
茶树白化突变研究进展
2020-04-25 07:15刘丁丁梅菊芬王君雅汤榕津陈亮马春雷
中国茶叶 2020年4期
刘丁丁,梅菊芬,王君雅,汤榕津,陈亮,马春雷*
1.中国农业科学院茶叶研究所国家茶树改良中心 农业农村部茶树生物学与资源利用重点实验室,310008;2.中国农业科学院研究生院,100081;3.江苏省茶叶研究所 江苏省茶树种质资源圃,214063
茶树起源于我国西南地区,作为一种多年生木本经济作物,在世界范围内广泛种植[1-2]。随着茶树栽培和选育技术的发展,育种者们发掘和培育了许多优良的茶树品种,其中白化茶树品种加工而成的茶叶因具有较高的氨基酸含量和优质的外形在市场上广受欢迎。我国自唐朝起就有对白化茶的介绍,历史上记载的描述白化茶树的史料有许多,如唐朝陆羽《茶经》“七之事”中记载“永嘉图经,永嘉县东三百里有白茶山”;北宋宋子安《东溪试茶录》中记载“茶之名有七,一曰白叶茶,民间大重,出于近岁,园焙时有之,地下以山川远近,发不以社之先后。芽发如纸,民间以为茶瑞”。1864年张星焕《皖游纪闻》中记载“兰生言,石埭某山近日间产白茶。其味绝殊,但不可多得,或千百株中偶有一株变白,或今年白,明年仍可复原色,土人以为瑞茶,得则珍藏之”;而产于慧苑岩火焰峰下的白鸡冠是延续至今的适制乌龙茶的白化茶树资源[3]。现代对白化茶树种质资源的研究,要追溯到上世纪七八十年代,研究者最初主要从白叶1号组织结构、生理生化等角度开展白化茶树资源的相关研究;近十几年,随着组学技术的快速发展,有大量利用转录组、代谢组、蛋白质组等技术对这一表型进行研究的文章发表。本文从白化茶树资源的分类、生理生化特征、分子机制等方面对相关研究进展进行了综述,以期为茶树白化机制的解析提供参考。
一、白化茶树突变体分类
白化茶树是一类在特定环境条件下茶树新梢呈现白化表型的茶树突变体。根据诱导白化产生的因素,现有白化资源主要分为生态敏感型、生态不敏感型、生态复合型和未知型几类。生态敏感型白化茶树主要是对气候因子比较敏感,可分为低温敏感型和光照敏感型(表1),其中低温敏感型茶树的白化现象主要是受早春茶芽萌发时的温度影响,并且随着温度的升高,白化表现减弱[4],这种类型的茶树在自然栽培条件下一般只在春末夏初发生“返白—复绿”现象,如白叶1号、千年雪等。而对于光照敏感型白化茶树,多数资源全年表现为白化现象,如黄金芽、御金香等。与生态敏感型白化茶树的“阶段性白化”不同的是,生态不敏感型白化茶树的白化现象属于“恒定性白化”,即表现为白者恒白,绿者恒绿的现象[5],如花月等。生态复合型白化茶树是指茶树组织的一部分依赖温度或者光照的变异,一部分对生态因子不敏感,如金玉缘、春雪3 号等。而对于未知类型的白化茶树资源,笔者认为属于上述3种类型之一,但仍需进一步的试验验证。

表1 白化茶树资源统计
对目前已有报道的白化茶树资源进行综合分析可发现,白化茶树也可依据芽叶颜色进行分类,具体可分为以白色系和黄色系为主的纯色类型和以斑马叶、条纹叶、斑点叶为主的非纯色类型(图1)。其中白色系茶树主要表现为低温敏感型,黄色系茶树主要表现为光照敏感型;在氨基酸含量上,白化茶树资源的氨基酸含量较常规绿色茶树品种高,但也并不是所有的白化茶树资源氨基酸含量都高。表明了这些资源虽同为叶色突变体,但是不同资源之间的叶色形成机制及生理生化特征存在一定的差异,也正是这种差异的存在为我们进行新种质的创制提供了丰富的材料。
二、白化茶树生理生化特征
