
甘蓝型油菜BnHY5基因的表达模式及非生物胁迫响应分析
2020-04-01 09:02郭媛媛王茂林
四川大学学报(自然科学版) 2020年2期
方 艳, 岳 出, 李 洁, 郭媛媛, 王茂林
(四川大学生命科学学院 生物资源与生态环境教育部重点实验室, 成都 610064)
1 引 言
甘蓝型油菜(BrassicanapusL.)是世界上重要的油料作物之一[1]. HY5转录因子对植物众多生理过程具有调节作用[2],属于亮氨酸拉链(bZIP)家族[3]. 该家族转录因子参与植物对病原体的防御过程,花的发育和种子的成熟以及光和激素信号传递及反应过程等[4].植物bZIP类的蛋白在结合表达基因的DNA序列时,会优先与包含特定作用元件的DNA序列结合,因此植物bZIP类蛋白会偏向于优先识别有G-box (CACGTG)、 A-box (TACGTA)、C-box (GACGTC)等原件的序列[5].碱性亮氨酸拉链基本结构是以亮氨酸残基开头每6个氨基酸形成的7肽单元;多个七肽单元可稳定二聚化[6].拟南芥中bZIP家族有78个成员,这些转录因子依据功能和结构域特点被细分为十三组,目前为止,这些成员的其中四十多个尚未做系统的功能和序列的分析[7].HY5及其同源基因HYH都含有COP1(Constitutive Photomorphogenesis1)相互作用结构域并有相似的功能,被单独分到了的一组,二者常常被一起研究.拟南芥HY5突变体在光照条件下具有细长的下胚轴的表型,在该突变体中过表达功能区完整的HYH可以弥补长的下胚轴表型[8],说明拟南芥HY5基因与其同源基因HYH在功能上冗余.拟南芥中光形态建成应答调控机制的研究揭示了HY5在光应答中的中关键的调控作用[2],HY5调控着基因组中上千个光诱导基因的表达[9].近年来相关研究发现,HY5不仅参与光信号响应,还可能参与非生物胁迫等过程[10-11],HY5转录因子位于调控的中心.但目前针对甘蓝型油菜BnHY5基因的研究较少,对非生物胁迫方面的研究也未见报道.基于该转录因子的重要作用,在甘蓝型油菜中研究其表达模式是非常必要的.本文研究分析了油菜BnHY5表达模式及胁迫逆响应,为进一步认识其功能与油菜遗传改良奠定基础.
2 材料与方法
2.1 材 料
甘蓝型油菜(BrassicanapusL.)科乐油1号种植于植物光照培养室.
2.2 方 法
2.2.1 RNA提取和cDNA合成 油菜总RNA的提取用植物总RNA提取试剂盒(E.Z.N.A R6627-02Plant RNA Kit OMEGA公司),操作按照提取试剂盒说明书进行.提取高质量的RNA后立即进行cDNA的合成.总RNA反转录使用反转录试剂盒(ONE-Step-gDNA Removal and cDNA Synthesis SuperMix 试剂盒;TRANSGENE公司).cDNA合成后于-20 ℃保存备用.(体系为20 μL,其中总RNA含量在500 ng~5 μg).
2.2.2 甘蓝型油菜BnHY5基因全长cDNA的克隆 以Genbank中已经公布的甘蓝型油菜(BrassicanapusL.)数据库中预测的HY5和HYH各个基因开放阅读框序列同源部分设计特异性引物.
以cDNA为模板进行PCR扩增,扩增产物用凝胶电泳检测,PCR产物经纯化后连接T克隆载体,蓝白斑筛选后挑取单菌落进行重组子鉴定,将阳性克隆扩大培养后提取质粒,送至华大基因有限公司进行测序.引物序列为见表1.
2.2.3BnHY5基因序列分析 目的基因测序结果用DNAMAN软件和NCBI中的BLAST工具进行比对分析.BnHY5蛋白理化性质利用在线软件Expasy-Protparam分析[15]和NCBI在线工具完成,使用SOPMA对BnHY5蛋白二级结构进行预测[12],而蛋白三级结构模型用Swiss Model在线工具分析.使用使用邻接法推断了进化历史,MEGA-X软件构建系统发育进化树[13].
