
CO2加富与不同钾素水平供应对黄瓜气孔状态的影响
2018-10-11 06:23达布希拉图
江苏农业科学 2018年17期
葛 涛, 达布希拉图
(云南农业大学资源与环境学院,云南昆明 650201
植物叶片气孔是与外界气体进行交换的主要通道,其数量和开度等对CO2浓度的变化很敏感[1],当环境、温度等条件发生变化时,气孔孔径大小就通过保卫细胞来改变,进而调节植物的蒸腾、呼吸、光合等重要生理活动,进而适应变化的环境[2]。
长期CO2加富导致光合能力减弱的现象称为光合适应[3]。长期高浓度CO2下生长的植物,其叶片气孔会发生光合适应现象,即其气孔导度显著低于常规大气条件下生长的植物[4-7]。Woodward研究植物气孔密度与大气CO2浓度的相关关系时首次证实,气孔密度的下降限制了CO2通过气孔进入,从而利于光合作用对高CO2浓度的适应[8]。另外,Gunderson等研究认为,植物的光合适应可能与气孔开度在CO2浓度升高时下降较低有关[9]。也有研究表明,气孔导度随CO2浓度的增大而减小[4]。这些研究多从不同角度分析高浓度CO2下气孔与光合适应的关系,且较集中于某个气孔状态指标,而从气孔密度、大小、开度及气孔导度等全面研究气孔状态与光合适应关系的相关报道较少,且在CO2加富和不同钾素水平下针对气孔状态的研究更是鲜见。本试验通过研究高浓度CO2条件下气孔与光合适应的内在机制及钾素应用能否缓解光合适应,以期为设施大棚CO2气肥的推广奠定基础。
1 材料与方法
1.1 试验处理
本试验于2015年8—10月在云南农业大学后山资源与环境学院温室大棚内进行,大棚内建设1个长2.5 m、宽 1.5 m、高1.5 m,由钢管支撑的长方体开顶式薄膜培养室。室内CO2浓度由美国GE产Telaire7001 型红外二氧化碳检测仪测定。采用CO2纯度为99.9%的高压钢瓶气体经减压缓冲向室内供气。
