
慢病毒介导的脯氨酰寡肽酶过表达对大鼠肝星状细胞功能的影响
2018-05-03 01:43何玲楠李梦婷陈源文范建高
胃肠病学和肝病学杂志 2018年4期
周 达, 王 晶, 何玲楠, 李梦婷, 孙 超, 陈源文, 范建高
上海交通大学医学院附属新华医院消化内科,上海 200092
脯氨酰寡肽酶(prolyloligopeptidase,POP,EC 3.4.21.26)是一种能特异性水解短肽中脯氨酸残基羧基端肽键的丝氨酸蛋白酶,在体内发挥重要生理功能,参与多肽的生成与代谢、调控细胞增殖与分化、调节体内炎症代谢等[1-3];但是,POP在外周组织中的具体生物学作用仍未知。肝脏中POP活性很高,提示其在肝内生物学功能值得探索[4-6]。同时已有研究[7]发现,POP参与肝内炎症及肝细胞增殖与分化。我们前期实验发现,POP水解胸腺素β4(thymosin β4,Tβ4)产物N-乙酰基-丝氨酸-天冬氨酸-赖氨酸-脯氨酸(N-acetyl-seryl-aspartyl-lysyl-proline,Ac-SDKP)具有抑制肝星状细胞(hepatic stellate cells,HSC)活性和抗肝纤维化作用[8-9];其他研究人员发现,Tβ4对四氯化碳诱导急性肝毒性有保护作用同时可以抑制HSC活性[10-11]。然而,POP在HSC内具体功能尚不明确。
肝纤维化主要表现肝内炎症及细胞外基质沉积,HSC起至关重要的作用[12-13]。本研究旨在以HSC作为切入点,通过携带POP基因的慢病毒感染HSC-T6,探讨POP在HSC内的生物学功能,进一步阐明POP在肝脏炎症与纤维化发生、发展中的作用。
1 材料与方法
1.1细胞培养大鼠肝星状细胞系HSC-T6细胞购自中国科学院上海生命科学研究院细胞资源中心。HSC-T6根据需要以适当密度接种于不同大小培养皿中,培养液为质量浓度为100 g/L的FBS/DMEM培养基(血清及培养基均购自Gibco,USA),置于含体积分数为5%的CO2孵育箱中,在细胞处于良好生长状态时进行下述相关实验。
1.2携带POP基因的慢病毒感染HSC通过PubMed搜索大鼠POP基因(Prep,NM_031324),设计携带POP和绿色荧光蛋白(GFP)基因的慢病毒(POP-virus)及空载病毒(Mock-virus),并由上海吉凯基因化学技术有限公司合成;选取不同的病毒感染复数(multiplicity of infection,MOI值)1、10、100进行感染预实验,荧光显微镜下观察感染效率,最终将MOI确定为10;接种一定数量HSC-T6至3.5 cm培养皿24 h后,弃去旧培养液,随后参照产品说明书进行感染操作,加入慢病毒或空病毒感染12 h后换液为质量浓度为100 g/L的FBS/DMEM培养基继续培养48~72 h(根据细胞密度决定),同时设立正常对照组,荧光显微镜下观察感染效率,随后进行各组细胞RNA及蛋白提取及后续实验。
1.3ELISA检测对POP慢病毒组、空病毒组及正常对照组进行细胞内Ac-SDKP测定,采用ELISA测定方法,按试剂盒说明书(购自SPI Bio and CEA,France)进行。Ac-SDKP的终浓度通过各组细胞数目进行校正,计算出每106个细胞中Ac-SDKP的量,以(nmol/L)/106表示。