植物叶片的色泽是叶绿体体内的光合色素和液泡中的花青素综合作用的结果,其中光合色素在光合作用中具有吸收、传递、转化光能的作用,其含量是决定光合效率的重要因素[34]。叶绿体不仅能通过光合作用为植物的生长发育提供养料,同时也是叶绿素、类胡萝卜素和叶黄素等色素物质合成的场所,因此其形态结构与植物叶片色泽的变化具有紧密的联系[35]。所以研究白化茶树突变体叶绿体结构变异以及光合性能变化是解答茶树白化机理的基础。

图1 不同叶色的茶树资源
1.叶绿体超微结构
杜颖颖[36]利用透射电镜观察了处于15℃、20℃、25℃条件下的小雪芽叶片中叶绿体的超微结构,发现在15℃条件下,叶肉细胞内无成熟叶绿体存在,可观察到大量的白色体,但白色体内无类囊体和基粒的堆叠。在20℃和25℃条件下,叶肉细胞内均观察到发育完整的叶绿体结构,内有发育良好的类囊体和紧密堆叠的基粒片层,不同的是在20℃条件下观察到的叶绿体膜有损伤,推测可能是此时叶绿体膜的发育还不完全。李素芳等[37]利用透射电镜观察白叶1号阶段性“白化—返绿”过程中叶绿体的超微结构,发现在返白阶段叶绿体不断退化解体、内部空泡化,在复绿阶段叶绿体结构又重新恢复正常,并提出白叶1 号的白化现象可能是由核基因或核质互作引起的[38]。李娜娜[39]则以白色系资源小雪芽为材料,研究发现其不同叶位叶绿体超微结构的变化与白叶1 号不同白化阶段的结果极其相似,在白色叶片中也出现了叶绿体解体和内部空泡化的现象,而低叶位的偏绿色叶片叶绿体结构又恢复了正常。韦康等[40]以光照敏感型白化品种中黄2 号和正常绿色品种龙井43 为试验材料比较叶绿体的超微结构,结果发现中黄2 号叶绿体结构完整,没有出现解体或大量囊泡,但是基粒片层堆叠少于龙井43。由此可以看出,几乎所有白化茶树叶色突变体的形成,在细胞结构层面都伴随着叶绿体空泡化、类囊体发育不完全、白色体的产生和基粒片层结构缺乏等现象,但它们的相互关系还有待进一步研究。
2.光合作用
光合作用为茶树碳、氮代谢提供碳源和能量,同时也是决定生产力高低的重要指标。但是茶树的光合能力不仅受到环境的影响,也受品种自身遗传因素的制约[41]。到目前为止,对白化茶树品种在生理生化、叶色相关基因表达调控等方面已有一些研究,但是对白化茶树的光合生理特性研究还较少。李勤等[42]从净光合效率、气孔导度、胞间CO2浓度和蒸腾速率等角度分析白叶1号在“返白—复绿”几个阶段的光合作用变化,发现在白化阶段(白化前期、白化中期、白化期)叶片的净光合速率、气孔导度、蒸腾速率呈现先升高后下降的趋势,而胞间CO2浓度则呈现先下降后上升的趋势;在复绿阶段(复绿前期、复绿中期、全绿期)叶片的净光合速率、气孔导度、蒸腾速率表现为逐渐上升的趋势,胞间CO2浓度表现为逐渐下降趋势。胞间CO2浓度在阶段性“返白—复绿”过程中的变化和净光合速率、气孔导度、蒸腾速率的变化呈现相反的趋势,这可能与叶片的发育程度和叶绿体的结构有关。Dong等[43]测定了黄金芽在自然光照和遮阴条件下叶片的净光合速率、气孔导度、胞间CO2浓度、蒸腾速率,发现和遮阴条件下的绿色叶片相比较,自然光照条件下的黄色叶片净光合速率、胞间CO2浓度均降低,而蒸腾速率和气孔导度均升高。由此可看出,白叶1 号和黄金芽虽同为叶色变异资源,但其光合性能还是有较大区别的。
3.生化成分
茶树作为一种叶用作物,叶片生化成分的种类和含量是评价茶叶品质和茶叶功能的重要指标[44]。对不同白化茶树进行生化水平上的分析,有助于了解不同白化茶树资源的特征特性,从而更好地指导相关资源的开发利用。