供试黄瓜品种为中农20号,将其种子在26 ℃培养箱中恒温催芽48 h,其间加水1次/d,保持滤纸湿润;将经催芽的种子播于装有蛭石的育种盆中育苗,25 ℃恒温培育至3片真叶时,移栽至清水中1周;使用修正的1/4强度Hogland营养液继续培养至6片真叶,钾素用K2SO4和KCl调节,钾质量浓度为60 mg/L;选生长整齐一致的幼苗,定植于装3 L 1/2强度Hogland营养液的塑料盆中,分别置于2个CO2控制室通入CO2,一为正常室,通入正常空气(自然条件),CO2摩尔分数为380 μmol/mol,一为升高室(CO2加富),CO2摩尔分数为1 000 μmol/mol,同时每个控制室设置2个钾素水平,用硫酸钾调节钾质量浓度分别为 20、80 mg/L。共计4个处理,重复6次,每盆1株,共计24盆。用通气泵24 h通气,CO2加富时间为每天08:00—17:00。营养液pH值为6.0,每3 d更换1次。分别于通入CO2气体后4、14、30 d分3次采样测定。
1.2 测定内容及方法
1.2.1 光合速率与气孔导度 采用美国LI-COR公司生产的便携式LI-6400光合作用分析仪测定黄瓜叶片光合速率与气孔导度,测定条件为发光强度800 cd、温度25 ℃。
1.2.2 气孔开度、气孔密度、气孔大小 采用透明胶带法[10]测定:将塑料透明胶带拉开10 cm左右,左手捏住叶柄将叶片正面由叶尖开始逐渐向前推移,使叶片正面平整地粘贴在胶带上,先粘1~2次以清理叶片表面灰尘;将胶带对折,手指用力对捏胶带,使胶带与叶片的两面充分粘贴;将对折的胶带撕开,叶片的下表皮就粘贴在胶带上,并置于提前准备的盒中;电子显微镜40倍下观察、测量、计数叶片下表皮气孔开度、密度和大小。塑料透明胶带宽度为18 mm,厚度为0.05 mm。
1.2.3 植株生物量 分别于通入CO2气体后3、30 d采集植株,用纯水冲洗干净;吸水纸吸干植株表面的水分,把叶、茎、果、根分开,分别称量鲜质量;105 ℃杀青30 min,75 ℃烘干至恒质量,测定干质量。