2.2.4BnHY5 基因在各时期各器官的表达模式分析及胁迫处理 分别取光照和黑暗下的幼苗材料,苗期(两叶一心期、四叶一心期)、抽薹期、花期的根、茎、叶材料.取样后于-80 ℃冰箱保存备用.油菜盆栽培养至2~3片真叶,将油菜根部泥土洗净置于1/2霍格兰溶液中继续培养.培养至3~4片真叶进行胁迫激素处理和胁迫处理.激素处理为150 μmol/L ABA和GA3,分别添加0.05%的乙醇和0.01%的Tween-20表面活性剂[1]喷施叶面,空白对照喷施含同浓度乙醇和表面活性剂的清水.将甘蓝型油菜幼苗分别放置40 ℃光照培养箱做高温处理,和4 ℃冷室内做低温处理,正常光照;盐胁迫,镉胁迫处理分别在培养液中添加含150 mmol/L的氯化钠,150 μmol/L氯化镉,未经过任何处理的材料作为空白对照.所有材料在处理后,0、3、6、9、12、24和48 h(高温胁迫、低温胁迫处理后每隔2 h)进行一次取样.取生长状况一致的幼苗地上部分,液氮冷冻后于-80 ℃保存.每次实验重复3次,实验结果是3次重复实验结果的平均值.
2.2.5BnHY5基因在逆境胁迫和外源激素处理下表达模式分析BnHY5基因的qPCR预实验确定了定量的最佳退火温度(56 ℃)和模板浓度.以甘蓝型油菜管家基因β-actin(GenBank登录号AF111812)为内参[1],设计特异性荧光定量引物,qPCR引物序列见表2.

表2 qPCR引物
qPCR检测BnHY5基因在不同胁迫处理后不同时间段的表达情况.荧光定量PCR仪为美国Bio-Rad CFX96[1].荧光定量试剂盒Trans Start Tip Green qPCR Supper Mix(TRANSGENE生物公司).qRT-PCR反应体系为20 μL:10 μL 2× Master Mix;1 μL模板cDNA(0.3 μg/μL);正反向引物各0.4 μL(10 μmol/L);8.2 μL 的dd H2O.qRT-PCR采用三步法,反应程序为:94 ℃预变性30 s;42个循环(94 ℃ 5 s;56 ℃ 15 s;72 ℃ 10 s)同时绘制熔解曲线.反应结束后,目标基因的相对定量用2-ΔΔCt法[14]计算,每个样品都进行3次重复实验.数据采用SPSS 20进行统计分析.
3 结果与分析
3.1 BnHY5基因全长的克隆与编码蛋白质分析
BnHY5及其同源基因PCR扩增见图1(a)(b).测序结果利用NCBI在线工具Blastn比对,成功获得HY5基因及同源基因HYH,命名为BnHY5和BnHYH.并且在油菜中它们存在多个基因,BnHY5有四个基因HY5-1(BnC09HY5.1;795bp);HY5-2(BnA10HY5;825bp);HY5-3(BnA02HY5;738bp);HY5-4(BnC03HY5.2;752bp).而BnHYH克隆到三个基因HYH-1,HYH-2,HYH-3.发现BnHY5-1与甘蓝型油菜预测转录因子HY5基因(LOC106391328)核苷酸序列相似性为99%.与拟南芥的HY5序列相似性为83.08%,与甘蓝的HY5序列相似性为98%.油菜HY5转录因子氨基酸序列的N端有能与 COP1 相互作用的结构域,C端则是bZIP结构域.同时,油菜中HY5基因四个重复基因的COP1 interaction区域与其主要作用结构域亮氨酸拉链的氨基酸序列基本上一致.

