
化石
2020-12-05 07:14王浩
生物进化 2020年3期
昆虫多样性结构色的形成机制

自然界中的颜色主要有三个方面的来源:即生物发光、色素色和结构色。色素色也叫化学色,结构色也叫物理色。结构色是自然界中色彩最为纯净且最强烈的颜色,通常由生物纳米光学结构与自然光相互作用而产生。由于结构色很容易在长期的化石埋藏作用中丢失,地质历史中原始的结构色的证据极其罕见。近期,研究者通过对白垩纪缅甸琥珀中大量的具金属色彩的昆虫进行了系统研究,发现纯净而强烈的颜色可直接在多种昆虫身体表面保存下来。通过琥珀超薄切片、电子显微镜和透射电子显微镜等技术的分析发现,一种青蜂科昆虫胸部表面的蓝绿色是由多层重复出现的纳米级构造组成,代表了一种典型且常见的结构色类型,即多层反射膜。通过进一步的光学理论模型分析得到,其反射波长与观察到的昆虫颜色波长接近的结论,证实白垩纪琥珀昆虫身体表面显示的颜色可能是原始颜色。这一发现直接证明了超微纳米级的光学元件在长期地质历史中可得到稳定保存,从而否定了前人关于昆虫金属色不能在中生代化石中保存的观点,对认识早期昆虫结构色生态功能的演化具有重要意义。(Proc. R. Soc. B,2020,20200301)
中国以外首次发现中华震旦角石
中华震旦角石(Sinoceras chinense)是一类已经灭绝的头足类软体动物,繁盛于晚奥陶世;自20 世纪20年代以来,一直被确定为晚奥陶世凯迪早期的标准化石。很长一段时间里,中华震旦角石仅在中国境内,包括华南、塔里木、西藏、滇西等地的相应层位有化石记录,在中国以外未见任何报道。近期,研究者在泰国西部地区进行野外考察时,首次在泰国上奥陶统地层中发现了中华震旦角石。这批化石产于泰国西部北碧府Si Sawat 县的Nautiloid Site 地质保护区内。长期以来,这些化石都被错误地鉴定为珠角石类(actinocerids),化石产出层位Tha Manao 组的时代确定也存在问题。在这次野外考察中,研究者首次将产于Tha Manao 组上部的头足类化石确认为中华震旦角石,且其时代应为晚奥陶世凯迪早期,与我国华南扬子台地的宝塔组、泰国南部的Pa Kae 组大致同期。泰国西部地区与我国滇西保山地区相同,在早古生代均属于滇缅马地体,位于冈瓦纳周缘的低纬度地区。本次确认的中华震旦角石是在泰国的首次记录,也是在中国以外地区的第一次报道。这一发现不仅对在古生代冈瓦纳东北缘的古板块重建提供了重要的化石证据,并且进一步证实了中奥陶世到晚奥陶世存在的滇缅马地体漂移、生物地理区系迁移等地质事件。(Palaeowolrd,2020,https://doi.org/10.1016/j.palwor.2020.06.004)
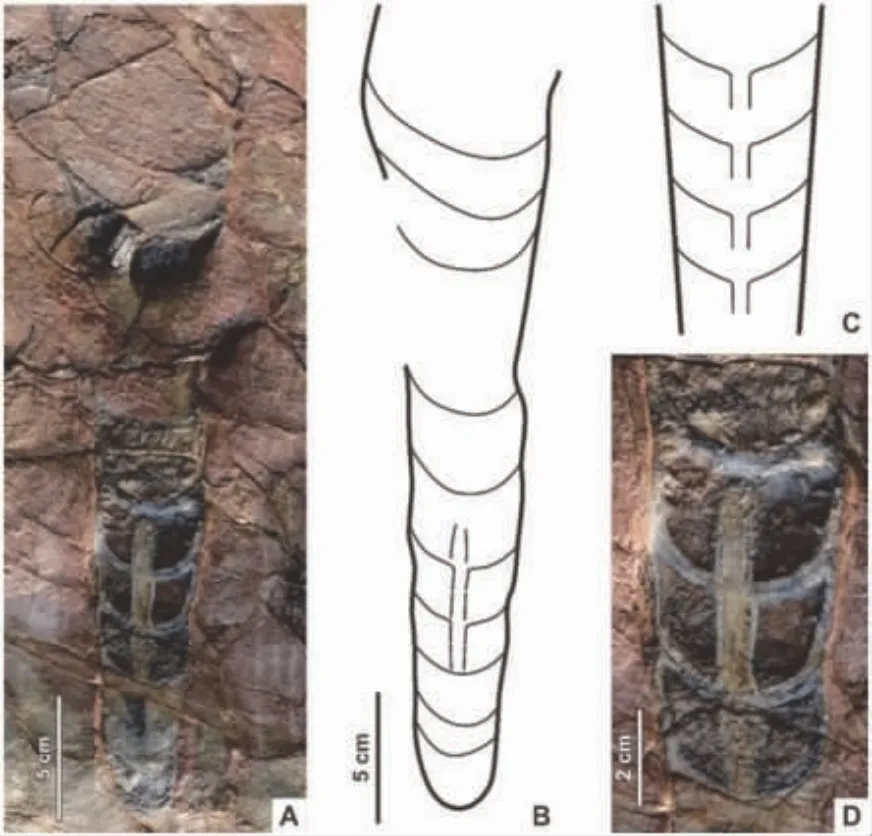
我国发现第二枚古生代蝎子化石