1.3 数据处理
采用Excel 2007、SPSS 17. 0软件进行数据的处理和统计分析。
2 结果与分析
2.1 CO2加富与不同K素水平对黄瓜生物量的影响
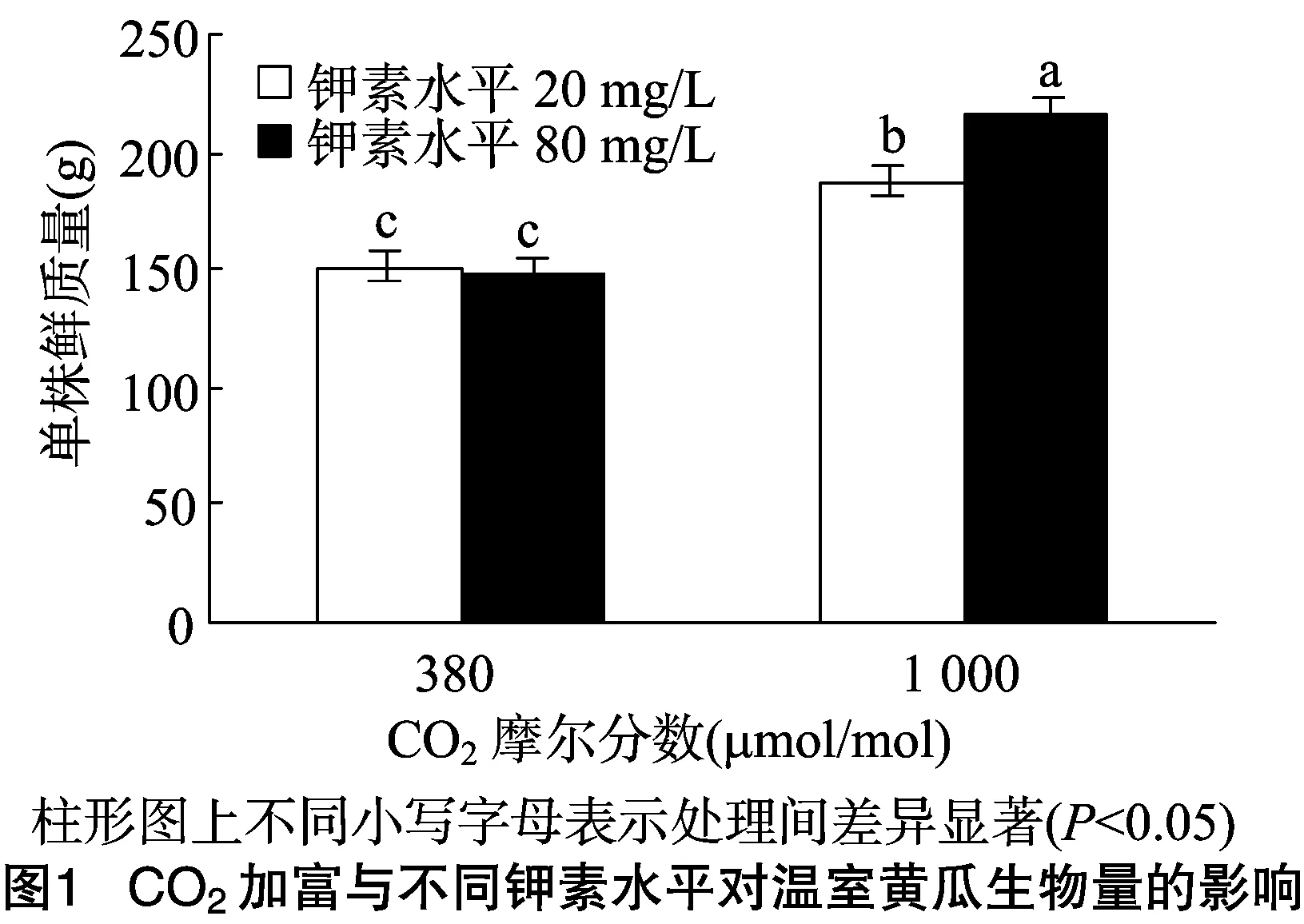
由图1可见,在不同K素水平上,经CO2加富(CO2摩尔分数为1 000 μmol/mol)的黄瓜,其植株生物量显著大于自然条件下(CO2摩尔分数为380 μmol/mol)生长的黄瓜(P<0.05);自然条件下,80 mg/L钾素处理的黄瓜单株鲜质量与 20 mg/L 钾素处理的差异不显著(P>0.05),CO2加富条件下,80 mg/L钾素处理的黄瓜单株鲜质量显著大于20 mg/L钾素处理的(P<0.05)。
2.2 CO2加富条件下黄瓜叶片净光合速率变化
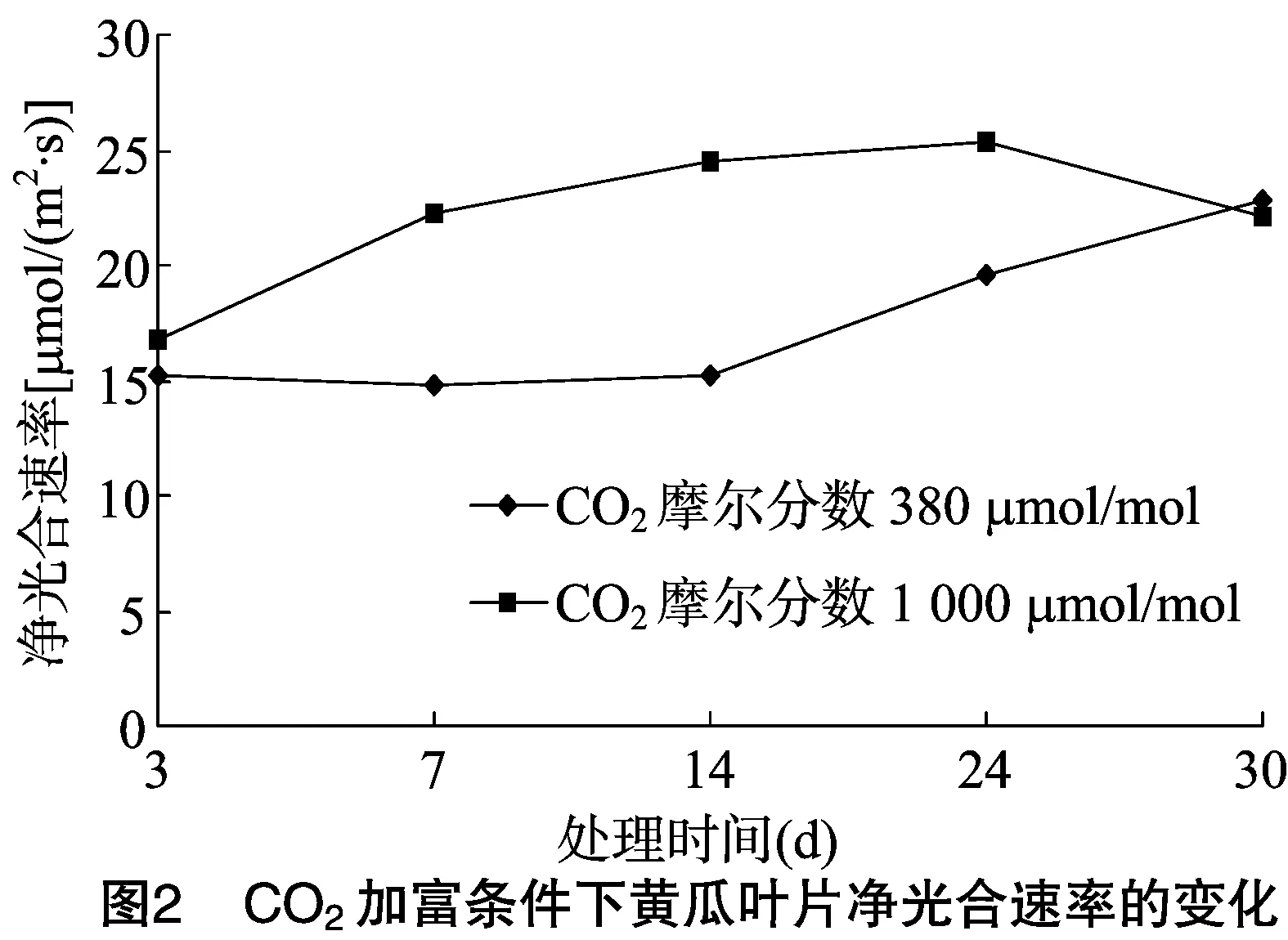
由图2可见,与自然条件相比,CO2加富可使黄瓜叶片的净光合速率明显升高,加富处理24 d时达到峰值;CO2加富30 d时的黄瓜叶片净光合速率低于自然CO2条件下的,说明长期阶段性CO2加富可降低黄瓜叶片的净光合速率,即出现光合适应现象。
2.3 CO2加富时间及不同钾素水平对黄瓜叶片气孔导度、密度、开度、大小的影响
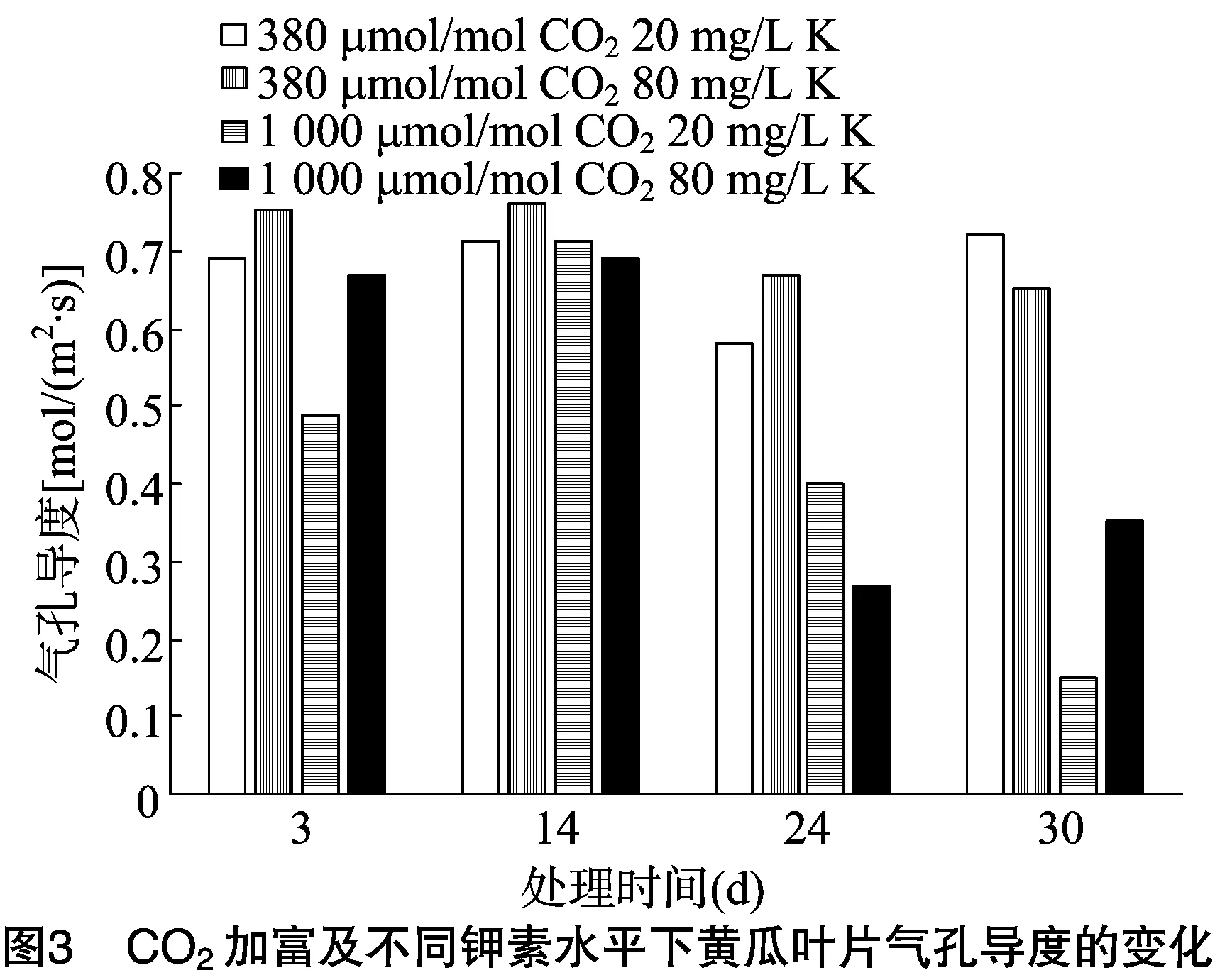
2.3.1 对黄瓜叶片气孔导度的影响 由图3可见,在黄瓜整个生长期,CO2加富的黄瓜叶片气孔导度低于自然条件下的;自然条件下,20 mg/L 钾素处理后3、14、24 d的黄瓜叶片气孔导度均低于80 mg/L 钾素处理的,而在处理后30 d高于 80 mg/L 钾素处理的,这与黄瓜叶片光合适应的产生时间相吻合;CO2加富条件下, 80 mg/L钾素处理后3 d的黄瓜叶片气孔导度高于20 mg/L 钾素处理的,而在处理后14、24 d低于20 mg/L 钾素处理的,处理后30 d时又高于20 mg/L 钾素处理的,这也与黄瓜叶片光合适应的产生时间相吻合。