
培养温度对腐败希瓦氏菌DSM6067生长动力学及细胞膜理化特性的影响
2020-03-06 15:11杨胜平钱韻芳
中国食品学报 2020年2期
杨胜平 章 缜 程 颖 钱韻芳 谢 晶*
(1 上海海洋大学食品学院 上海201306
2 上海水产品加工及贮藏工程技术研究中心 上海201306)
冷链流通是保持生鲜水产品品质的重要手段,研究发现低温冷藏并不能完全抑制微生物的生长,部分能够适应低温环境的腐败微生物仍会生长繁殖,并最终导致冷藏水产品腐败变质[1-3],这已成为水产品冷链流通保鲜的主要限制因素。低温冷藏条件对嗜热菌、嗜中温菌的生长有明显抑制作用,而对水产品中主要存在的耐冷菌(psychrotrophs)的抑制作用有限。国内外研究发现,耐冷菌在鲻鱼[4]、凡纳滨对虾[5-6]、鱿鱼圈[7]、史氏鲟鱼糜[8]等冷藏水产品微生物菌群中占主导地位。
在这些水产品耐冷菌中,以腐败希瓦氏菌(Shewanella putrefaciens)最为典型。腐败希瓦氏菌是冷藏凡纳滨对虾、带鱼的主要腐败菌[2,9-10],同时它也是牙鲆、大黄鱼等大宗水产品的主要腐败菌[11]。腐败希瓦氏菌是一种革兰氏阴性运动杆菌,具有较强的致腐败能力,在其生长代谢活动中不仅能够分解肌肉蛋白,将氨基酸脱羧生成腐胺、尸胺等生物胺[12],还可还原氧化三甲胺(TMAO)为三甲胺(TMA)[13],产生H2S等臭味物质[14],其形成的生物被膜可导致水产品表面发粘[15]。腐败希瓦氏菌是导致低温贮藏水产品发生腐败的重要因素之一。
腐败希瓦氏菌具有较强的温度适应能力,不仅能在常温下较快生长,也能适应低温环境并保持一定生长速率,因而成为水产品中的优势菌。有研究表明,腐败希瓦氏菌在新鲜水产品微生物菌群中的比例较小,在低温贮藏过程中才迅速增长,如新鲜凡纳滨对虾中腐败希瓦氏菌的数量仅占微生物菌落总数的1.6%,而4℃贮藏10 d后就增加到约24.7%[2]。在低温贮藏的草鱼片[12]、大西洋马鲛鱼[16]、牡蛎[17]等水产品中亦有类似发现。如何抑制低温下腐败希瓦氏菌的生长成为水产品冷藏保鲜中亟待解决的问题。
低温胁迫下腐败希瓦氏菌适冷调控机制对其生长动力学起着至关重要的作用,在适冷过程中,微生物细胞膜流动性的维持是其在低温环境下进行正常代谢的关键因素[18],而流动性的维持主要通过调节细胞膜中脂肪酸组成,降低膜脂熔点来实现。一般认为常见的调节方式包括:①增加不饱和脂肪酸(unsaturated fatty acid,UFA)含量;②缩短脂肪酸酰基链的长度;③增加支链脂肪酸和异构分支脂肪酸的比例;④减少环状脂肪酸的含量[19]。其中,以脂肪酸不饱和度的调节最为关键,该调控通路可作为抑菌的靶标[20]。