(1)主要代谢物
氨基酸、多酚类物质和咖啡碱等是茶树中重要的代谢物质,也是茶叶中重要的呈味物质,对茶树生长发育和茶叶品质影响很大。而白化茶树资源普遍表现为高氨基酸含量、适中的茶多酚,但在不同的处理下,其生化物质也会发生明显的变化。如Li 等[45]对黄金芽进行遮阴处理,结果表明,遮阴处理的黄金芽新梢咖啡碱含量高于自然光照下的黄金芽新梢中的含量;随着遮阴程度增加,黄金芽新梢中总氨基酸含量增加;而自然光照下,新梢中茶多酚总量明显低于单层遮阴的黄金芽,但高于双层遮阴的黄金芽。这说明遮阴有利于咖啡碱的积累和氨基酸含量的增加,但是过度遮阴会引起茶多酚含量的降低,这可能是遮阴导致茶树体内碳氮代谢发生变化,进而引起各种次级代谢物质含量发生变化的原因。此外,Lu等[46]以一个“带白化枝条的正常绿色茶树”为材料,比较白化枝条和绿色枝条中次级代谢物质的变化。研究表明,与绿色叶片相比,白化叶片中EGCG、EGC、ECG、EC 含量大幅度降低;咖啡碱含量略微降低;而游离氨基酸,如茶氨酸、谷氨酸、谷氨酰胺、精氨酸等显著升高。杜颖颖[36]以小雪芽、白叶1 号、千年雪、黄金芽和福鼎大白茶为材料,研究白化资源与绿叶品种中茶多酚和咖啡碱含量的差异,结果表明,白化资源中茶多酚和咖啡碱含量显著低于绿叶品种福鼎大白茶。
(2)色素含量
叶绿素和类胡萝卜素作为光合色素,不仅在光合作用中起到捕获、吸收、传递光能的作用,同时作为主要的脂溶性色素,其含量的高低直接决定茶树叶片的颜色,进而影响茶叶干茶和叶底色泽的形成。Cheng等[47]以绿色正常茶树品种英红9号及其白化突变体为材料,在春季和冬季分别测定了叶绿素a和叶绿素b的含量,发现两个季节的白化突变体中叶绿素a和叶绿素b的含量均显著低于正常茶树品种中的含量。Li 等[45]以黄金芽为研究材料,对其进行遮阴处理,研究遮阴处理前后黄金芽的各种色素含量的差异。结果表明,遮阴处理绿色叶片的叶绿素a、叶绿素b和叶绿素总量均明显高于光照下黄色叶片的含量;而遮阴处理绿色叶片的紫黄质、新黄质、叶黄素和类胡萝卜素总量均低于光照下黄色叶片的含量,分析认为光照强度增强可能刺激了类胡萝卜素的积累。Wang 等[48]分别测定中黄2 号和龙井43 中的叶绿素、类胡萝卜素和叶黄素的含量,发现中黄2 号中叶绿素、类胡萝卜素和叶黄素的含量均显著低于龙井43。
(3)酶活性
研究发现,白化茶树新梢在白化时期体内酶活性会发生显著的变化,主要表现为1,5-二磷酸核酮糖羧化酶活性下降,蛋白水解酶活性增强;在复绿时期两种酶活性变化与白化时期相反。李素芳[49]研究白叶1号各白化时期和复绿时期酶活性与氨基酸含量的关系时发现:随白化程度加深,蛋白水解酶活性增强,将体内多余蛋白质水解,导致游离氨基酸含量增加,推测这可能是白叶1 号在白化时期氨基酸含量较高的原因。
过氧化物酶(POD)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)作为茶树体内的保护酶,在茶树白化期间其活性也会发生变化。陆建良等[50]研究发现,在白叶1号返白过程中,POD酶活力升高,SOD 和CAT 酶活力降低,而当芽梢复绿后,POD 酶活力降低,CAT、SOD 酶活力回升;Wu等[51]在研究光照和遮阴处理条件下白鸡冠叶片中POD、SOD、CAT 酶活性变化时,也得出相同的结论。推测造成此类现象的原因可能是叶片内叶绿体结构发育异常,打破了活性氧平衡,使茶树启动了保护酶机制。