(a)BnHY5

(b)BnHYH
1.BnHY5-1;2.BnHY5-2;3.BnHY5-3;4-5.BnHY5-4 1-2.BnHYH-1;3-4.BnHYH-2;5.BnHYH-3
图1 基因全长PCR扩增
M.DNA marker;number.PCR products
Fig.1 Full-length PCR amplification ofBnHY5 and BnHYH genes
拟南芥、甘蓝、芜菁HY5氨基酸序列与油菜HY5氨基酸序列比对见图2.与拟南芥HY5氨基酸序列相比,油菜的氨基酸序列存在四处氨基酸不同.另外,BnHY5氨基酸序列中部50个氨基酸处AtHY5氨基酸序列少了4个氨基酸RESG(精氨酸,谷氨酸,丝氨酸,甘氨酸)这四个氨基酸均为亲水性较强的氨基酸;并且在bZIP的LZ(亮氨酸拉链)基本结构区即七肽单元区末尾处存在个别氨基酸的差异,例如肽链50个氨基酸处拟南芥的氨基酸是缬氨酸,而油菜的却是苏氨酸.甘蓝型油菜与两个亲本甘蓝和芜菁的HY5氨基酸序列进行比对,发现蛋白序列76与158个氨基酸处,甘蓝型油菜氨基酸差异与亲本芜菁和甘蓝有着相同的特点:甘蓝的为G,芜菁的为S,油菜HY5-1的为G,HY5-2的为S(图2).

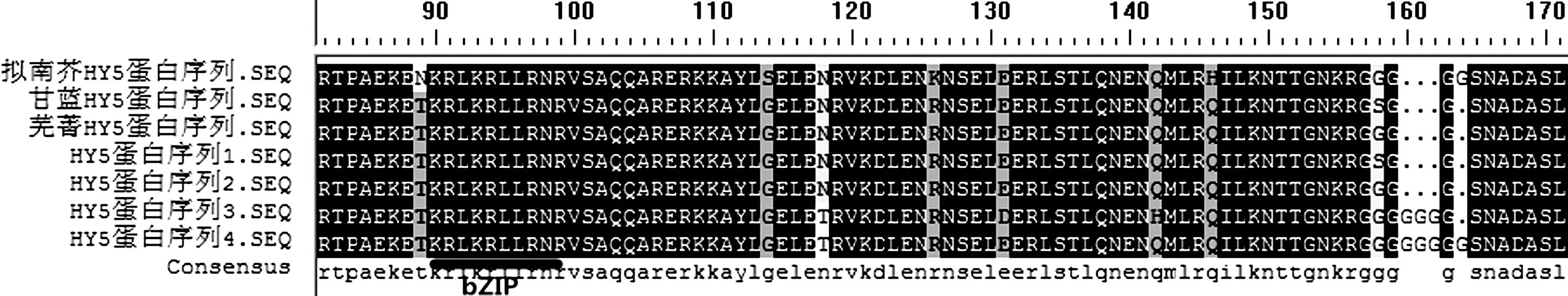
图2 拟南芥HY5蛋白与芜菁、甘蓝、甘蓝型油菜HY5蛋白氨基酸序列比对
Fig.2 Amino acid sequence comparison ofBnHY5 with arabidopsis AtHY5 and HY5 of brassica rape, brassica oleracea amino acid sequence
3.2 BnHY5蛋白理化性质分析与结构预测分析
3.2.1 BnHY5蛋白理化性质分析

表3 BnHY5蛋白的理化性质
BnHY5-1蛋白相对分子质量为18 037.82,不稳定指数为61.67,为不稳定类蛋白;等电点为9.29;亲水性的平均值(GRAVY)为-1.198.结果表明,该蛋白为不稳定、亲水性蛋白.
3.2.2 BnHY5蛋白质三级结构预测 BnHY5蛋白二级结构主要显示蛋白结构域主要是α-螺旋结构,另外在非结构域部分为无规则卷曲和片层结构,以及少量β-折叠.构象均存在亮氨酸拉链α螺旋以及enhancer-binding protein beta区域(CCAAT),进一步证明属于bZIP家族.BnHY5氨基酸序列预测得到三级结构构象有2种:BnHY5-1和BnHY5-4的构象如图B1,而BnHY5-2,BnHY5-3的构象如图B2;而拟南芥AtHY5转录因子氨基酸序列预测得到的三级结构构象有两个独立构象(图3),B3、B4为分开的两个独立构象,B5为合在一起的构象.根据BnHY5和AtHY5构象差异可以看出比拟南芥HY5能够产生的构象类型更为简单,推测甘蓝型油菜的HY5转录因子的作用功能更有可能与其他的转录因子相互作用以起到调控作用.