目前,有关腐败希瓦氏菌对细胞膜脂肪酸含量调节机制的报道较少。本文以水产品中常见的腐败希瓦氏菌的模式菌株DSM6067为对象,研究不同培养温度条件下的腐败希瓦氏菌生长动力学参数、细胞膜脂肪酸组成、膜蛋白含量、细胞微观结构和细胞膜流动性变化,分析温度对腐败希瓦氏菌生长动力学参数及其细胞膜理化特性的影响,以期为通过表征细胞蛋白质组差异表达,推测温度敏感因子和不饱和脂肪酸代谢通路;通过研究不饱和脂肪酸代谢通路中去饱和酶基因的转录量以及转录调控因子含量与温度的关系,探讨腐败希瓦氏菌在适冷过程中的分子调控机理。对于进一步研究抑制冷链流通中腐败希瓦氏菌的生长,保障水产品品质的方法提供一定的理论依据。
1 材料与方法
1.1 材料、试剂与仪器
1.1.1 腐败希瓦氏菌模式菌株 腐败希瓦氏菌模式菌株DSM 6067,德国Leibniz-Institut DSMZ GmbH微生物研究所。
1.1.2 试剂与仪器 试剂:脑心浸肉汤(BHI)、胰蛋白胨大豆肉汤(TSB)等微生物培养基,青岛海博生物技术有限公司;8-苯胺-1-萘磺酸(ANS),sigma公司;Bradford蛋白浓度测定试剂盒,上海碧云天生物技术有限公司;12%电泳预制胶、电泳缓冲液,南京建成生物有限公司;双色预染蛋白Marker、蛋白Loading Buffer、C510041高灵敏快速考马斯亮蓝染色试剂盒,生工生物工程(上海)股份有限公司。甲醇、正己烷、氯仿、十九烷酸甲酯、浓盐酸、磷酸氢二钠、磷酸二氢钠等试剂,国药集团化学试剂有限公司。
主要设备与仪器:LDZX-50KBS蒸汽灭菌器,上海申安公司;VS-1300L-U洁净工作台,苏州苏净泰安集团;Eppendorf 5427R型高速冷冻离心机,德国艾本德公司;H-2050R台式冷冻离心机,湖南湘仪仪器有限公司;Ymnl-150型超声波细胞粉碎机,南京以马内利仪器设备有限公司;SHA-2型制冷恒温振荡器,金坛市城西峥嵘实验仪器厂;JEM-1200EX透射电子显微镜,日本JEOL公司;RF-6000荧光分光光度计,日本岛津公司;Trace 1310 ISQ气相色谱质谱联用仪,ThermoFisher公司;iMark酶标仪、Mini-PROTEAN Tetra电泳仪、小型Trans-Blot垂直电泳槽,美国Bio-Red伯乐生命医学产品有限公司。
1.2 方法
1.2.1 菌种活化 腐败希瓦氏菌模式菌株DSM 6067菌粉溶解于已灭菌的BHI培养基中,于30℃下活化培养12~18 h至菌液浓度达到108CFU/mL。取适量菌液接种于已灭菌的TSB培养基中再次活化,待用。