三、组学技术在白化茶树资源研究中的应用
近些年来,随着各种组学技术的快速发展,将茶学的相关研究带入了一个新的高度。截至目前,已有2 个茶树基因组草图相继发表[52-53],标志着茶学研究正式进入了后基因组时代,这为不同白化茶树分子机制的解析提供了新的研究方向。
1.基因差异表达
基因差异表达作为功能基因组学的重要组成部分,是从RNA水平上研究生物体在不同组织结构、不同发育时期、不同环境条件下基因的表达情况。在白化资源的研究中,相关的分析方法已得到广泛应用。通过将不同叶色新梢差异表达的基因进行Blast、GO、KEGG等分析,可初步了解引起白化表型的基因有哪些,以及它们主要富集在哪些代谢通路中,从而为探究白化茶树资源的白化分子机制提供支撑。
因白叶1 号是较早发现的叶色变异资源,所以针对它的相关研究也最为系统,不同阶段的差异表达技术在这一资源上均有应用,如王新超等[54]利 用DDRT-PCR (Differential display of reverse transcriptional PCR)技术从白叶1 号不同叶色新梢中获得58 条差异表达片段,进一步通过RT-PCR分析,验证了12条阳性片段;Blastx序列比对发现,5个阳性片段分别与编码血红素结合蛋白、甲硫氨酸合酶、1-氨基环丙烷羧酸合酶等的基因序列高度相似。李娟[55]利用SSH(Suppression subtractive hybridization)技术从白叶1号绿色叶和白化期叶片中分离80个差异表达基因,进一步分析筛选到1个可能调控白叶1号返白现象的钙调热激蛋白基因和2 个与高氨基酸性状相关的丝氨酸-乙醛酸转氨酶、丝氨酸羟甲基转移酶基因。吴 扬[56]、Ruan 等[57]分 别 利 用cDNA-AFLP (cDNA-amplified fragment length polymorphism)技术从白叶1 号不同叶色新梢中获得了31 个和127 条差异表达片段,对这些差异表达的基因进行同源序列比对发现,均与细胞周期、信号传导、物质和能量代谢等相关。Ma等[58]利用基因芯片技术从白叶1号不同叶色发育时期的叶片中获得671个差异表达基因,这些基因主要参与到能量代谢、碳固定、次级代谢、植物生长和防御等过程,并鉴定了一些参与叶绿素合成和叶绿体发育的催化酶和调控蛋白,其中发现编码镁-螯合酶基因的表达趋势和叶色性状的变化相一致。Li等[59]对白叶1号黄绿期、白化期、返绿期3 个时期的新梢进行RNA-Seq 测序,通过两两比较分析差异表达基因,这些差异表达基因主要参与到代谢途径、次级代谢物生物合成、苯丙烷代谢和碳固定途径中。
近些年,随着叶色变异资源的增多,基因差异显示技术也开始在更多的品种资源中得到应用(表2),如李娜娜等[60]利用RNA-Seq法对福鼎大白茶和小雪芽叶片的差异基因进行了研究,筛选到1 821 个上调表达基因和1 779 个下调表达基因,进一步对差异基因进行GO 和COG 分析,认为引起小雪芽新梢白化的遗传机制可能与蛋白质翻译后修饰过程有关。Wu等[51]利用RNA-Seq技术对白鸡冠的白化叶片和绿色叶片的差异表达基因进行研究,这些差异表达基因主要参与叶绿体发育、活性氧清除、光合色素合成、次级代谢和昼夜节律调控过程。一些科研人员也分别利用RNA-Seq技术研究了御金香、华白1 号、黄金芽、炎陵银边茶等不同叶色时期的差异表达基因,结果发现差异表达基因同样参与了叶绿体发育、光合色素合成、次级代谢等过程[61-66]。这说明虽然不同白化资源的遗传背景并不相同,但在基因表达层面上,叶绿体发育异常和叶绿素合成受阻是导致这类资源白化表型产生的直接原因。