1.4CCK-8细胞增殖毒性检测将POP慢病毒、空病毒及正常的HSC-T6以相同数量接种于96孔板中共培养24 h 和48 h,每孔加入200 μl培养液,每孔加入100 μl新鲜培养液,同时设立空白对照组,再加入10 μl CCK-8试剂,避光37 ℃孵育2 h,随后通过酶标仪(uQuant,Biotek,USA)测定每孔在450 nm处吸光度值(OD值)。各孔OD值均需先减去空白孔OD值后分析各组HSC-T6的增殖生长差异。
1.5细胞凋亡检测将感染慢病毒、空病毒及正常的HSC-T6以相同数量接种于3.5 cm培养皿中培养24 h,使用不含EDTA的胰酶(购自GIBCO,USA)消化提取细胞,然后通过流式细胞学检测HSC-T6凋亡,具体步骤参照Annexin V-PE/7-AAD凋亡试剂盒(Becton-Dickinson,USA)。
1.6实时定量PCR检测对POP慢病毒组、空病毒组及正常对照组进行HSC细胞内RNA提取,采用Trizol法(Trizol购自Takara)测定各组提取的RNA浓度及纯度,通过逆转录试剂盒逆转录为cDNA(primescriptRTmaster mix购自Takara),进而利用cDNA进行荧光定量实时定量PCR(SYBR Premix Ex Taq购自Takara,仪器为Biosystems 7500 Real-time PCR system)检测POP、转化生长因子-β1(TGF-β1)、α平滑肌肌动蛋白(α-SMA)、单核细胞趋化蛋白-1(MCP-1)、Ⅰ型胶原(Col I)相关基因mRNA表达量,各基因设置3个复孔,各基因引物由生工生物工程(上海)股份有限公司合成(见表1),最终结果使用相对表达量RQ值(2-ΔΔCt)表示,具体操作步骤参考实际说明书。
1.7Westernblotting检测提取POP慢病毒组、空病毒组及正常对照组细胞内蛋白,采用BCA法测定各组蛋白浓度(BCA试剂购自碧云天生物技术有限公司)。随后进行SDS-PAGE电泳,转膜,质量浓度为50 g/L的BSA封闭2 h,各相关蛋白一抗孵育(4 ℃过夜),1×TBST洗膜10~15 min×3次,二抗孵育1~2 h,1×TBST洗膜10~15 min×3次,最后使用辣根过氧化物酶法检测(购自Millipore Corporation,Billerica,USA),使用Image Lab分析蛋白显影条带。POP、TGF-β1、α-SMA三者抗体均购自Sigma-Aldrich公司,p-Smad2/3抗体购自巴傲得生物科技有限公司,Smad7、过氧化物酶体增殖物激活受体-γ(PPAR-γ)购自生工生物工程(上海)股份有限公司,相关二抗及Western blotting相关其他试剂购自碧云天生物技术有限公司,具体步骤参考相关试剂说明书。


表1 Real-time-PCR引物序列Tab 1 Sequence of primers of Real-time-PCR
注:*:Prep为POP的基因名称。
2 结果
2.1慢病毒感染效率48~72 h后进行荧光显微镜下GFP检测,慢病毒及空病毒感染效率均95%以上(见图1A),为进一步检测病毒感染HSC效率,检测HSC内POP的基因及蛋白表达量,慢病毒组POP mRNA和蛋白表达较空病毒组及正常对照组显著升高(见图1B~1C)。
