
母仔猪精氨酸的生理代谢与营养作用
2020-03-27 08:17徐运杰
猪业科学 2020年1期
徐运杰
(山东和美集团有限公司,山东 惠民 251700;唐人神集团股份有限公司,湖南 株洲 412000)
1 母仔猪精氨酸的生理代谢
1.1 合成代谢
20 世纪90 年代初,人们认识到,以前的养猪研究人员未能确定母猪奶水中所有生理上重要的氨基酸(如谷氨酰胺、瓜氨酸和鸟氨酸)。因此,在泌乳的第1 ~28 天,人们开始对母猪初乳和乳汁中游离和结合氨基酸进行定量研究。与泌乳母猪血浆中0.3 ~0.4 mM 谷氨酰胺相比,奶水中游离谷氨酰胺的浓度随着泌乳时间的延长而逐渐增加,在泌乳的第28 天达到3.5 mM 的最高平均值。相比之下,在整个哺乳期,母猪乳汁中精氨酸(Arg,游离加结合)的浓度远远低于谷氨酰胺加谷氨酸、脯氨酸、赖氨酸和支链氨基酸的浓度(表1)。母猪初乳或乳汁中游离精氨酸的浓度少于总Arg含量的0.7%。以克为基础,母猪在泌乳第7 天的奶水中Arg/Lys 的比值为0.35,而7 日龄仔猪的组织蛋白中Arg/Lys 的比值为0.97,表明母乳中缺乏精氨酸来满足仔猪最大生长的需求[1,2]。仔猪组织蛋白中精氨酸的高丰度和母猪奶水中精氨酸的明显缺乏引发了人们以下思考:1)母猪奶水中真的缺乏精氨酸?2)母猪奶水中高含量的谷氨酰胺和脯氨酸是否有营养意义?
研究表明,猪肠上皮细胞能够利用谷氨酰胺合成瓜氨酸和精氨酸。对14 ~58 日龄猪的空肠动脉和空肠静脉进行插管研究表明,小肠能够积极利用日粮和动脉血中的谷氨酰胺大量合成瓜氨酸[3]。为了确定负责瓜氨酸和精氨酸合成的细胞 类 型,Wu 等(1994)[4]从1 ~58 日龄猪小肠中分离出具有生化活性的肠上皮细胞,将谷氨酰胺的胞外浓度从0.5 增加到5 mM,研究发现呈剂量依赖性地增加猪肠上皮细胞通过吡咯-5-羧酸(P5C)合成酶合成瓜氨酸和精氨酸。与Blachier 等(1993)[5]的研究结果一致。合成途径所需底物包括氨、谷氨酸、天冬氨酸和ATP,都是由谷氨酰胺分解代谢产生。吡咯-5-羧酸合成酶和N-乙酰谷氨酸合成酶是由谷氨酰胺转瓜氨酸的两种关键调节酶[6]。断奶前和断奶后猪的其他主要组织(包括肝脏、肾脏、心脏、胰腺、大脑、大肠和骨骼肌)缺乏从谷氨酰胺或谷氨酸合成瓜氨酸的P5C 合成酶[7]。这些发现为肠上皮细胞从谷氨酰胺合成瓜氨酸和精氨酸奠定了基础。由谷氨酰胺生成瓜氨酸的反应始于肠上皮细胞线粒体,第一种酶是谷氨酰胺酶,将谷氨酰胺水解为谷氨酸,随后瓜氨酸转化为精氨酸在胞浆中进行[8]。值得注意的是,与线粒体中产生的鸟氨酸相比,细胞外鸟氨酸是肠上皮细胞中合成瓜氨酸和精氨酸的非常差的底物,这可能是由于细胞外鸟氨酸优先进入脯氨酸的生产[9]。这些结果表明:1)谷氨酰胺在瓜氨酸和精氨酸的肠道合成中起着重要作用;2)乳汁中谷氨酰胺的含量对弥补仔猪最佳生长所需的日粮精氨酸供给不足具有重要的营养意义。