表2 白化茶树资源的基因差异表达研究
2.蛋白质组学
蛋白质组学是以生物体内基因所表达的全部蛋白质为研究对象,从整体、动态的角度研究蛋白质特征,从而帮助科研人员从蛋白质水平上整体而全面地理解生物体的组织变化、物质代谢等过程[67-68]。目前,基于双向凝胶电泳、质谱的蛋白质组学技术在白化茶树资源的研究中已得到初步应用(表3)。如Li 等[69]在白叶1 号3 个叶色发育阶段利用基质辅助激光解吸电离飞行时间质谱(Matrix-assisted laser desorption/ionization time of flight mass spectrometry,MALDI-TOF-MS)技术鉴定了26个生育期差异表达蛋白,这些差异表达蛋白主要参与碳氮硫代谢、光合作用、RNA 和蛋白质加工、胁迫防御等过程;之后同样在白叶1 号3个叶色发育阶段利用质谱鉴定了22个叶绿体差异表达蛋白,发现与光合作用、物质代谢、核酸代谢、应激响应等相关[70]。Xu 等[71-72]从转录后修饰角度,利用LC-MS/MS (Liquid chromatography-tandem mass spectrometry)技术先后从白叶1号不同白化时期的表达蛋白中鉴定了3 161 个乙酰化位点和3 530 个琥珀酰化位点,其中与光合作用、碳固定、氨基酸生物合成和叶绿素代谢相关蛋白的琥珀酰化水平在各个白化时期存在差异,同时与光合作用和次级代谢相关蛋白的乙酰化水平也发生了变化,而且部分琥珀酰化和乙酰化蛋白的作用存在部分重叠。除了对低温敏感型白叶1 号进行研究之外,还有研究分别利用同位素相对标记和绝对定量技术(Isobaric tags for relative and absolute quantitation, iTRAQ)分别鉴定了光照敏感型的黄金芽和中黄1 号的173 个、56 个、437 个差异表达蛋白,大多数蛋白均参与光合作用、类黄酮代谢、碳水化合物的转运与代谢、次级代谢物的合成等过程[43,73-74]。
3.代谢组学
代谢组学可以通过定性或定量分析生物体在不同发育时期、不同组织结构、不同环境条件下代谢产物的变化情况,从而整体揭示生物体的代谢调控机制和生命活动规律。近些年来,研究者们利用代谢组学技术与多元统计分析相结合,发现不同叶色茶树品种之间、同一茶树品种不同叶色之间的代谢物变化均存在显著的差异(表4)。如Zeng 等[75]利用UPLC-QTOF-MS(Ultra-high performance liquid chromatography-quadrupole time-of-Flight mass spectrometry)技术在白叶1 号3 个叶色发育阶段共鉴定了92 种差异代谢物,这些差异代谢物主要参与类黄酮、苯丙烷、氨基酸等的生物合成途径,并发现在白化期内氨基酸含量较高、碳水化合物含量较低,儿茶素和黄酮类物质含量在白化前期和返绿期含量较高;Li等[76]利用LC-MS技术同样在白叶1号的3个叶色发育阶段鉴定了65个差异代谢物,这些差异代谢物与半乳糖、色氨酸代谢和苯丙烷、类黄酮生物合成等相关。