图3 BnHY5蛋白三级结构预测B1.BnHY5蛋白三级结构一类预测构象;B2.BnHY5蛋白三级二类预测构象;B5.AtHY5蛋白构象1和构象2共存时三级结构预测构象;B3,B4为AtHY5蛋白构象1和构象2;Fig.3 Prediction of tertiary structure of BnHY5 protein
3.3 BnHY5蛋白系统进化树的构建
利用MEGA-X软件[13]中Neighbor Joining算法构建系统进化树(图4),使用泊松校正方法计算进化距离,使用最大复合似然法计算进化距离,
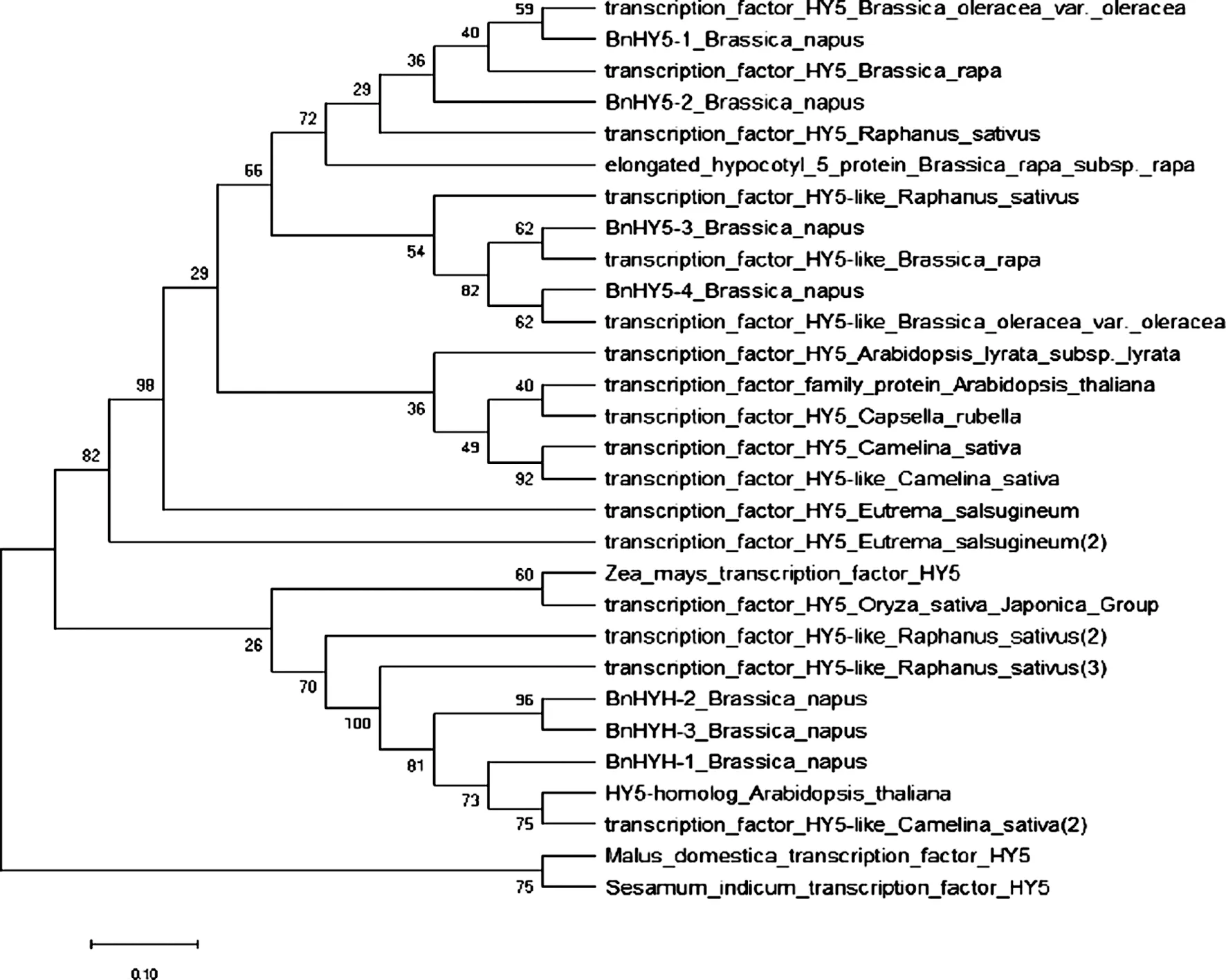
图4 BnHY5蛋白系统进化树分析标尺上数字代表碱基替换率,在分支旁边显示了相关分类群在引导测试中聚集在一起的进化树的百分比(1 000个重复).Fig.4 Evolutionary relationships of BnHY5 protein
并以每个位点的碱基取代数为单位[17]. BnHY5与双子叶植物十字花科物种中甘蓝和芜菁HY5的亲源关系最近,与芜菁的同源性为89%,与萝卜的为86.8%.与白菜等HY5的蛋白具有较高的同源性.与亚麻荠的亲源关系最远,蛋白同源性为42.86%.十字花科不同种属的植物的bZIP作用结构域是高度保守的.进一步说明bZIP家族蛋白的遗传进化是保守的.另外从进化树可以看出,甘蓝型油菜HY5与双子叶植物苹果、芝麻HY5的亲源关系较远;与单子叶植物水稻,玉米HY5的亲源关系又比双子叶植物的更远.