1.2.2 微生物生长曲线绘制及腐败菌生长动力学模型的选取 细菌生长曲线的绘制:吸取0.1mL活化培养至108CFU/mL的TSB菌液于装有100 mL新灭菌的TSB培养液的三角瓶中,分别在30,10℃和4℃下培养,在波长595 nm处测定菌液的OD值,根据OD595值绘制生长曲线。
腐败希瓦氏菌生长动力学模型的选取:参照傅鹏等[21]及Zwietering等[22]修正的Gompertz方程描述的腐败菌生长动态方法。
1.2.3 细菌脂肪酸组成及含量的测定 参照Diomandé等[23]的方法并略有修改。将细菌培养至各温度下对应的对数生长中期(OD595值约为0.6),将菌液5 000 g离心,沉淀经PBS缓冲溶液反复冲洗3次,收集菌体。菌体经液氮猝灭,于-80℃隔夜冻藏,次日冷冻干燥成菌粉。取80~100 mg冻干菌粉加入15mL离心管中,继续加入2mL 5%盐酸甲醇溶液,3mL体积比为1∶1的氯仿-甲醇溶液,100μL十九烷酸甲酯内标。在85℃水浴锅中水浴1 h后降至室温,在离心管中加入1mL正己烷,震荡萃取2min,静置1 h,待分层。取上层清液100μL,用正己烷定容1mL,0.45μm滤膜过膜后上机分析。
色谱条件:色谱柱:TG-5MS(30m×0.25mm×0.25μm),升温程序:80℃保持1min,以10℃/min的速率升至200℃,继续以5℃/min的速率升至250℃,最后以2℃/min的速率升到270℃,保持3 min。进样口温度:290℃,载气流速:1.2mL/min,不分流进样,开阀时间1min。
质谱条件:离子源温度:280℃,传输线温度:280℃,溶剂延迟时间:5.00min,扫描范围:30~400 amu,离子源:EI源70 eV。
1.2.4 细菌细胞膜蛋白聚丙烯酰胺凝胶电泳与膜蛋白含量的测定 将30,10℃和4℃条件下培养至对数生长中期(OD595值约为0.6)的细菌菌液于5 000 g下离心,沉淀经PBS缓冲溶液反复冲洗3次后收集菌体。聚丙烯酰胺凝胶电泳(SDSPAGE)参照雷雨婷等[24]的方法进行。样品经丙酮洗涤后,加入适量loading buffer煮沸5min,冷却后加样于1mm厚的凝胶孔道中,上样量5μL。膜蛋白含量参照Dong等[25]的方法,使用Bradford蛋白浓度测定试剂盒测定。
1.2.5 透射电子显微镜(TEM)观察细菌微观结构变化 将不同温度下培养至对数生长中期的菌株离心,收集菌体并用0.2mol/L磷酸缓冲液稍清洗,加入2.5%的戊二醛固定液(勿冲散菌体),4℃下固定过夜,切片,透射电子显微镜观察并拍照[26]。