蝎目属于节肢动物门螯肢亚门蛛形纲,是蛛形纲动物中体型较大、较为古老的一类。蝎子的触肢(第二对附肢)末端为钳状,前腹部有7个体节,后腹部为5 个体节加尾节,尾节由球状毒囊和针状毒刺组成,内部具有毒腺;蝎子腹部腹面有一对梳状的栉板,是其独有的感觉器官。化石蝎可分属于48 科近150种,中国古生代蝎子化石极其稀少,先前仅有一例报道。近期,研究者报道了产自乌达煤矿的一枚较为完整且精致的蝎子化石,将其归入始蝎属、未定种(Eoscorpiussp.),是目前中国报道的第二枚古生代蝎子化石,也是世界范围内报道的第三枚产自二叠纪地层的蝎子化石。该标本保存较为完整,头胸甲前中部有一隆起的水滴形中眼丘,中眼之间的一倒“V”型脊将其分开;中眼丘为一“Y”型沟环绕,将头胸甲背面分为较为凸起的中部头顶区域和凸起不明显的两侧胸部区域。螯肢大,突出于头胸部前方;触肢长而相对粗壮,掌宽且短,指长;第二对步足保存有胫节距。后腹部体节的长和宽均依次减少。尾节毒囊呈灯泡状,蛰针粗壮,长且弯曲。其身体表面有许多丘状小颗粒装饰,表皮上较为精细的结构等都清晰可见。由于该标本保存于凝灰岩层中部,表明其生活于树木的高层,但其底层环境为水体,这表明在早二叠世早期,此类蝎子可能就已经适应了陆生生活。此外,作为生态系统中的食肉捕食者,该蝎子化石的发现可证明当时的沼泽森林中已有较为丰富的动物量。该研究较为全面地揭示了早二叠世的“植物庞贝”生态系统群落面貌。(Proceedings of the Geologists’Association,2020,131:160-167)

华北寒武纪奇虾类研究新进展

奇虾类是寒武纪最引人注目的动物类群之一,这类原始节肢动物体型巨大,身体构型独特,具有良好的视力和高超的捕食能力。作为显生宙海洋生态系统中最早出现的顶级捕食者,一直备受人们的关注。传统意义上的奇虾类以奇虾属为代表,而赫德虾类则是近年来人们识别出的另一大奇虾类群。由3 个大板片组成的头壳复合体是赫德虾类的典型标志,此外,具有刀片状内叶的前附肢和四辐对称的口器也是这一类群的重要识别特征。自2009年布尔吉斯页岩中的赫德虾类被识别为奇虾类群的一份子以来,许多新的赫德虾类化石被陆续发现,也有许多曾被归为双瓣壳节肢动物的化石标本被重新解释为赫德虾类的头部板片。到目前为止,已发现的赫德虾类包含9 个属,均产出于全球寒武纪早期至泥盆纪早期的18 个特异埋藏化石群。因此,赫德虾类在寒武纪生态系统中的重要地位逐渐得以显露,且这类在沉积物或水体中利用特化的前附肢兜取食物甚至滤食的奇虾,也大大拓展了我们对奇虾分类和生态多样性的认识。寒武纪中期是赫德虾类形态和生态演化的关键时期,因此了解这一时期不同地理区域之间赫德虾类的多样性非常重要。但由于赫德虾类非矿化的身体仅能在特异埋藏化石库中保存,而寒武纪中期全球特异埋藏化石群主要集中分布在劳伦大陆(今北美大陆的主体),使得我们对于这一时期其他地理区域的赫德虾类了解甚少。