猪肠细胞能够利用脯氨酸合成瓜氨酸和精氨酸。与0 ~7 日龄仔猪相比,14 ~21 日龄仔猪利用谷氨酰胺合成瓜氨酸的速率显著降低[10]。脯氨酸是哺乳仔猪肠上皮细胞合成精氨酸的替代前体物质。用鸟氨酸转氨酶抑制剂加巴曲林(gabaculine)处理母猪喂养的仔猪后,血浆中谷氨酰胺和脯氨酸的浓度增加了2 倍,提示脯氨酸可能是瓜氨酸合成的底物[11]。使用放射化学和色谱法研究发现,脯氨酸通过猪肠上皮细胞广泛降解,通过脯氨酸氧化酶途径产生P5C、瓜氨酸和Arg[12]。脯氨酸转化为瓜氨酸需要氨和谷氨酸,两者都是由谷氨酰胺降解提供的。由于猪小肠对动脉血中脯氨酸的摄取量很小,因此,猪奶水为肠内提供大量脯氨酸对奶水中精氨酸缺乏的补偿作用至关重要[3]。Brunton等(1999)[13]也研究报道,仔猪动脉血中脯氨酸合成精氨酸的作用很小,经胃给予脯氨酸可有效改善新生儿精氨酸缺乏。
另外,尽管精氨酸是通过尿素循环在肝脏中形成的,但在哺乳动物(包括猪)中,这一器官并没有精氨酸的净合成,因为胞浆精氨酸酶活性极高,能迅速水解Arg[8]。谷氨酰胺和脯氨酸在肠上皮细胞中形成瓜氨酸和精氨酸,在肾脏中瓜氨酸经精氨琥珀酸合成酶和精氨琥珀酸裂解酶(ASL)转化为精氨酸,这被称为从头合成Arg 的肠-肾轴。包括猪在内的许多哺乳动物物种中都存在这种机制,这是精氨酸肠外合成的主要部位。一些哺乳动物(如猫)肠道内缺乏瓜氨酸的合成,导致它们对膳食中精氨酸的需求很高[14]。
出生时,ASL 活性在肠上皮细胞中很高,但在肾脏中很低,因此,大多数谷氨酰胺和脯氨酸代谢合成的瓜氨酸能够被1 ~7 d 猪的肠上皮细胞用于合成精氨酸[10]。1 ~21 d 断奶仔猪肠道缺乏精氨酸酶,限制了精氨酸降解,有助于新生儿小肠Arg 的最大化[15]。Wu 等(1995)[10]估计,母猪奶水最多只能提供7 日龄仔猪所需精氨酸的40%。因此,通过肠-肾轴的Arg 合成在母猪喂养的仔猪维持Arg的稳态起着至关重要的作用。为支持这一观点,抑制肠道鸟氨酸转氨酶12 h,可使4 日龄母猪喂养的仔猪血浆中鸟氨酸、瓜氨酸和精氨酸的浓度分别降 低59%、52%和76%[11]。Wilkinson等(2004)[16]研究报道,给7 日龄仔猪喂食高精氨酸日粮(高精氨酸与精氨酸是一对同源氨基酸),与低精氨酸对照组相比,降低了脯氨酸对精氨酸的内源性合成,但这一发现不应解释为在生理范围内增加精氨酸的摄入量会降低瓜氨酸和精氨酸的肠道合成,因为喂食高精氨酸日粮(生长下降17%)的仔猪可能因氨基酸的严重不平衡而受损。