由表1可见,通气后3、30 d,自然条件或CO2加富条件下不同钾素水平的黄瓜叶片,其气孔导度相互间差异不显著(P>0.05);通气后3 d时,20 mg/L 钾素水平条件下,自然条件和CO2加富处理的黄瓜叶片气孔导度相互间有显著性变化,而 80 mg/L 钾素水平条件下相互间差异不显著;通气后30 d时,20、80 mg/L钾素水平条件下,自然条件和CO2加富处理的黄瓜叶片气孔导度相互间差异均为显著(P<0.05)。
2.3.2 对黄瓜叶片气孔密度、开度、大小的影响 由表1可见,通气后3 d,自然条件下不同钾素水平处理的黄瓜叶片气孔密度差异显著(P<0.05),而气孔开度差异不显著;CO2加富条件下,不同钾素水平处理的黄瓜叶片气孔开度差异显著(P<0.05),而气孔密度差异不显著;20 mg/L钾素水平条件下,自然条件和CO2加富处理的黄瓜叶片,其气孔开度有显著性变化(P<0.05),而气孔密度差异不显著;80 mg/L钾素水平条件下,自然条件和CO2加富处理的黄瓜叶片,其气孔密度、气孔开度相互间有显著性变化(P<0.05)。通气后 30 d,CO2加富条件下不同钾素水平处理的黄瓜叶片,其气孔开度、气孔密度相互间有显著性变化(P<0.05);自然条件下,不同钾素水平处理的黄瓜叶片,其气孔密度、气孔开度相互间差异不显著;20 mg/L 钾素水平条件下,自然条件和CO2加富处理的黄瓜叶片,其气孔密度、气孔开度相互间有显著性变化(P<0.05);80 mg/L钾素水平条件下,自然条件和CO2加富处理的黄瓜叶片气孔开度相互间差异显著(P<0.05),而气孔密度差异不显著。通气后3、30 d,各处理黄瓜叶片的气孔长度相互间差异不显著。