2.2POP慢病毒感染对HSC内Ac-SDKP水平的影响POP慢病毒组细胞内Ac-SDKP较正常对照组及空病毒组显著升高约50%(P<0.05),空病毒组与正常对照组比较,差异无统计学意义(P>0.05)(见图2A)。
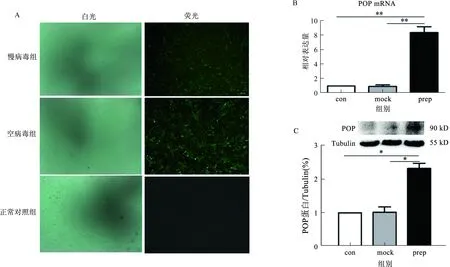
注:con:正常对照组;mock:空病毒组;prep:POP慢病毒组。*P<0.05;**P<0.01。
CCK-8检测发现,慢病毒组细胞在24 h及48 h均较正常对照组增殖生长快,在48 h时较空病毒组多增殖约50%,而空病毒组与正常对照组细胞增殖比较,差异无统计学意义(P>0.05)(见图2B)。不同组别HSC-T6相同条件下培养24 h后,HSC-T6凋亡比例虽较其他两组稍低,但差异无统计学意义(P>0.05)(见图2C)。
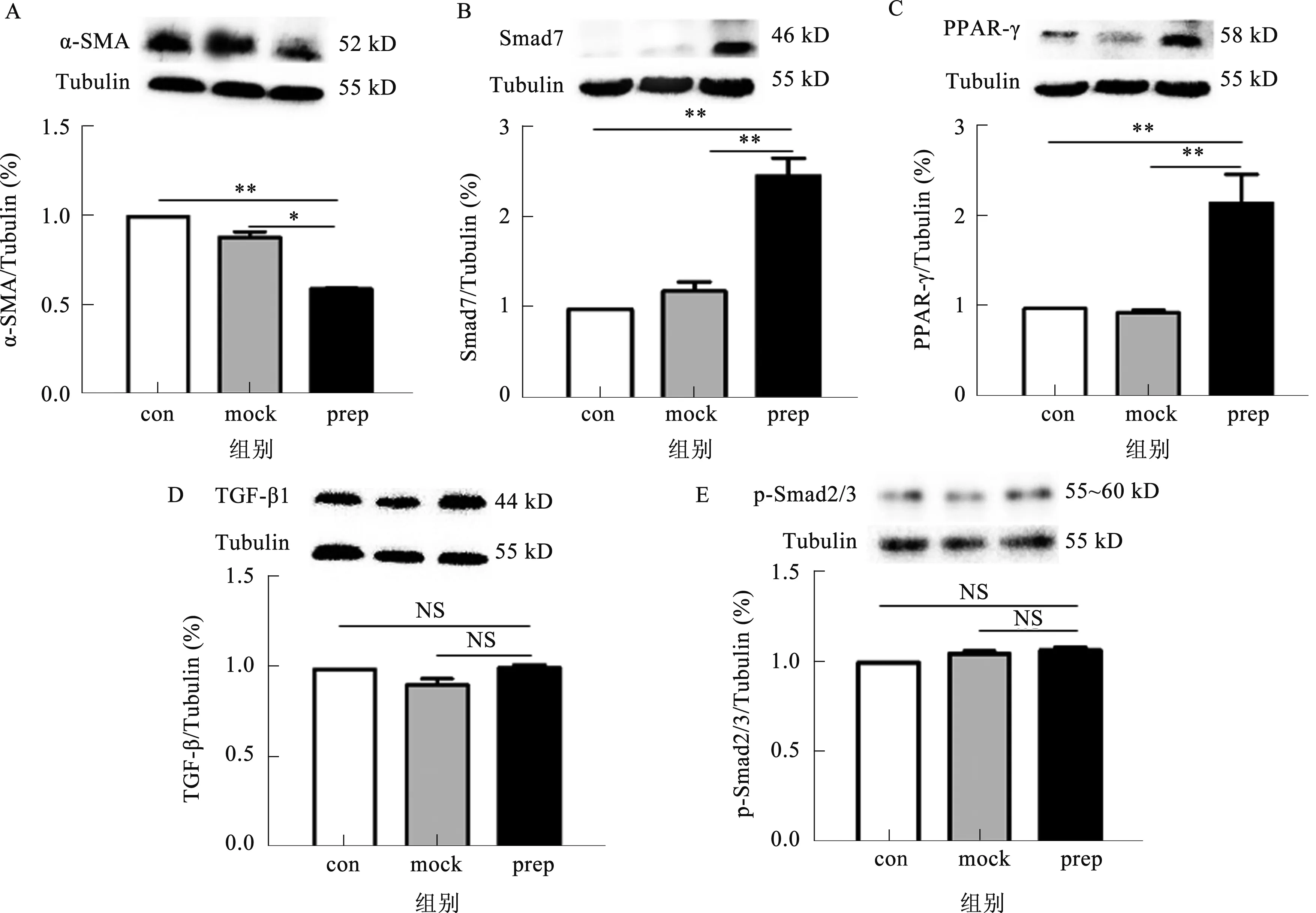
2.4POP慢病毒感染对细胞炎症与纤维化基因mRNA和蛋白表达的影响POP慢病毒组MCP-1及α-SMA mRNA表达均较正常对照组显著下降,TGF-β1和Col I mRNA表达各组间比较,差异无统计学意义(P>0.05)(见图3)。POP病毒组α-SMA蛋白表达较正常对照组显著下调,而Smad7、PPAR-γ蛋白表达较正常对照组显著上调,TGF-β1和p-Smad2/3蛋白表达在各组间差异无统计学意义(P>0.05)(见图4)。

注:con:正常对照组;mock;空病毒组;prep;POP慢病毒组。*P<0.05,**P<0.01,NS:无差异。

注:con:正常对照组;mock:空病毒组;prep:POP慢病毒组。*P<0.05,**P<0.01。
3 讨论
本研究利用携带POP基因的慢病毒感染HSC,发现POP可抑制HSC分泌炎性因子MCP-1及α-SMA的表达,并可能通过上调Smad7、PPAR-γ的表达抑制HSC的活化,从而具有抗纤维化的作用。
从HSC-T6表达荧光蛋白、胞内POP基因转录水平及蛋白表达均提示POP慢病毒感染成功,且在细胞内稳定表达POP;同时测定各组HSC-T6胞内Ac-SDKP水平变化,提示细胞内表达的POP存在活性;Ac-SDKP为POP水解Tβ4的产物[14],具有抗多种器官纤维化作用[8-9,15-16]。本课题组[9]在前期工作中已探索Ac-SDKP抗CCl4诱导肝纤维化的机制,主要为抑制TGF-β1和p-Smad2/3的表达,同时抑制HSC的增殖生长,对Smad7表达无影响。本研究发现,POP对TGF-β1和p-Samd2/3表达无影响,而是促进Smad7表达,而Smad7为TGF-Smad信号通路抑制蛋白,具有抗纤维化作用。同时研究发现,POP明显促进HSC-T6的增殖,与Ac-SDKP作用相反;通过本研究可以推断水解酶活性仅是POP一小部分功能,POP更重要的功能在于与其他蛋白结合的协同作用,调控基因转录和细胞生长,这种非酶解功能在中枢系统中研究较多[17-19],在肝脏系统中研究甚少,值得我们进一步深入研究。此外,POP抑制α-SMA和MCP-1的表达,α-SMA为活化HSC的标志,MCP-1为HSC分泌的主要促炎促纤维化因子[20],与前期Ac-SDKP作用结果一致。值得一提的是,POP与其下游产物Ac-SDKP在抗肝纤维化效应的具体作用仍不能完全区分,目前缺乏有效方法可以将Ac-SDKP抑制。