1.2.6 细菌细胞膜流动性的测定 根据康亦兼等[27]的方法进行,略有修改。将活化好的菌种分别接种至100mL新灭菌的TSB培养液中,30℃培养8 h,此后将其转移,置4℃环境下适冷培养。分别在30℃培养8 h,4℃培养24,72,144 h及216 h取菌样,4℃下5 000 g离心10min,收集菌体,经PBS缓冲液清洗3次,重悬于含0.5mmol/L DTT和0.1%TWEEN 20的PBS缓冲液中。将菌液于冰浴下超声细胞粉碎机粉碎6min,之后将其在4℃下10 000 g离心20min,弃沉淀,测定上清液中蛋白浓度并保存于-40℃下待用。用Tris-Hcl缓冲液配制5mmol/L ANS母液,临用前将其稀释至1.0×10-4mol/L。膜蛋白终质量浓度为5mg/L,ANS终质量浓度为10μmol/L,于室温孵育20 min,用荧光分光光度计在激发波长385 nm、发射波长480 nm条件下测定。
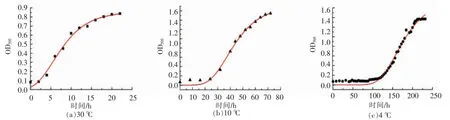
图1 不同培养温度下腐败希瓦氏菌的生长曲线Fig.1 The growth curve of Shewanella putrefaciens at different temperatures

表1 不同培养温度下腐败希瓦氏菌的生长动力学参数Table1 The growth parameters of Shewanella putrefaciens at different temperatures
2 结果与分析
2.1 腐败希瓦氏菌在30,10℃及4℃条件下的生长情况
腐败希瓦氏菌模式菌株在30,10℃和4℃下的生长动态采用修正的Gompertz方程描述,在细菌生长期间OD595值拟合曲线如图1所示。通过Compertz方程拟合所得腐败希瓦氏菌的生长动力学参数见表1。
Compertz方程拟合的30,10℃及4℃培养下的生长曲线相关系数R2分别为0.9949,0.9971和0.9859,具有较高的拟合度,能够较好地反映细菌的生长情况。由生长动力学参数可知腐败希瓦氏菌在30,10℃及4℃下生长的延滞时间分别为3.35,25.94 h和122.03 h,随着培养温度的下降,细菌的延滞时间延长。腐败希瓦氏菌在3种培养温度下的最大比生长速率μmax:0.047 h-1(10℃),其与30℃下的0.081 h-1相比下降较明显,而在4℃时的μmax仅为0.017 h-1。可见,培养温度越低,微生物的增长越缓慢。30℃是腐败希瓦氏菌的最适生长温度,细菌迅速增长繁殖,经短暂的延滞期后快速进入对数生长期,在8 h左右达到对数生长中期,并在24 h后基本趋于稳定。培养温度10℃下,希瓦氏菌经40 h培养后达到对数生长中期。培养温度4℃下的细菌增长速度较慢,腐败希瓦氏菌在前122 h均在延滞期,168 h后达到对数生长中期。由腐败希瓦氏菌生长动力学参数表明,不同培养温度下腐败希瓦氏菌DSM6067的增长虽然随培养温度的下降而减缓,但是仍能适应低温环境生长。