近期,研究者报道了在华北地台发现的两属两种寒武纪中期赫德虾类,分别是线纹心虾(Cordaticaris striatus)和镰状寒武耙虾相似种(Cambrorastercf.falcatus),其中线纹心虾是一全新物种。这些进展代表了赫德虾类在华北地台的首次发现,也揭示了赫德虾类在寒武纪中期具有丰富的形态多样性和广泛的地理分布。(Palaeogeography Palaeoclimatology, Palaeoecology 2020,558:109947;Journal of Paleontology 2020,https://doi.org/10.1017/jpa.2020.21)
安徽繁昌发现上猿超科新属种
在系统发育上,包括现生长臂猿科、猩猩、大猩猩和黑猩猩在内的真正的猿类属于冠狭鼻猴类的一个分支,而上猿超科则属于已经绝灭且没有现生代表的干狭鼻猴类的一个分支。简而言之,上猿超科是在人猿超科和猴超科发生分化之前的一个分支,并且中间还隔有萨阿丹猿超科、树猿超科、原康修尔猿超科等分支,其系统发育关系与真正的猿类较远,因此与猿类并不存在直接的祖裔关系。近期,研究者发表了一种发现于安徽繁昌县的上猿超科新属种:金氏繁昌上猿(Fanchangiajini)。属名意为化石产地所在的繁昌县,种名献给古脊椎动物与古人类研究所的金昌柱研究员,以纪念他发现了繁昌上猿的化石以及对安徽繁昌人字洞古人类活动遗址的研究工作所作的贡献。金氏繁昌上猿的化石材料包括108 颗游离牙齿。除下侧门齿外,涵盖了上下齿列中所有齿种。伴生的啮齿类化石表明,含金氏繁昌上猿的裂隙堆积的年代可以与山东临朐山旺组(约1800-1700万年前)对比,而比江苏泗洪的下草湾组(约1900-1800万年前)稍显年轻,属早中新世晚期。
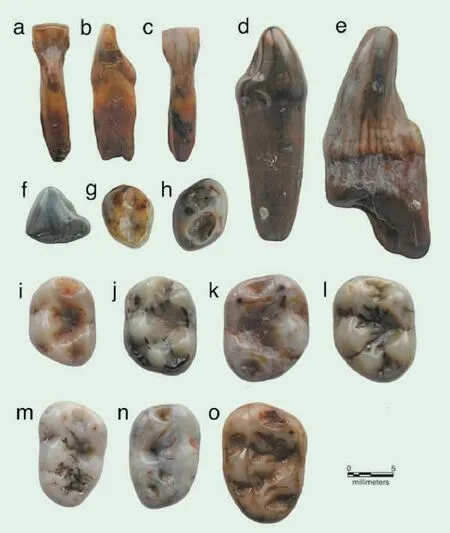
狭鼻猴类于古近纪起源于非洲-阿拉伯陆块。在该陆块于早中新世(约1700-2000万年前)与欧亚大陆发生碰撞形成连接之前,狭鼻猴类已经演化出了原上猿超科、上猿超科、原康修尔猿超科、树猿超科以及猴超科和人猿超科等主要类群。基于目前的化石证据,上猿超科是最早走出非洲进入欧亚的狭鼻猴类类群。上猿超科的化石记录仅分布于欧亚大陆的中新世地层。最早的上猿超科成员是产自中国东部江苏泗洪下草湾的醉猿科,年代为约1900-1800万年前。虽然这一时期欧洲的化石记录非常丰富,但并没有上猿超科化石出现。这就意味着,最初的上猿超科基干成员是通过阿拉伯半岛并沿着环印度洋途径,从非洲进入亚洲南部和东部的热带和亚热带地区的。先前的观点认为,从最初走出非洲的包括醉猿科在内的这些上猿超科成员中演化出了上猿科,并在约1600-1700万年前的早中新世末期向西扩散到了欧洲,更为特化的克鲁泽尔猿科则于中中新世在欧洲起源于上猿科的某个成员。而作为克鲁泽尔猿科干群成员的繁昌上猿的时代早于该科和上猿科在欧洲的出现。在中国的发现表明,上猿科和克鲁泽尔猿科在中国发生分化,并于早中新世晚期或中中新世才扩散到欧洲。