3.4 BnHY5基因在油菜不同生长期不同组织的表达分析
以β-actin为内参,油菜中各个器官中BnHY5基因的表达情况如图5(ss2、ss4表示苗期的二叶一心和四叶一心时期;bs表示抽薹期,fs表示花期).BnHY5基因在甘蓝型油菜整个生长周期中均要表达,但表达量存在差异.在油菜的营养生长期,叶中的表达量最高,根、茎中表达量递减,同一生长期茎中表达量最低.但在甘蓝型油菜进入生殖生长期后发生变化,叶片中的表达量随着植物体逐渐老化而逐渐降低.抽薹期根中的表达量显著升高(P<0.05);BnHY5-1花期茎中的表达量比抽薹期的高出1/3.在不断膨大的油菜角果中的表达量也比较高,花中的表达量与同时期叶中的表达量相当.

(a)BnHY5-1 (b)BnHY5-2

(c)BnHY5-3 (d)BnHY5-4
图5BnHY5基因在油菜不同发育时期不同器官的相对表达量
a,b,c表示P<0.05,同组数据间达到差异显著
Fig.5 Relative expression ofBnHY5 gene in different organs of different stages of rape
3.5 甘蓝型油菜BnHY5基因与同源基因BnHYH表达差异分析
光是影响HY5表达量变化的重要因素.分析甘蓝型油菜BnHY5与其同源基因BnHYH在光照和黑暗下的表达差异.以β-actin为内参基因,BnHY5-1的表达量为标准,BnHY5多个基因的表达量如图6.甘蓝型油菜HY5和HYH各基因光照下的表达量均高于黑暗下的表达量;但HYH各

图6 甘蓝型油菜幼苗光照与黑暗下重复基因的相对表达量
*:P<0.05,同个基因在光照下表达量与黑暗下的相比达到差异显著
Fig.6 Relative expression of repeat genes in light and dark conditions ofBrassicanapusL.
基因在光照和黑暗下的表达差异要比HY5基因更大;并且在黑暗下,HYH的表达量趋近于0,说明黑暗条件下油菜中主要发挥作用的是HY5.甘蓝型油菜幼苗时期BnHY5四个基因均有表达,但表达量存在差异.根据实验结果得知,无论是在黑暗还是光照下,HY5-2与HY5-3的表达量相当且高于其他基因的表达量,HYH-1和HYH-2的表达量高于其他基因的表达量.HYH与HY5多个基因在无光条件下的表达量低于光照条件下的表达量.表明BnHY5与BnHYH对光因子的响应存在相似的表达调控机制和一定的协同性.