我们还发现,POP的HSC-T6胞内PPAR-γ表达显著升高,后者最早被认为是重要成脂基因之一[21],后越来越多证据证实其为HSC活化和表型转化的关键因子,在维持HSC保持静止状态发挥重要作用,可以抑制α-SMA和TGF-β1等表达,是抑制肝脏炎症纤维化重要因子[22-23]。另有研究[24-26]发现,PPAR-γ可以阻断TGF-β信号通路及抑制Smad蛋白依赖的启动子发挥作用,直接拮抗Smad3在成纤维细胞中的作用,但是不减少Smad3的蛋白表达也不增加Smad7的表达。HSC另外一个特点是储存维生素A类物质,随着其活化,这种物质会逐渐减少直至消失,尽管这种现象与HSC活化、疾病进展关系未完全阐明[27-29]。PPAR-γ可以使HSC恢复这种聚类维生素A物质功能,同时减轻胰岛素抵抗、脂肪性肝炎[23],同时我们课题组在近期的研究工作中发现,POP通过调控肝细胞内脂质代谢合成基因抑制肝细胞内脂质代谢[30];进一步结合前面所述POP与HSC增殖的关系,表面上POP促进HSC的增殖可促进纤维化进展,但POP促进HSC增殖的同时也使其从活化状态转变为静止储脂状态,进一步抑制其促纤维化活性[31]。因此,我们推断POP在脂肪性肝炎及其所致纤维化方面也存在重要作用,同时调控HSC活性。
注:con:正常对照组;mock组别:空病毒组;prep:POP慢病毒组。*P<0.05,**P<0.01,NS:无差异。
总之,本研究证实了POP蛋白酶在HSC中生物学功能显著,反映其可能具备肝内抗炎、抗纤维化作用,仍需进一步体内实验验证。但是肝脏系统复杂,本研究仅针对HSC,POP肝内具体功能仍存在许多未解之谜,尤其它的非酶解功能,值得进一步深入挖掘探索,可能为肝脏疾病提供新的诊疗切入点。
[1] BABKOVA K, KORABECNY J, SOUKUP O, et al. Prolyl oligopeptidase and its role in the organism: attention to the most promising and clinically relevant inhibitors [J]. Future Med Chem, 2017, 9(10): 1015-1038. DOI: 10.4155/fmc-2017-0030.
[2] MARUYAMA Y, MATSUBARA S, KIMURA A P. Mouse prolyl oligopeptidase plays a role in trophoblast stem cell differentiation into trophoblast giant cell and spongiotrophoblast [J]. Placenta, 2017, 53: 8-15. DOI: 10.1016/j.placenta.2017.03.004.