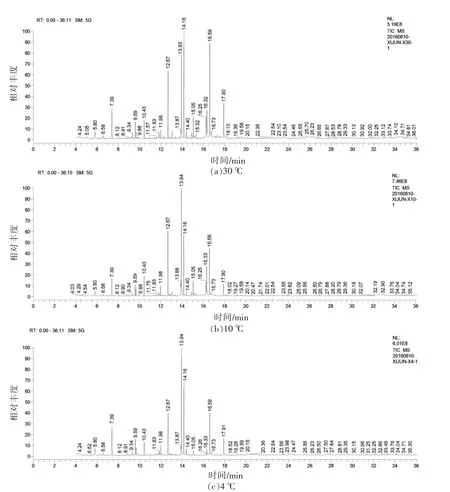
图2 不同培养温度下腐败希瓦氏菌细胞膜脂肪酸GC-MS图谱Fig.2 GC-MS of fatty acids in Shewanella putrefaciens cell membrane at different temperatures
2.2 30,10℃及4℃条件下腐败希瓦氏菌细胞膜脂肪酸组成含量变化
微生物为了适应低温环境,细胞膜脂肪酸组成会发生一定变化[28]。以C8:0(辛酸)、C24:1(神经酸)等共35种脂肪酸作为标准品,分析30,10℃和4℃不同培养温度下腐败希瓦氏模式菌株在对数生长中期的细胞膜中脂肪酸情况,最终检出13种脂肪酸。脂肪酸GC-MS图谱及检出的脂肪酸相对含量见图2和图3。从图3可知,腐败希瓦氏菌细胞膜脂肪酸以C16:0(棕榈酸)、C16:1(棕榈油酸)和C18:0(硬脂酸)为主。随着培养温度的下降,腐败希瓦氏菌细胞膜中的C15:0、C16:0、C17:0、C17:1、C18:0、C18:1N9C和C18:1N9T含量均下降,而C12:0、C14:0及C16:1含量则上升较为明显。其中4℃与30℃培养下细菌相比C16:1含量上升最为显著,该现象与窦京娇等[29]对北极海冰细菌在低温胁迫下脂肪酸的变化研究结果相一致。由此说明单不饱和脂肪酸C16:1在维持细胞膜流动性方面可能起关键性作用。
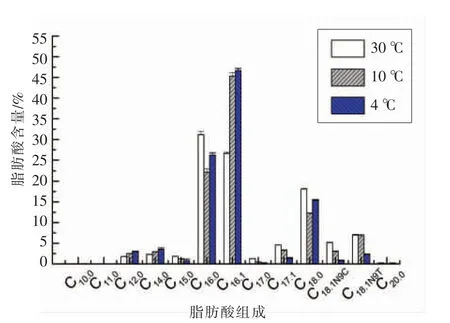
图3 不同培养温度下腐败希瓦氏菌细胞膜脂肪酸相对含量变化Fig.3 The changes of membrane fatty acids in Shewanella putrefaciens at different temperatures
2.3 30,10℃及4℃条件下腐败希瓦氏菌细胞膜蛋白含量的变化
腐败希瓦氏菌细胞膜蛋白SDS-PAGE电泳图谱如图4所示。随着培养温度的下降,腐败希瓦氏菌膜蛋白种类有所增加,其中低温培养下以分子质量在30 ku及60~120 ku间的蛋白表达量较多。Bisht S C等[30]有关嗜冷菌的研究表明,细菌在低温下产生的分子质量为30 ku的蛋白一般为冷激蛋白。图5显示腐败希瓦氏菌膜蛋白浓度情况。细菌膜蛋白浓度也随着培养温度的降低而上升。可见,腐败希瓦氏菌为了适应低温环境,其细胞膜蛋白表达种类和含量均有上升。对于不同培养温度下腐败希瓦氏菌所表达的差异蛋白种类还有待进一步研究。
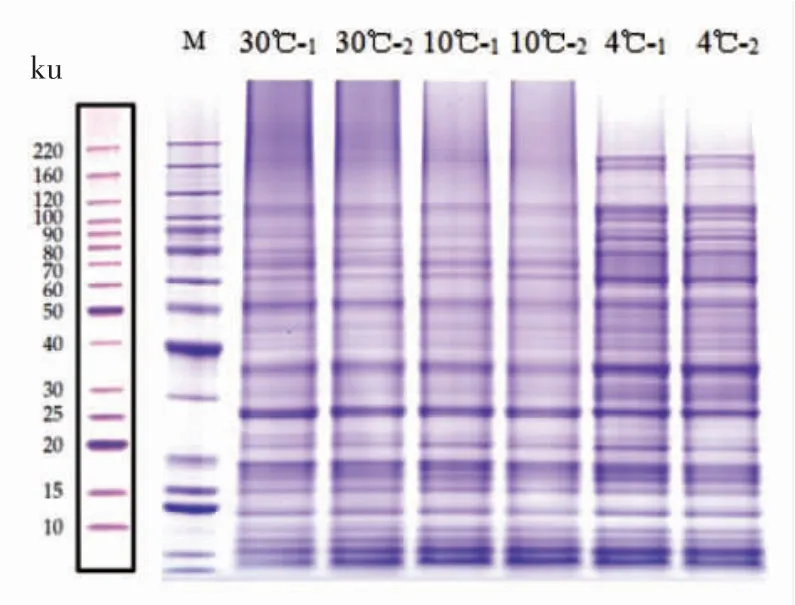