断奶后短期皮质醇升高能够诱导猪肠上皮细胞P5C合成酶的表达,促进谷氨酰胺合成瓜氨酸[17]。在断奶后的猪中,ASL 活性在肾脏中高,而在肠上皮细胞中低[10,12]。因此,小肠将大部分合成的瓜氨酸释放到血液循环中。在断奶前和断奶后的猪中,小肠释放的瓜氨酸没有被肝脏提取,而是主要用于肾脏中的精氨酸合成。由于肝细胞中氨基酸转运系统y+(一种碱性氨基酸转运体)的活性较低,限制了猪肝脏对生理浓度Arg 的摄取,因此,小肠利用谷氨酰胺和脯氨酸内源合成精氨酸对猪至关重要[18]。谷氨酰胺和脯氨酸在肠道中生成的瓜氨酸和精氨酸作为精氨酸的来源对整个身体同样有效(图1)。Wu 等(2016)研究报道,日粮中添加0.5%至2%精氨酸,不会影响断奶后仔猪利用谷氨酰胺和脯氨酸肠道合成瓜氨酸和精氨酸[19]。
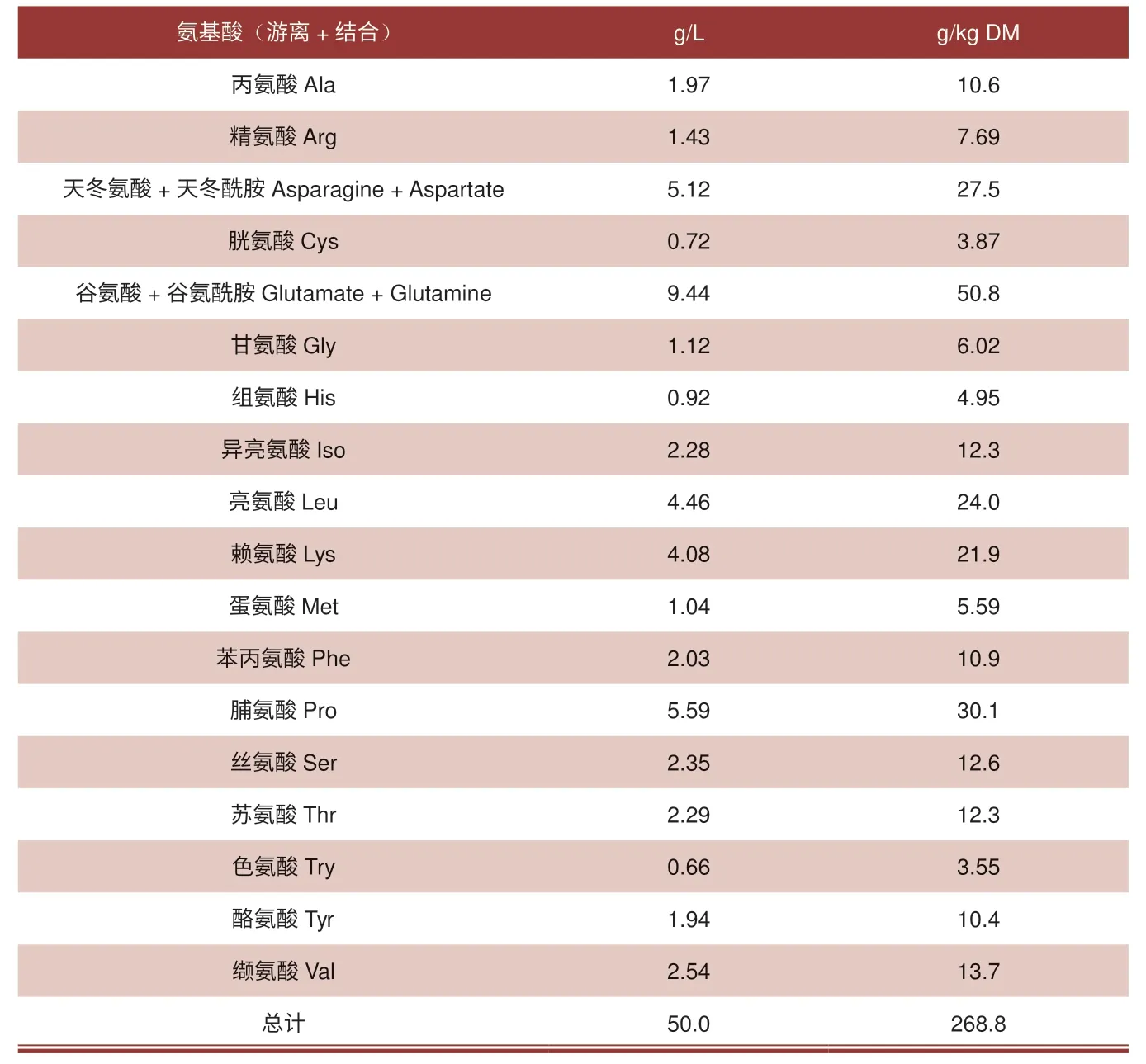
表1 泌乳第7 ~21 天母猪全乳中氨基酸的含量
很多因素,包括遗传、营养和应激(如拥挤、疾病、环境温度和空气污染)都会影响仔猪的生长。人工饲养系统的研究数据表明,新生仔猪的生长潜力至少为400 g/d,或大于母猪喂养仔猪(230 g/d)的74%以上,哺乳仔猪出生后第8 天开始出现亚最大生长[21]。有趣的是,哺乳仔猪的亚最大生长发生在线粒体中N-乙酰谷氨酸合成酶相对缺乏时,肠上皮细胞合成的瓜氨酸和Arg 显著减少[6,10]。与新生仔猪相比,7 日龄乳猪以谷氨酰胺为原料肠道合成瓜氨酸和精氨酸减少了70%~73%,肠上皮细胞以脯氨酸为原料合成瓜氨酸和精氨酸的速率也降低了75%~88%,14 ~21 日龄进一步减少[12]。说明哺乳期仔猪内源性Arg 合成显著减少导致Arg 的缺乏限制了母猪喂养仔猪的最大生长。
1.2 分解代谢
作为调节精氨酸稳态的一种机制,猪精氨酸分解代谢主要通过精氨酸酶途径发生,并随着日粮精氨酸摄入量的增加而逐渐增加。根据7 日龄仔猪尿液中肌酸酐的排泄量,Wu 等(2004)[6]估计,新生仔猪每天每千克体重消耗70 mg 精氨酸合成肌酸,占体内分解Arg 的21%或母乳提供Arg 的17%。类似地,Brosnan 等(2009)[22]报告,仔猪在4 ~11 日龄期间每天每千克体重消耗110 mg 精氨酸合成肌酸。对于4 月龄的猪,每天每千克体重消耗84 mg 精氨酸产生肌酸[23]。尽管高精氨酸是由赖氨酸分解代谢或其前体Arg 的氨基转移合成,但该途径每天每千克体重仅消耗0.036 mg 精氨酸,在数量上无关紧要。