[3] TANAKA S, SUZUKI K, SAKAGUCHI M. The prolyl oligopeptidase inhibitor SUAM-14746 attenuates the proliferation of human breast cancer cell lines in vitro [J]. Breast Cancer, 2017, 24(5): 658-666. DOI: 10.1007/s12282-017-0752-5.
[4] PENTTINEN A, TENORIO-LARANGA J, SIIKANEN A, et al. Prolyl oligopeptidase: a rising star on the stage of neuroinflammation research [J]. CNS Neurol Disord Drug Targets, 2011, 10(3): 340-348.
[5] SUZUKI K, SAKAGUCHI M, TANAKA S, et al. Prolyl oligopeptidase inhibition-induced growth arrest of human gastric cancer cells [J]. Biochem Biophys Res Commun, 2014, 443(1): 91-96. DOI: 10.1016/j.bbrc.2013.11.051.
[6] TENORIO-LARANGA J, MNNISTÖP T, STORVIK M, et al. Four day inhibition of prolyl oligopeptidase causes significant changes in the peptidome of rat brain, liver and kidney [J]. Biochimie, 2012, 94(9): 1849-1859. DOI: 10.1016/j.biochi.2012.04.005.
[7] YAMAKAWA N, SHIMENO H, SOEDA S, et al. Regulation of prolyl oligopeptidase activity in regenerating rat liver [J]. Biochim Biophys Acta, 1994, 1199(3): 279-284.
[8] ZHANG L, XU L M, CHEN Y W, et al. Antifibrotic effect of N-acetyl-seryl-aspartyl-lysyl-proline on bile duct ligation induced liver fibrosis in rats [J]. World J Gastroenterol, 2012, 18(37): 5283-5288. DOI: 10.3748/wjg.v18.i37.5283.
[9] CHEN Y W, LIU B W, ZHANG Y J, et al. Preservation of basal AcSDKP attenuates carbon tetrachloride-induced fibrosis in the rat liver [J]. J Hepatol, 2010, 53(3): 528-536. DOI: 10.1016/j.jhep.2010.03.027.
[10] REYES-GORDILLO K, SHAH R, ARELLANES-ROBLEDO J, et al. Protective effects of thymosin β4 on carbon tetrachloride-induced acute hepatotoxicity in rats [J]. Ann N Y Acad Sci, 2012, 1269: 61-68. DOI: 10.1111/j.1749-6632.2012.06728.x.
[11] XIAO Y, QU C, GE W, et al. Depletion of thymosin β4 promotes the proliferation, migration, and activation of human hepatic stellate cells [J]. Cell Physiol Biochem, 2014, 34(2): 356-367. DOI: 10.1159/000363005.
[12] ATZORI L, POLI G, PERRA A. Hepatic stellate cell: a star cell in the liver [J]. Int J Biochem Cell Biol, 2009, 41(8-9): 1639-1642. DOI: 10.1016/j.biocel.2009.03.001.
[13] FRIEDMAN S L. Evolving challenges in hepatic fibrosis [J]. Nat Rev Gastroenterol Hepatol, 2010, 7(8): 425-436. DOI: 10.1038/nrgastro.2010.97.
[14] CAVASIN M A, RHALEB N E, YANG X P, et al. Prolyl oligopeptidase is involved in release of the antifibrotic peptide Ac-SDKP [J]. Hypertension, 2004, 43(5): 1140-1145. DOI: 10.1161/01.HYP.0000126172.01673.84.
[15] KANASAKI K, NAGAI T, NITTA K, et al. N-acetyl-seryl-aspartyl-lysyl-proline: a valuable endogenous anti-fibrotic peptide for combating kidney fibrosis in diabetes [J]. Front Pharmacol, 2014, 5: 70. DOI: 10.3389/fphar.2014.00070.
[16] CHAN G C, WU H J, CHAN K W, et al. N-acetyl-seryl-aspartyl-lysyl-proline mediates the anti-fibrotic properties of captopril in unilateral ureteric obstructed BALB/C mice [J]. Nephrology (Carlton), 2017. DOI: 10.1111/nep.13000.
[18] DI DANIEL E, GLOVER C P, GROT E, et al. Prolyl oligopeptidase binds to GAP-43 and functions without its peptidase activity [J]. Mol Cell Neurosci, 2009, 41(3): 373-382. DOI: 10.1016/j.mcn.2009.03.003.