图4 不同培养温度下腐败希瓦氏菌细胞膜蛋白SDS-PAGE电泳图谱Fig.4 SDS-PAGE of membrane proteins in Shewanella putrefaciens at different temperatures
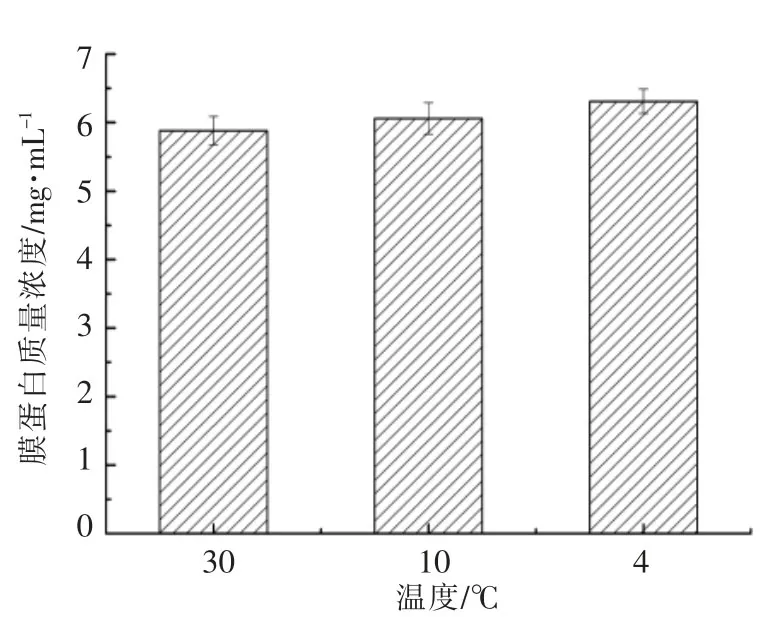
图5 不同培养温度下腐败希瓦氏菌细胞膜蛋白含量变化Fig.5 The changes of membrane proteins in Shewanella putrefaciens at different temperatures
2.4 30,10℃及4℃条件下腐败希瓦氏菌细胞微观形态变化
透射电子显微镜(TEM)观察腐败希瓦氏菌模式菌株DSM6067的细胞微观结构如图6所示。在30℃培养时,腐败希瓦氏菌的细胞壁膜呈现波浪状褶皱,可能是因为腐败希瓦氏菌的最适生长温度为30℃,细胞壁膜出现的褶皱使其表面积增大,使细胞有利于代谢营养物质的吸收。随着培养温度的降低,腐败希瓦氏菌细胞壁膜变得较为平滑,其表面积相对较小,从而影响其胞内外物质交换,细胞代谢增长速率下降。此外,透射电子显微镜下的腐败希瓦氏菌部分细胞内部出现空泡,可能与细胞内容物外漏有关,此现象与蔡瑾[31]用TEM观察细菌的结果类似。与10℃和4℃低温培养下的细菌相比,30℃培养下的腐败希瓦氏菌出现的空泡较多,可能是由于培养温度较高,细菌细胞膜流动性较强,在TEM样品前处理时易导致细胞膜破损,细胞内容物流出形成空泡。
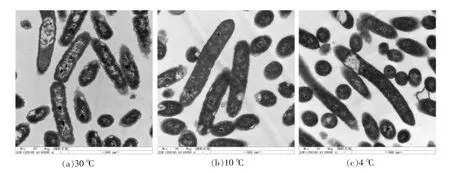
图6 不同培养温度下腐败希瓦氏菌细胞TEM显微结构变化Fig.6 The microscopic structure of Shewanella putrefaciens at different temperatures by TEM
2.5 腐败希瓦氏菌在冷适应过程中细胞膜流动性变化
8-苯胺-1-萘磺酸(8-Anilino-1-naphthalenesulfonic acid,ANS)是一种荧光染料,其常作为一种蛋白质荧光探测剂,较适用于标记测定亲和力疏水性配体及其相应的结合蛋白,如具有疏水性游离脂肪酸的脂肪酸结合蛋白(FABPs)。ANS的荧光强度值与细胞膜的流动性呈反比,即荧光强度越高,细胞膜的流动性越低[29]。本文采用ANS荧光探针法研究细胞膜流动性,不同培养温度下腐败希瓦氏菌细胞膜流动性如图7所示。腐败希瓦氏菌在波长485 nm处出现荧光强度峰值,细菌在30℃培养8 h,荧光强度值较低,说明细胞膜流动性较好。将其转移至4℃环境中培养24 h后,细胞膜流动性显著降低。可见,腐败希瓦氏菌在低温胁迫下细胞膜流动性下降。随着4℃低温环境中培养时间的延长,希瓦氏菌的细胞膜流动性增强,表明腐败希瓦氏菌逐渐适应了低温环境。