3.5 外源激素和非生物胁迫处理下BnHY5-1的表达分析
外源激素处理下甘蓝型油菜BnHY5-1基因的相对表达量如图7所示,以0 hBnHY5-1在油菜中的表达量为标准,得到该基因在不同时间段甘蓝型油菜中的相对表达量.分析发现甘蓝型油菜在GA3处理后,BnHY5-1的表达下调,其3 h的表达量是0 h的1/10;暗示GA3是HY5的负调控因子.在 ABA处理后BnHY5-1表达上调,其3 h的表达量是0 h的9倍,在处理 3 h后其表达量迅速增加,处理9 h后其表达量恢复至与0 h的相当非生物胁迫处理下甘蓝型油菜BnHY5-1的表达情况如下:盐胁迫与镉胁迫处理后BnHY5-1表达量如图8(a),(b)所示,分析发现甘蓝型油菜在盐胁迫下,BnHY5-1的表达下调,暗示HY5是盐胁迫应答调控中的负调控因子.镉胁迫处理后,BnHY5-1表达上调,处理后 3 h的表达量是未处理 时的 3.2倍;低温胁迫处理后BnHY5-1表达上调,并且其表达量在很短的时间内都呈大幅度变化的趋势,处理2 h后其表达量是未处理时的 12倍,见图8(d).高温胁迫处理后,BnHY5-1表达上调,处理4 h后其表达量是未处理时的13倍,见图8(c),说明BnHY5-1响应非生物胁迫.
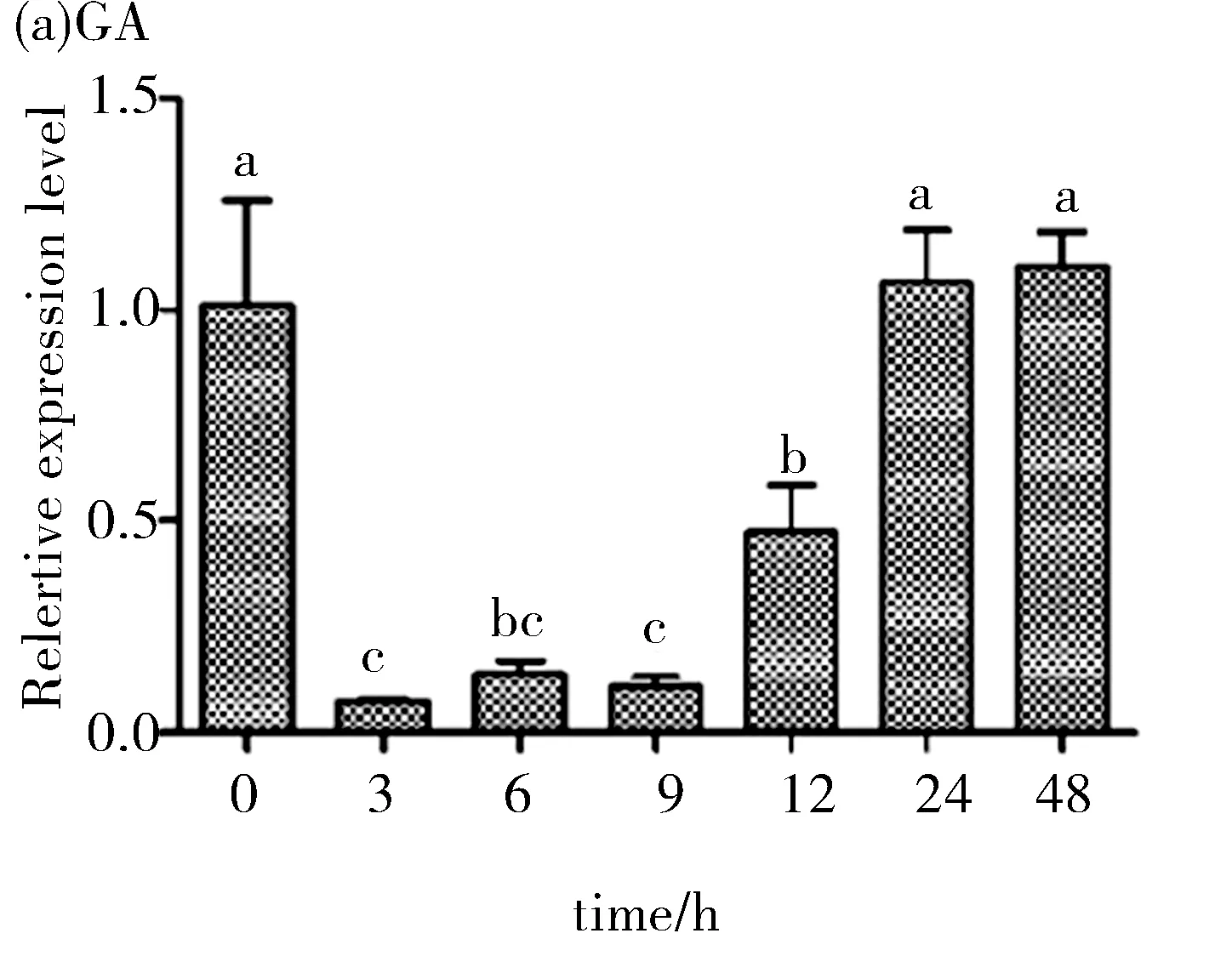
a,b,c,d表示该处理下表达量数据间存在显着差异(P<0.05)
图7 植物激素ABA与GA3处理下甘蓝型油菜BnHY5-1基因的相对表达量
Fig.7 Relative expression ofBnHY5-1 under ABA and GA3treatment
3.6 甘蓝型油菜BnHY5重复基因表达差异分析
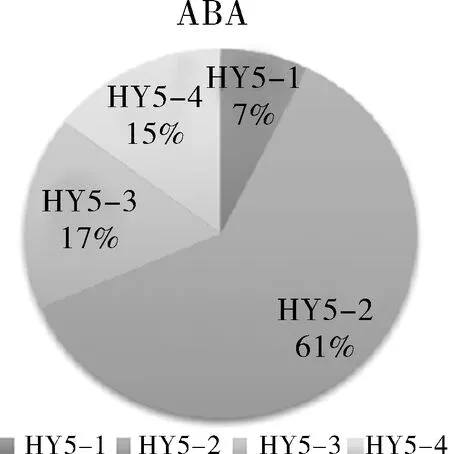