泌乳母猪乳腺组织中精氨酸的分解代谢也非常可观。乳腺吸收了大量的精氨酸,但乳汁中精氨酸的含量却远低于乳腺组织对精氨酸的吸收[24]。为了理解这一点,O'Quinn等(2002)[25]利用泌乳母猪的乳腺组织进行代谢研究,发现该组织能主动降解精氨酸,形成脯氨酸、多胺和NO。精氨酸酶的2 个亚型(I型和Ⅱ型)负责精氨酸水解产生尿素和鸟氨酸,随后通过鸟氨酸转氨酶和P5C 还原酶转化为脯氨酸。但是,由于缺乏脯氨酸氧化酶,脯氨酸的分解代谢在猪乳腺组织中没有检测到。这些发现有助于我们解释为什么精氨酸在母猪的乳汁中非常缺乏,而脯氨酸却非常丰富。
断奶前猪的肠上皮细胞几乎不表达精氨酸酶,因此不能从精氨酸合成脯氨酸。这有助于解释为什么脯氨酸是幼猪必需的营养氨基酸。在断奶期间,皮质醇的激增诱导了仔猪的肠道表达精氨酸酶[26]。此后,生长育肥猪和成年猪肠上皮细胞中精氨酸酶的活性持续升高[19]。断奶后猪的肠上皮细胞表达I 型和Ⅱ型精氨酸酶,能够积极地将精氨酸降解为鸟氨酸和尿素,鸟氨酸转氨酶和P5C 还原酶进一步将鸟氨酸转化为脯氨酸[15]。猪小肠管腔中的细菌也会将一些日粮中的精氨酸代谢为鸟氨酸,可能还有少量短链脂肪酸[27]。因此,胃肠道微生物群可能影响Arg 的利用率和宿主健康。仔猪断奶后,精氨酸的肠道分解代谢减少了日粮精氨酸进入门静脉的数量。Wu 等[19,28](2007 和2016)估计,大约40%的口服精氨酸(通过分解代谢和蛋白质合成)被生长猪和成年猪的小肠利用,其余部分精氨酸进入门静脉。在肠外组织中,精氨酸是由精氨酸酶、精氨酸;甘氨酸氨基转移酶、精氨酸脱羧酶和一氧化氮合成酶启动进入分解代谢途径,这些途径分别用于合成鸟氨酸、肌酸、精胺和一氧化氮(图2)。在胃肠道中,由胞外蛋白酶和肽酶水解日粮蛋白质释放出精氨酸、其他氨基酸和小肽进入肠腔。肠细胞吸收这些消化产物,降解精氨酸,释放瓜氨酸。氨基酸的分解代谢也由肠道内的细菌发生。在断奶后的猪和大鼠中,在第1 次进入门静脉时,大约40%的Arg 和15%(例如色氨酸)至95%(例如谷氨酸)的其他氨基酸在小肠内腔中通过分解代谢和蛋白质合成被利用。因此,小肠内腔中有5%(例如谷氨酸)到85%(例如色氨酸)的氨基酸进入门静脉。
在新生仔猪肠上皮细胞中发现瓜氨酸和精氨酸合成,那么,这一途径是否可能在胎猪中就开始发生?通过分析妊娠不同时期猪的胎液发现,与母体血浆水平(0.1 ~0.14 mM)相比,妊娠第40 天猪尿囊液中的精氨酸含量异常高(4 ~6 mM)。此外,在妊娠第40 天,与母体血浆水平(鸟氨酸为0.05 ~0.1 mM,谷氨酰胺为0.3 ~0.45 mM)相比,猪尿囊液中的鸟氨酸(1 ~3 mM)和谷氨酰胺(3 ~4 mM)浓度也特别高[29-30]。值得注意的是,猪尿囊液中精氨酸、鸟氨酸和谷氨酰胺的浓度在妊娠第30~40 天之间分别增加了23、18 和4倍,它们的氮占游离α 氨基酸总氮的67%。胎液中精氨酸的异常丰度有利于胎盘中NO 和多胺的合成。这些新发现为精氨酸依赖性代谢途径在胚胎存活、生长和发育中的关键作用提供了生理基础[31]。