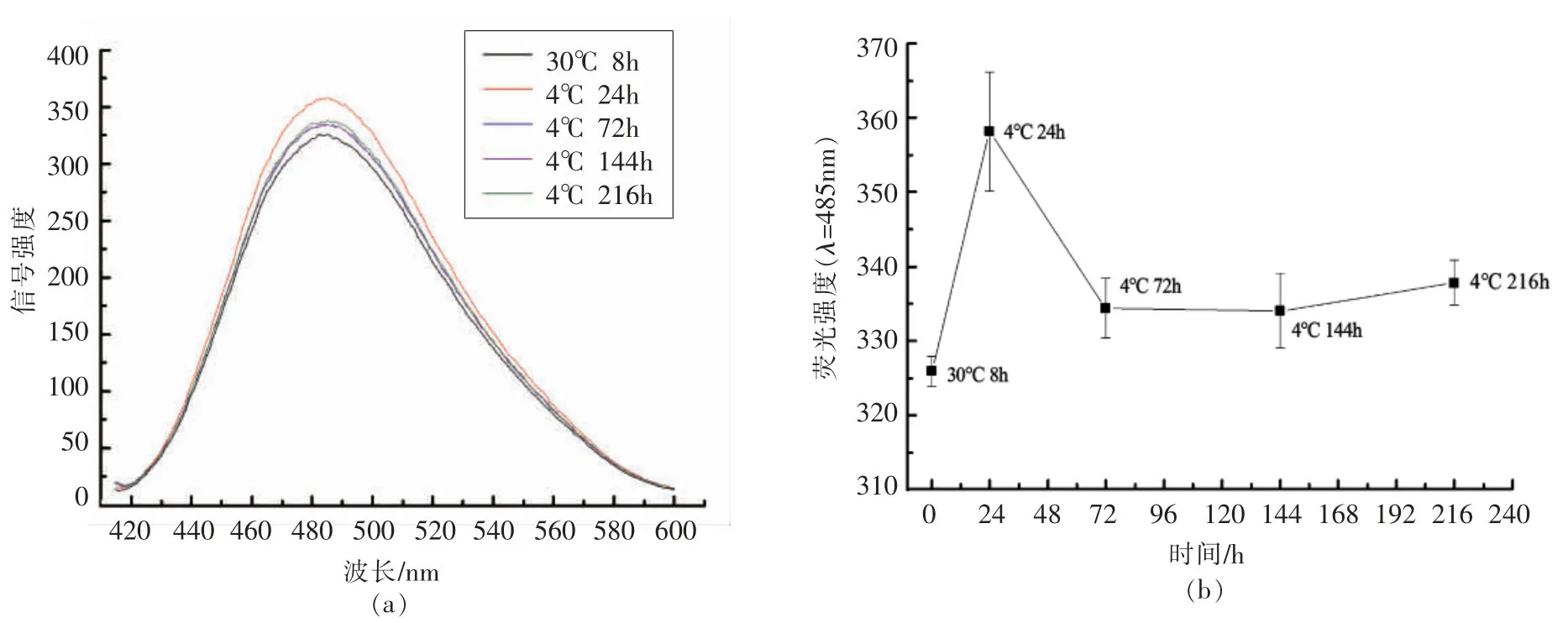
图7 腐败希瓦氏菌适冷过程中细胞膜流动性变化Fig.7 The changes in membrane fluidity of Shewanella putrefaciens during cold adaptation process
3 结论
对不同培养温度下腐败希瓦氏菌模式菌株DSM067生长情况及细胞膜理化特性的研究表明,腐败希瓦氏菌在低温胁迫下生长速率下降,细胞膜理化特性发生一定的变化。腐败希瓦氏菌模式菌株在30℃培养下经3.35 h延滞期后即进入快速增长的对数生长期,而10℃下延滞期为25.94 h,4℃培养下细菌经122.03 h才进入对数生长期;腐败希瓦氏菌最大比生长速率表明,4℃下μmax为0.017 h-1,显著低于30℃培养下的0.081 h-1。培养温度越低细菌生长繁殖越缓慢。腐败希瓦氏菌细胞膜脂肪酸组成以C16:0、C16:1和C18:0为主,随着培养温度的下降,单不饱和脂肪酸C16:1含量上升,其含量由30℃的26.62%升至4℃培养的46.66%。C16:1在腐败希瓦氏菌适冷过程中维持细胞膜流动性方面起关键性作用。不同培养温度下的腐败希瓦氏菌膜蛋白质表达出现差异,随着培养温度的降低,分子质量在30 ku及60~120 ku间的细菌蛋白表达量增多。腐败希瓦氏菌细胞TEM显微结构显示:30℃培养下的细菌细胞壁膜呈现波浪状褶皱,细胞膜表面积大,而培养温度下降,细胞壁膜较为平滑。细胞中出现的空泡从一定程度上反映细胞膜的流动性,30℃最适培养温度下细菌细胞膜流动性较好。对细菌细胞膜流动性的研究发现,腐败希瓦氏菌从最适的30℃培养转入4℃培养环境,细菌的膜流动性明显下降,然而经过一定时间的适冷,细菌细胞膜流动性得到一定的恢复,表明腐败希瓦氏菌经自身理化特性的变化逐渐适应了低温环境,从而能够继续生长繁殖。
猜你喜欢
中国实验诊断学(2022年5期)2022-05-27
福建轻纺(2021年12期)2021-12-16
实用老年医学(2021年12期)2021-12-05
心肺血管病杂志(2020年5期)2021-01-14
食品安全导刊(2020年8期)2020-08-16
食品安全导刊(2020年22期)2020-08-12
读与写(2019年31期)2019-11-26
食品安全导刊(2018年36期)2018-05-25
科学中国人(2015年13期)2015-02-28
食品工业科技(2014年23期)2014-03-11