与未做处理时油菜HY5各基因的表达量相比,甘蓝型油菜在高温胁迫(HT),冷胁迫(LT)和ABA喷洒后,BnHY5的表达均上调.在盐胁迫(salt)处理下,BnHY5的表达均下调.各个基因的表达量以同种处理中最低的一个为标准1,甘蓝型油菜BnHY5各基因不同处理下的表达量见图9(a).研究结果表明,在不同的非生物胁迫下,甘蓝型油菜中BnHY5四个基因表达量存在差异,BnHY5-2的表达量是最高的.以四个基因的表达量之和为100%,分别计算单个基因的表达量占总量的比例,见图9(b).以此分析各基因在不同胁迫下表达量占比的差异.据结果可以看出,同类型的胁迫下四个基因的表达模式相同,不同的胁迫下,四个基因的表达模式发生变化.例如在高温和低温胁迫下,四个基因的表达量占比有着相同的特点:HY5-2的表达量占比最高,达到总量的一半,HY5-4和HY5-3的占比为24%和16%.而HY5-1的占比最少.而在盐胁迫中(Salt stress),HY5-1的表达量占比仅此于HY5-2,达到31%.由此可知,重复基因在非生物胁迫下的差异表达与非生物胁迫的类型相关.不同类型的非生物胁迫,植物中参与应答的调控通路不一样,因此在这些通路中与HY5蛋白发生相互作用的因子就不同.这可能是导致BnHY5重复基因在不同非生物胁迫下,有不同的表达量占比的原因.

a,b,c,d代表该处理下BnHY5-1基因表达量存在显著差异(P<0.05)
图8 非生物胁迫和外源激素处理下BnHY5-1基因的相对标准表达量
Fig.8 Relative standard expression ofBnHY5-1 gene under abiotic stress treatment
4 讨 论
4.1 拟南芥HY5蛋白与甘蓝型油菜HY5蛋白的差异分析
根据氨基酸序列比对,找到了拟南芥HY5蛋白与油菜HY5蛋白主要的存在差异的部位.发现氨基酸序列50~60处4个亲水性较强的氨基酸的缺少以及肽链中额外存在4处亲疏水性不同氨基酸的替代.这种差异可能使得该结构域局部构型发生变化.但是运用蛋白结构建模得出的HY5 的LZ整体结构没有发生特别明显的改变.这种差异对这个蛋白的结构与功能有什么影响需要进一步的研究.