2 母仔猪精氨酸的营养作用
很多研究结果表明,在传统日粮中添加精氨酸可以改善现代猪品种在妊娠期、哺乳期、保育期、断奶期和生长发育期的生长、生产性能和健康状况。2012 年,NRC 认识到精氨酸在猪生产的各个阶段都是条件性必需氨基酸。NRC(2012)[32]推荐了在妊娠、哺乳、保育、断奶和生长肥育猪日粮中精氨酸的最低含量(表2)。
2.1 新生仔猪精氨酸的营养作用
仔猪的生长对日粮中精氨酸的供应非常敏感,精氨酸的严重缺乏(例如仔猪血浆中精氨酸浓度≤26μM)将迅速导致高血氨症,甚至新生儿死亡。如前所述,母猪的乳汁中精氨酸含量相对较低。因此,日粮中添加0.2%和0.4%精氨酸至7 ~21 d乳猪的液态奶中进行人工饲养,发现可剂量依赖性地增加血浆中精氨酸浓度(30%和61%),降低血氨浓度(20%和35%),并增加仔猪的体重增长(28%和66%)[1]。最近,Yang 等(2016)[34]报道,在代乳日粮中添加0.4%或0.8%精氨酸可使4 ~24 d 仔猪的体重分别增加19%和22%,而不影响饲料摄入量。有趣的是,在断奶前日粮添加精氨酸对25 ~45 d 仔猪的肠道生长发育有促进作用。张莉莉等(2017)[35]研究了精氨酸对子宫内发育迟缓(IUGR)仔猪抗氧化功能的影响,结果表明,日粮补充Arg 显著降低IUGR 仔猪背最长肌和空肠黏膜丙二醛含量;显著提高空肠黏膜总抗氧化能力和谷胱甘肽过氧化物酶活性;显著提高血清和腔肠黏膜中的NO 水平、总NO 合成酶、诱导型NO 合成酶、结构型NO 合成酶活性,以及空肠黏膜与血清中Arg 水平和Arg 代谢相关的氨基酸水平。说明在早期断奶IUGR 仔猪日粮中添加精氨酸具有增强IUGR 仔猪抗氧化功能和促进精氨酸代谢的作用。
2.2 断奶仔猪精氨酸的营养作用
仔猪可有效利用日粮瓜氨酸合成精氨酸,但日粮鸟氨酸对体内精氨酸合成的影响不大。在喂食含13.1%粗蛋白、0.89%赖氨酸和0.18%精氨酸的半纯合日粮的断奶仔猪中,与对照组比较,添加0.18%精氨酸或等摩尔量瓜氨酸的试验组增加了血浆中精氨酸的浓度和生长性能,但在基础日粮中添加等摩尔量的鸟氨酸对血浆精氨酸的浓度和生长性能都没有影响[36]。
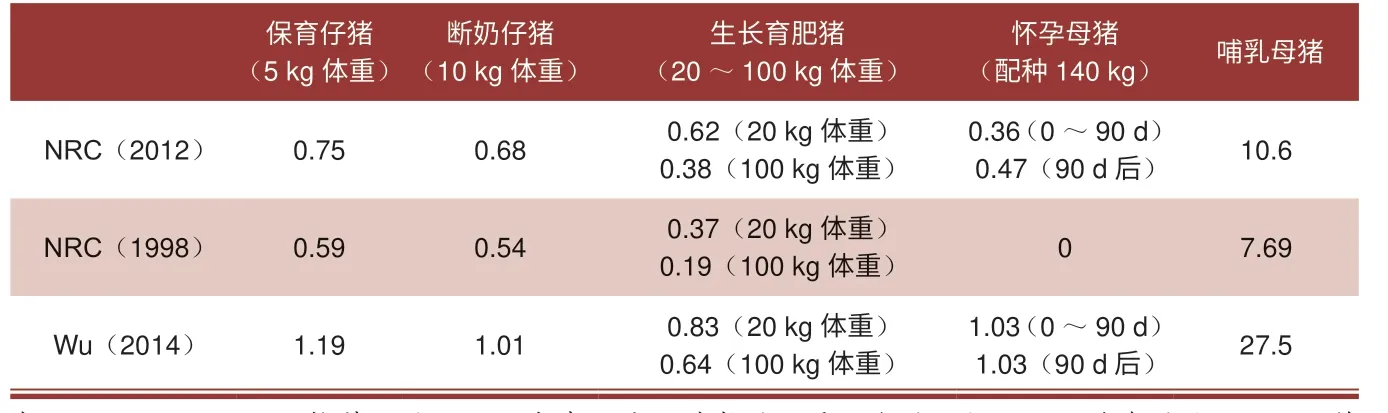
表2 NRC 推荐的猪日粮中精氨酸的最低含量和Wu 推荐的猪日粮中精氨酸的最佳含量(数值为总日粮的百分比)