2.3.3 黄瓜叶片气孔导度、气孔密度、气孔开度、气孔大小的相关性分析 果期时,自然条件下黄瓜叶片气孔开度与气孔密度呈极显著负相关(P<0.01),苗期时呈负相关,而CO2加富条件下黄瓜叶片气孔开度与气孔密度在苗期时呈显著正相关(P<0.05),在果期时呈负相关,这说明CO2加富处理可能使黄瓜叶片产生了光合适应;在自然条件下,黄瓜叶片气孔导度与气孔密度在苗期时呈负相关,在果期时正相关,而在CO2加富条件下正好相反,黄瓜叶片气孔导度与气孔密度在苗期时呈正相关, 在果期时负相关;苗期与果期时,气孔导度与气孔开度呈正相关,但相关性不显著(表2)。

表1 CO2加富时间及不同钾素水平条件下黄瓜气孔导度、密度、开度、大小的变化
注:同列数据后不同字母表示同一调查时期处理间差异显著(P<0.05)。小写字母表示同一CO2浓度下不同钾素浓度条件下的差异显著性;大写字母表示同一钾素浓度不同CO2浓度条件下的差异显著性。
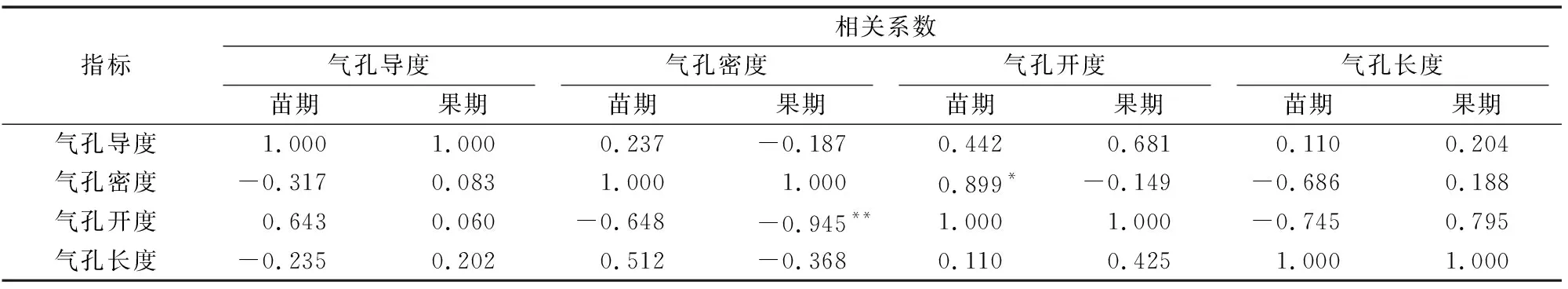
表2 苗期与果期黄瓜叶片气孔状态的相关性
注:数据后标注*、**分别表示两者之间0有显著(P<0.05)、极显著性(P<0.01)相关。表中左下角为自然条件下的相关系数,右上角为CO2加富条件下的相关系数。苗期为CO2加富后3 d,果期为CO2加富后30 d。
3 结论与讨论
环境CO2浓度会增高到720 μmol/mol,甚至更高[11],这对植物存在选择压力,必然要通过自身形态及生理、生化调节来适应这种不断变化的环境。在环境中CO2浓度升高时,植物会通过关闭气孔等来减少对CO2的吸收,但在长期高浓度CO2条件下,植物减少对CO2吸收最有效的方式往往是通过减少气孔数量来实现的[12-13]。有研究表明,叶片气孔的大小和密度变化对气孔导度有一定影响,提高气孔导度可以促进作物的光合作用,进而影响光合产物的生成。