4.2 甘蓝型油菜BnHY5与其同源基因BnHYH的异同
甘蓝型油菜HY5与其同源基因HYH在蛋白结构上均有bZIP功能结构域和COP1 interaction区,说明两者在光信号相关的信号通路中有着相似的功能.甘蓝型油菜HYH蛋白亮氨酸拉链(LZ)部分关键的50个(90~140)氨基酸结构建模结果表明,该部分不是仅仅可以形成LZ Homodimer部分,还可以形成与异源二聚化和异源三聚化相关的Nucleoporin Nup58/Nup45 Curated Alternative类似的结构域;和与核孔蛋白形成以及核定位相关的Nuclear Distribution Protein Nude-like 1结构.而对油菜HY5 LZ这50个氨基酸序列建模,发现只存在亮氨酸拉链结构域;说明油菜中HYH可能有着更多未知的功能.甘蓝型油菜BnHY5与其同源基因BnHYH在黑暗下的表达量都低于光照下的表达量,这与拟南芥中相关研究的结论一致[8-9].光照条件下HY5 不能与移动到细胞核外的COP1相互作用,使其大量积累,促进了 HY5 转录因子下游信号的传递[19].黑暗条件下,HY5与在细胞核内积累的COP1 的N末端结合使HY5蛋白泛素化降解,负调控 HY5 下游光形态建成下游基因的表达[20],因此HY5在光照下比黑暗下的表达高.油菜在黑暗下,HYH的表达量趋近于0,说明黑暗条件下油菜中主要发挥作用的是HY5. 油菜BnHY5与其同源基因BnHYH在黑暗下的表达量都低于光照下的表达量,这与拟南芥中相关研究结论一致[9,18,21].二者的表达模式与拟南芥HY5表达模式有着相同的特点.对HY5同源基因HYH功能差异方面的研究需要更加精细的结构研究和功能分析.

(a)



(b)
图9 甘蓝型油菜BnHY5重复基因在不同胁迫下的差异表达
(a) 不同处理下的各个基因的相对表达量;
(b)不同处理下各个重复基因的表达占比a,b,c表示同个处理下基因相对表达量存在显著差异(P<0.05)
Fig.9 Analysis of expression ofBnHY5 duplication with different treatments
4.3 甘蓝型油菜BnHY5与非生物胁迫的关系
甘蓝型油菜BnHY5在非生物胁迫下表达量的变化是确实存在的,说明甘蓝型油菜BnHY5与非生物胁迫相关.盐胁迫下BnHY5的表达量下降,说明该基因在油菜盐拮抗的调控中是一个负调控因子.甘蓝型油菜在外源GA3处理后,BnHY5的表达量下调了;这与其他研究中GA3能够抑制 HY5 的积累[21]的结论相证.其他胁迫处理后BnHY5的表达变化证明了甘蓝型油菜HY5基因响应非生物胁迫.从BnHY5表达分析看来,在油菜中具体的调控过程中,BnHY5表达量的差异可能更强调量的变化带来的作用.此外,同类型的胁迫处理下BnHY5重复基因有着相近的表达占比,而不同的非生物胁迫下,BnHY5多个基因表达量占比存在差异;说明BnHY5重复基因很有可能与非生物胁迫相关.推测在不同非生物胁迫下,它们的应答机制及反应时间和速度都是不同的.但是重复基因表达的具体调控过程还需要更进一步的研究.
研究表明,在拟南芥光信号传递过程中有超过60%的基因是 HY5的靶基因[22-23].HY5转录因子ACGT序列是植物中的各种刺激响应基因启动子的一个核心区[23].HY5转录因子参与植物胁迫下生命活动相关基因的调控,所以分析多种胁迫条件下HY5表达变化趋势是必要的.HY5基因在非生物胁迫上的研究很少.而甘蓝型油菜中该基因还存在多个重复基因.本研究克隆所得四个甘蓝型油菜HY5重复基因存在功能,并且与非生物胁迫相关.通过对甘蓝型油菜非生物胁迫下BnHY5的表达分析,了解到甘蓝型油菜在不同的非生物胁迫下的表达特点,以及重复基因对非生物胁迫的响应.为未来培育抗逆性强的油菜品种提供了分子育种的理论基础.
猜你喜欢
作物学报(2022年12期)2022-10-14
湖北农业科学(2022年11期)2022-07-18
作物学报(2022年4期)2022-02-10
作物学报(2022年4期)2022-02-10
作物学报(2022年3期)2022-01-22
园艺与种苗(2020年12期)2021-01-08
实用肿瘤学杂志(2020年4期)2020-12-08
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
医学综述(2011年12期)2011-12-09