与猪乳一样,断奶仔猪的典型玉米-豆粕型日粮(不添加鱼粉或血浆)中精氨酸的含量相对较低。另外,与哺乳仔猪相比,断奶仔猪在断奶后第1 周的采食量通常降低,导致日粮氨基酸(如谷氨酰胺、谷氨酸和脯氨酸)摄入不足,加上线粒体中N-乙酰谷氨酸合成酶活性低,双重影响了精氨酸的内源合成,导致断奶仔猪精氨酸缺乏,从而极大地限制了断奶仔猪的生长。在以玉米、豆粕为基础的日粮中添加0.6%精氨酸,可使断奶仔猪的小肠重量和日增重分别增加22%和145%[37]。
2.3 妊娠母猪精氨酸的营养作用
现代高产母猪每次排20 ~30个卵细胞,最终产10 ~15 头甚至更多的活仔猪。大量研究报告证实,精氨酸对胎盘重量、胚胎和胎儿存活、仔猪出生重等方面具有重要生理作用。第一,在实际生产条件下,在妊娠第22 ~114 天期间,饲喂补充0.83%精氨酸日粮的母猪,其胎盘重量增加16%、每窝产活仔猪数增加1.1 头和窝出生重增加1.7 kg[38]。第二,在实际生产条件下,在妊娠第14 ~28 天期间的母猪日粮中添加1%的精氨酸,每窝产活仔猪数增加约1 头[39]。第三,与对照组母猪相比,在妊娠第14 ~25 天之间的母猪日粮中添加0.4%或0.8%的精氨酸,可使胎盘生长增加21%~34%,每胎产仔数增加约2 头,值得注意的是,0.4%和0.8%精氨酸组之间的胚胎存活率没有差异[40]。郭鹏等(2017)[41]研究表明,精氨酸能通过其代谢产物NO 和腐胺激活β-catenin 和mTOR 信号通路,促进母猪胎盘滋养层细胞的增殖。第四,在妊娠第14 ~28 天期间母猪日粮中添加0.9%精氨酸盐酸盐,能够使超数排卵母猪在妊娠第70 天的胎儿数增加3.7 头,并使后代的出生体重、骨骼肌和器官重量以及肌纤维增生增加[42]。第五,在妊娠第90 ~114 天期间的母猪日粮中添加0.83%精氨酸,可使仔猪的平均出生体重提高16%[43]。
2.4 泌乳母猪精氨酸的营养作用
生理水平的NO 能够促进哺乳动物(如母猪)乳腺的血液流动、产奶以及乳腺上皮细胞的蛋白质合成[44-45]。精氨酸的供应增加了乳腺组织中NO 和多胺的产生,因此,它促进了乳汁的生成[25,44]。在初产母猪日粮中添加0.83%精氨酸(即1%精氨酸盐酸盐)可使初产母猪在泌乳第一周的窝增重和产奶量分别 提 高21%和11%[46]。Wu 等(2006)[47]对初产母猪饲喂添加或不添加精氨酸的日粮,比较了低出生体重和正常出生体重仔猪0~14 d 的生长性能,结果表明,精氨酸能够使低出生体重仔猪的日增重比正常出生体重仔猪的大。此外,哺乳母猪日粮中添加精氨酸可提高奶水中脂质的生成[48]。Laspiur 等(2001)[49]报道,在泌乳母猪日粮中添加精氨酸降低了其体重损失,提高了饲料效率,特别是在炎热的环境中,添加精氨酸有利于改善母猪的心跳和呼吸速率以及仔猪的增重。由此可知,泌乳期日粮中添加精氨酸对改善母仔猪的健康状况以及仔猪的生长和存活具有重要的营养、生理和现实意义,特别是对于那些经历过宫内生长受限的仔猪。