Zhang等[77]在黄金芽白化叶片和绿色叶片中共鉴定了52个差异代谢物,与类黄酮物质生物合成和苯丙烷代谢相关,并发现CG、槲皮素和黄酮醇苷类物质在白化叶片中大量积累,而杨梅苷和EGCG含量则较低;之后Zhang等[78]又鉴定了41 个差异代谢物,与氨基酸和碳骨架代谢相关,其中在白化叶片中谷氨酸、谷氨酰胺、茶氨酸、柠檬酸和α-酮戊二酸含量增加,而单糖、二糖和咖啡碱含量降低。Shen 等[79]为解析白化叶片氨基酸代谢增强的机制,利用HPLC (High performance liquid chromatography)检测了黄山白茶白化前期、芽期、白化期和返绿期新梢中的游离氨基酸含量,并以绿色品种黄山种为对照,同时利用qRT-PCR 技术检测了参与氨基酸代谢的基因在几个不同时期的表达量,结果表明茶氨酸合成增强和氨基酸转运能力降低是导致白化时期新梢中氨基酸含量增加的主要原因。此外,Feng 等[80]利用HPLC 技术研究了不同叶色品种之间的代谢谱差异,发现和绿色品种福鼎大白茶相比,白化品种白叶1 号、黄金芽、御金香叶片中含有较低的叶绿素、类胡萝卜素、咖啡碱、儿茶素,而玉米黄素和茶氨酸含量较高。
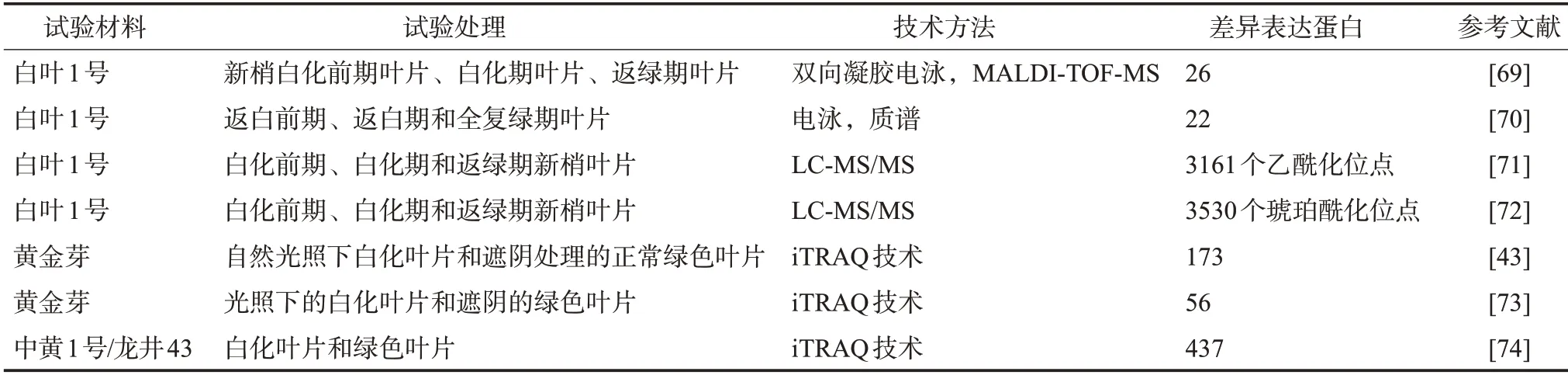
表3 白化茶树资源的蛋白质组学研究
目前,利用代谢组学技术研究白化茶树资源的代谢物变化,主要关注氨基酸、黄酮类和糖类等物质,而对香气物质的报道很少。根据已有研究,Dong 等[81]利用GC-MS 技术分析了4月、8月和12月份英红9 号白化叶片和绿色叶片中香气物质的变化,发现3 个时期中白化叶片的顺-3-乙烯醇和芳樟醇含量均显著低于绿色叶片,其他香气物质含量变化不明显;这可能就是导致白化叶片中香气物质含量低于绿色叶片的直接原因。

表4 白化茶树资源的代谢组学研究
四、分子标记技术在白化资源研究中的应用
随着测序技术的快速发展,也推动了DNA分子标记技术的广泛应用,与形态学标记、同工酶标记相比,DNA分子标记直接从基因组水平上研究生物的遗传变异,不受发育时期、环境因素的限制[82]。目前分子标记技术在白化茶树资源研究中主要应用于遗传多样性分析、品种鉴定、遗传图谱构建及QTL定位等方面。
王松琳[83]等利用SSR (Simple sequence repeats)分子标记对16个白化茶树品种进行遗传多样性分析,通过分析等位基因数、多态性信息含量和shannon 值等,发现品种间遗传结构差异明显。并筛选出3 对核心SSR 引物,用于品种鉴定。王开荣等[3]利用RAPD(Random amplification polymorphic DNA)技术结合DNA序列测定,以福鼎大白茶为对照,研究黄金芽、千年雪、御金香、白叶1 号、四明雪芽、金光和花月等7 个白化品种的特异DNA 分子标记,以期找到与白化相关的基因。通过对单一品种唯一扩增位点和缺失位点分析,发现一些位点可能与叶绿素代谢和抑制与叶绿素代谢相关基因的表达有关;进一步筛选了21个特异分子标记,并成功测序了4条DNA序列,通过Blast序列比对发现与蛋白激酶、转运蛋白家族基因等具有同源性;同时这一分子标记也可应用于白化茶树品种的遗传鉴定。吴永胜[84]以白叶1号、黄金芽、御金香、四明雪芽等10份茶树资源为材料,先利用50对RAPD反应体系及程序进行PCR 扩增,引物S20 在御金香中扩增出1条特异性片段,之后在引物S20基础上设计SCAR引物SY20,用于扩增10份材料,发现该引物同样在御金香中扩增出特异性片段,表明了RAPD标记成功地转化为SCAR标记,并且利用该标记能够将御金香从多种茶树资源中鉴别出来。