本试验结果表明,高CO2浓度(CO2摩尔分数为1 000 μmol/mol)与常规CO2浓度(CO2摩尔分数为380 μmol/mol)相比,黄瓜叶片气孔导度会降低,这与蒋跃林等的研究结果[14]一致;长期高浓度CO2处理,黄瓜叶片气孔导度会下降到更低水平,出现光合适应现象,而不同CO2浓度条件下,不同钾素水平处理的黄瓜叶片气孔导度变化趋势相反,说明较高的钾素水平有可能缓解光合适应的发生。
有研究表明,大气CO2浓度与植物气孔密度存在负相关关系。郑凤英等统计狭叶山黄麻和光叶山黄麻近1个世纪的蜡叶标本气孔密度时发现,从20世纪20年代至90年代,狭叶山黄麻和光叶山黄麻的气孔密度分别降低28.1%、40.0%,且2种植物的气孔密度与大气CO2浓度呈显著负相关[15];Ramalho等研究表明,随着CO2浓度的升高,叶片气孔密度有明显下降趋势,当CO2浓度上升到 550 μmol/mol 时下降非常显著,而CO2浓度在 550~700 μmol/mol 之间时,气孔密度下降趋于平缓[11]。也有一些研究表明,大气CO2浓度与植物气孔性状的相关关系存在很多可能性[16],如当环境中CO2浓度超过340 mg/L时,植物的气孔性状没有发生变化,或为植物的气孔性状与CO2浓度呈正相关[17-18],这可能与采样时间、叶片生育期及植物基因型不同有关[19-21]。
猜你喜欢
生态学报(2024年1期)2024-01-25
干旱地区农业研究(2022年1期)2022-01-28
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
装备制造技术(2019年12期)2019-12-25
中国铸造装备与技术(2015年5期)2015-12-10
植物营养与肥料学报(2014年1期)2014-03-11
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2011年2期)2011-10-26