[19] SAVOLAINEN M H, YAN X, MYÖHNEN T T, et al. Prolyl oligopeptidase enhances α-synuclein dimerization via direct protein-protein interaction [J]. J Biol Chem, 2015, 290(8): 5117-5126. DOI: 10.1074/jbc.M114.592931.
[20] MARRA F, TACKE F. Roles for chemokines in liver disease [J]. Gastroenterology, 2014, 147(3): 577-594, e1. DOI: 10.1053/j.gastro.2014.06.043.
[21] SAMUEL V T, SHULMAN G I. Nonalcoholic fatty liver disease as a nexus of metabolic and hepatic diseases [J]. Cell Metab, 2018, 27(1): 22-41. DOI: 10.1016/j.cmet.2017.08.002.
[22] ZARDI E M, NAVARINI L, SAMBATARO G, et al. Hepatic PPARs: their role in liver physiology, fibrosis and treatment [J]. Curr Med Chem, 2013, 20(27): 3370-3396.
[23] YU J, ZHANG S, CHU E S, et al. Peroxisome proliferator-activated receptors gamma reverses hepatic nutritional fibrosis in mice and suppresses activation of hepatic stellate cells in vitro [J]. Int J Biochem Cell Biol, 2010, 42(6): 948-957. DOI: 10.1016/j.biocel.2010.02.006.
[24] JEON K I, KULKARNI A, WOELLER C F, et al. Inhibitory effects of PPARγ ligands on TGF-β1-induced corneal myofibroblast transformation [J]. Am J Pathol, 2014, 184(5): 1429-1445. DOI: 10.1016/j.ajpath.2014.01.026.
[25] BIAN D, ZHANG J, WU X, et al. Asiatic acid isolated from centella asiatica inhibits TGF-β1-induced collagen expression in human keloid fibroblasts via PPAR-γ activation [J]. Int J Biol Sci, 2013, 9(10): 1032-1042. DOI: 10.7150/ijbs.7273.
[26] GHOSH A K, BHATTACHARYYA S, LAKOS G, et al. Disruption of transforming growth factor beta signaling and profibrotic responses in normal skin fibroblasts by peroxisome proliferator-activated receptor gamma [J]. Arthritis Rheum, 2004, 50(4): 1305-1318. DOI: 10.1002/art.20104.
[27] KLUWE J, WONGSIRIROJ N, TROEGER J S, et al. Absence of hepatic stellate cell retinoid lipid droplets does not enhance hepatic fibrosis but decreases hepatic carcinogenesis [J]. Gut, 2011, 60(9): 1260-1268. DOI: 10.1136/gut.2010.209551.
[28] PARK S, CHOI S, LEE M G, et al. Retinol binding protein-albumin domain Ⅲ fusion protein deactivates hepatic stellate cells [J]. Mol Cells, 2012, 34(6): 517-522. DOI: 10.1007/s10059-012-0183-2.
[29] BLANER W S, O’BYRNE S M, WONGSIRIROJ N, et al. Hepatic stellate cell lipid droplets: a specialized lipid droplet for retinoid storage [J]. Biochim Biophys Acta, 2009, 1791(6): 467-473. DOI: 10.1016/j.bbalip.2008.11.001.
[30] ZHOU D, LI B H, WANG J, et al. Prolyl oligopeptidase inhibition attenuates steatosis in the L02 human liver cell line [J]. PLoS One, 2016, 11(10): e0165224. DOI: 10.1371/journal.pone.0165224.
[31] MALLAT A, LOTERSZTAJN S. Reversion of hepatic stellate cell to a quiescent phenotype: From myth to reality? [J]. J Hepatol, 2013, 59(2): 383-386. DOI: 10.1016/j.jhep.2013.03.031.
猜你喜欢
中国病理生理杂志(2022年11期)2022-12-03
中老年保健(2022年1期)2022-08-17
传染病信息(2022年3期)2022-07-15
医药与保健(2021年4期)2021-04-07
兽医导刊(2019年1期)2019-02-21
猪业科学(2018年8期)2018-09-28
中华神经外科疾病研究杂志(2018年4期)2018-08-20
中华肺部疾病杂志(电子版)(2018年6期)2018-03-11
医学研究杂志(2015年6期)2015-07-01
中国中医药现代远程教育(2014年13期)2014-03-01