叶绿素、类胡萝卜素等色素物质直接影响茶树新梢叶片色泽,属于数量性状。为研究其遗传机制,王松琳[35]以白鸡冠和龙井43 作为父母本构建遗传群体,对亲本和子代个体测序获得大量SNP(Single nucleotide polymorphisms)标记,结合拟测交策略构建遗传图谱,随后利用遗传图谱对叶绿素和类胡萝卜素这2 个性状进行QTL(Quantitative trait locus)作图,结果共定位了10个QTL,分布于4 个连锁群上,表型贡献率在18.2%~72.0%,均为主效QTL。
五、展望
综上所述,茶树白化现象的产生与细胞色素的合成和降解、光合系统、叶绿体的发育等密切相关。在生理水平上表现为叶绿体解体或退化消失,进而影响了叶绿素和类胡萝卜素等色素的生物合成;在生化成分上体现为含有较高含量的游离氨基酸;但是在分子水平上茶树的白化机理仍不明确。目前,虽也有部分参与叶绿素、类胡萝卜素、氨基酸合成相关的茶树基因得到了分离克隆,如邓婷婷等[85-86]根据cDNA-AFLP 的结果,利用RACE(Rapid amplification of cDNA ends)技术克隆了泛素活化酶基因和八氢番茄红素脱氢酶3 基因;马春雷等[87]根据芯片杂交结果,克隆了3 个参与叶绿素合成途径的基因:谷氨酸-tRNA 还原酶基因、叶绿素合酶基因、叶绿素醋酸氧化酶基因。但相关研究均属于同源克隆,因此到底是哪个基因的突变导致了白叶1 号、黄金芽等白化突变体的白化表型问题仍未有答案。
从已报道的研究结果来看,在茶树白化突变体研究中还存在诸多亟待解决的问题。
首先,虽然近些年各地陆续发现了很多的白化茶树突变材料,但大多数资源的谱系关系和遗传背景均不清楚;以这些突变体为研究材料的相关研究也多集中在生理生化、基因表达和代谢层面,而缺少遗传层面的深入分析。
第二,因茶树是多年生木本植物,生长周期长,导致构建白化相关遗传群体的难度很大;而茶树基因组庞大,遗传位点高度杂合,使得茶树白化性状的遗传解析困难重重。
第三,因茶树转基因技术尚不成熟,很难通过本源表达验证基因功能,从而导致白化相关基因的鉴定很难开展。针对这些问题,课题组从2013年开始,对保存在国家种质杭州茶树圃的十几份白化茶树资源的遗传特性进行了连续多年的观察,发现这些资源中的大多数都具有叶色遗传规律,如黄金芽、白鸡冠、安吉黄茶、中黄1 号等(表5)。

表5 不同白化品种资源的遗传特性分析
随后课题组以其中3 个具有叶色遗传特性的白化资源为父本,通过多年的连续杂交,已建立了3 个白化性状在子代中能有效分离的F1 群体,这些遗传群体将是以后解答白化茶树资源叶色突变机制的重要研究材料。
猜你喜欢
语文世界(小学版)(2021年5期)2021-06-04
野生动物学报(2021年1期)2021-01-13
热带农业科学(2020年7期)2020-08-31
煤矿安全(2020年4期)2020-04-24
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
山东农业科学(2018年3期)2018-03-29
山东农业科学(2018年11期)2018-01-17
江苏农业科学(2016年2期)2016-04-11
江苏农业科学(2014年3期)2014-07-16