3 日粮补充精氨酸的安全性
猪日粮中添加精氨酸并没有影响血浆中赖氨酸或组氨酸的浓度(表明碱性氨基酸之间没有颉颃作用),但有益于增加瘦肉组织质量和减少白脂肪,以及血浆中氨、游离脂肪酸、三酰甘油酯和胆固醇的浓度[50]。Hu等(2015)[51]评估了长期补充精氨酸盐酸盐或精氨酸对30 ~121 日龄猪的安全性,结果表明,日粮中添加0%、1%、1.5%或2%精氨酸,血液生化结果(包括红细胞和白细胞的数量、血红蛋白和pH、血浆中总胆红素、氨基酸、葡萄糖、尿素、氨、肌酐、脂质和矿物质)均在生理范围内,没有受到添加精氨酸的负面影响。但是,与其他营养素一样,过量摄入精氨酸会对动物产生不良影响。在以玉米、豆粕为基础的日粮中添加4%精氨酸会对仔猪产生不利影响,表现为氨基酸失衡和生长性能下降。精氨酸和赖氨酸的最佳日粮比例可能随猪的年龄和健康状况而变化。日粮中精氨酸与赖氨酸的比例大于3:1 时,断奶仔猪的采食量和生长性能降低[20]。
4 结语
总之,精氨酸在猪的营养中是一种功能性和条件性必需氨基酸。大量的研究已经确认精氨酸是妊娠期猪体内及其胎儿体内最丰富的氨基酸之一。日粮中约40%的精氨酸在第1 次通过小肠时被分解,通过氨基酸的器官间代谢进行内源性合成对维持精氨酸在全身的稳态至关重要(图1)。在细胞水平上,精氨酸对蛋白质和其他含氮物质(包括NO、肌酸、多胺和高精氨酸)的合成至关重要,这些物质在体内具有关键的代谢功能。因此,精氨酸对改善猪的健康、存活、生长、发育、泌乳和繁殖性能具有重要作用。精氨酸既是哺乳仔猪和断奶仔猪的功能性必需氨基酸,也是支持母猪最大繁殖和泌乳性能的条件性必需氨基酸。日粮中添加0.5%~1%精氨酸可以改善母仔猪的生产性能和繁殖性能。
猜你喜欢
中老年保健(2022年3期)2022-11-21
体育科技文献通报(2022年4期)2022-10-21
现代妇产科进展(2022年9期)2022-10-07
中国动物保健(2021年8期)2021-11-24
安徽化工(2021年3期)2021-05-29
特产研究(2021年2期)2021-04-13
天津医科大学学报(2021年2期)2021-03-29
岭南现代临床外科(2020年5期)2020-12-13
现代农业科技(2020年10期)2020-06-04
科学与财富(2019年15期)2019-10-21
