不同光强对烤烟质体色素及其降解产物的影响 [J]. 华北农学报, 2013,28(1): 234-238.
LIU D S, LIU G S, JIA F F,etal. Effects of light intensity on plastid pigment and its degraded products in flue-cured tobacco[J].ActaAgriculturaeBoreali-Sinica, 2013,28(1): 234-238.
[34]乔新荣, 郭桥燕, 刘国顺, 等. 光强对烤烟生长发育及光合特性的影响[J]. 华北农学报, 2007,22(3): 76-79.
QIAO X R, GUO Q Y, LIU G S,etal. Effects of light intensity on growth and photosynthetic characteristics on flue-cured tobacco[J].ActaAgriculturaeBoreali-Sinica, 2007,22(3): 76-79.
[35]王瑞, 刘国顺, 陈国华,等. 光强对苗期烤烟光合作用及干物质生产的影响[J]. 应用生态学报, 2010,21(8): 2 072-2 077.
WANG R, LIU G S, CHEN G H,etal. Effects of light intensity on photosynthesis and dry matter production of flue-cured tobacco at its seedling stage[J].ChineseJournalofAppliedEcology, 2010,21(8): 2 072-2 077.
[36]魏明月, 云菲, 刘国顺,等. 不同光环境下烟草光合特性及同化产物的积累与分配机制[J]. 应用生态学报, 2017,28(1): 159-168.
WEI M Y, YUN F, LIU G S,etal. Response of photosynthetic characteristics and accumulation and distribution of assimilation products in tobacco to different light environments[J].ChineseJournalofAppliedEcology, 2017,28(1): 159-168.
[37]SUZUKI N, KOUSSEVITZKY S, MITTLER R,etal. ROS and redox signaling in the responses of plants to abiotic stress[J].PlantCellandEnvironment, 2012,35: 259-270.
[38]王竞红,多多。多效唑对6种草坪草苗期抗旱性影响的研究[J]. 草业学报, 2014,23(6): 253-258.
WANG J H, DUO D. Effects of paclobutrazol on drought resistance of six turfgrass cultivars during the seedlings stage[J].ActaPrataculturaeSinica, 2014,23(6): 253-258.
[39]杨俊兴, 郭庆军, 韩雁, 等. 水分胁迫条件下钙赤合剂和磷浸种对冬小麦幼苗净光合速率和抗氧化系统的影响[J]. 作物杂志, 2012,4: 25-29.
YANG j X, GUO Q J, HAN Y,etal. Effects of seed soaking with Ca-GA mixture and phosphorus on photosynthetic rate and antioxidant system of wheat seedlings under water stress condition[J].Crops, 2012,4: 25-29.
[40]齐红岩, 王丹, 齐明芳, 等. 不同形态钙对高温逆境下番茄叶片光合作用的调控作用[J]. 应用生态学报, 2014,25(12): 3 540-3 546.
QI H Y, WANG D, QI M F,etal. Regulation of different calcium forms on the photosynthesis of tomato leaves under heat stress[J].ChineseJournalofAppliedEcology, 2014,25(12): 3 540-3 546.
[41]齐明芳, 刘玉凤, 周龙发, 等. 钙对亚高温下番茄幼苗叶片光合作用的调控作用[J]. 中国农业科学, 2011,44(3): 531-537.
QI M F, LIU Y F, ZHOU L F,etal. Regulation of calcium on photosynthesis of tomato leaves under sub-high temperature stress[J].ScientiaAgriculturalSinica, 2011,44(3): 531-537.
[42]毕焕改,董旭兵,王美玲,等. 钙和水杨酸对亚适温弱光下黄瓜幼苗光合酶活性结合基因表达的影响[J]. 园艺学报, 2015,42(1): 56-64.
BI H G, DONG X B, WANG M L,etal. Foliar spray calcium and salicylic acid improve the activities and gene expression of photosynthetic enzymes in cucumber seedlings under low light intensity and suboptimal temperature[J].ActaHorticulturaeSinica, 2015,42(1): 56-64.
[43]刘伟, 艾希珍, 梁文娟, 等. 低温弱光下水杨酸对黄瓜幼苗光合作用及抗氧化酶活性的影响[J]. 应用生态学报, 2009,20(2): 441-445.
LIU W, AI X Z, LIANG W J,etal. Effects of salicylic acid on leaf photosynthesis and antioxidant enzyme activities of cucumber seedlings under low temperature and light intensity[J].ChineseJournalofAppliedEcology, 2009,20(2): 441-445.
[44]WANG H, LIU G S, YUN F,etal. Effects of salicylic acid on photosynthesis and chlorophyll fluorescence characteristics of flue-cured tobacco leaf in subdued light[J].AgriculturalScienceandTechnology, 2014,15(1): 33-38.
[45]XU C B, LI X M,etal. The effect of calcium chloride on growth, photosynthesis, and antioxidant responses ofZoysiajaponicaunder drought conditions[J].PlosOne, 2013,8(7): e68214
[46]LIANG W J, WANG M L, AI X Z. The role of calcium in regulating photosynthesis and related physiological indexes of cucumber seedlings under low light intensity and suboptimal temperature stress[J].ScientiaHorticulturae, 2013,123: 34-38.
[47]WANG H, LIU G S, YUN F,etal. Effects of salicylic acid on photosynthesis and fluorescence characteristics of flue-cured tobacco leaf in subdued light[J].AgriculturalScienceandTechnology, 2014,15(1): 33-38.
[48]BANINASAB B, GHOBADI C. Influence of paclobutrazol and application methods on high-temperature stress injury in cucumber seedlings[J].JournalofPlantGrowthRegulation, 2011,30: 213-219.
[49]HORVATH E, SZALAI G, JANDA T. Induction of abiotic stress tolerance by salicylic acid signaling[J].JournalofPlantGrowthRegulation, 2007,26: 290-300.
猜你喜欢
江西通信科技(2022年2期)2022-08-08
温州大学学报(自然科学版)(2022年2期)2022-05-30
现代农业科技(2022年1期)2022-01-17
潍坊学院学报(2020年2期)2021-01-18
数字通信世界(2019年10期)2019-02-14
现代园艺(2017年23期)2018-01-18
制导与引信(2017年3期)2017-11-02
西南农业学报(2017年4期)2017-05-17
海军航空大学学报(2015年4期)2015-02-27
中国烟草科学(2014年3期)2014-05-25