繁昌上猿所保留的原始特征说明克鲁泽尔猿科演化自一个来自上猿科或者醉猿科的祖先。欧洲的克鲁泽尔猿科成员都比繁昌上猿属的衍生程度高,并且在繁昌上猿在中国出现至少300万年之后才开始出现。另外,尽管内蒙大庙的上猿超科标本只有一颗上臼齿,但可能代表了一种原始的克鲁泽尔猿科成员。如果这些结论属实,则说明克鲁泽尔猿科在亚洲于中中新世期间持续多样化,然后才扩散到欧洲。繁昌上猿的发现为上猿超科的演化历史提供了新的认识,证明东亚地区是上猿超科重要的早期演化中心。(JournalofHumanEvolution,2020,145:102838)
小巧漏卧鱼——最古老的清道夫型新鳍鱼类

三叠纪是新鳍鱼类辐射演化的重要时期,出现了适应不同生态类型的新鳍鱼类:有牙齿粗壮而锋利的顶端捕食者,有半球形齿的食甲型鱼类,也有牙齿细小或无齿的食浮游生物鱼类。研究者在滇东(曲靖市罗平县和红河州泸西县)发现了一种2.44 亿年前(中三叠世安尼期)的清道夫型新鳍鱼类化石,命名为小巧漏卧鱼(Louwoichthys pusillus)。以小巧漏卧鱼和其他相关的新鳍鱼类为研究对象,在过去10年的分支系统学研究基础上,研究者建立了辐鳍鱼亚纲的一个新目:漏卧鱼目。小巧漏卧鱼是漏卧鱼目最古老的属种,代表了已知最早的清道夫型基干新鳍鱼类。小巧漏卧鱼和这些的捕食者都不一样,它是一种食腐性清道夫型鱼类。它的头骨形态十分特别,颅顶骨骼几乎愈合成一整块,前鳃盖骨下部前倾,上颌较短、后端膨大,下颌纤细,口缘前半部分具有异乎寻常的长而尖的牙齿。它的标准体长只有三个厘米左右,很不起眼,但却是海洋生态系统中的重要一环,因为它可以比微生物更快速地分解其他动物(如大型海生爬行动物)的遗骸,是海底的清洁工,对整个海洋生态系统的正常运行非常重要。漏卧鱼属名源于古代罗平及周边地区的漏卧侯国。得益于这一地区丰富的三叠纪鱼化石材料,研究者在新鳍鱼类的演化关系和分类研究上取得突破性进展。在世界上首次重建了较完整的早期新鳍鱼类进化树,提出了新的基干新鳍鱼类分类体系,将小巧漏卧鱼、贝氏罗平鱼和欧洲的贝洛特梳齿鱼归入新建立的漏卧鱼科。该科的近亲是此前发现于南美及欧洲中、晚三叠世的伪比肯鱼科鱼类,两者组成漏卧鱼目。该研究对于了解三叠纪海洋生态系统食物网的营养结构以及早期新鳍鱼类的演化、分类和生物地理具有重要意义。(Zoological Journal of the Linnean Society,2020,zlaa053)
山西新现二齿兽类白氏桃河兽
中国二叠纪四足类的研究由来已久,在新疆发现过大量二齿兽类化石,而很长一段时间,在华北广泛出露的二叠系地层中未发现二齿兽类,只有几件锯齿龙类化石。1989年报道在内蒙古大青山脑包沟组发现了边缘大青山兽,打破了这一沉寂,但是在出露更为广阔的孙家沟组却依然没有任何发现。2013年,山西阳泉的化石爱好者白志君在孙家沟组发现了一些四足动物化石,后来又陆续有了更多的发现。近期,研究者对已修复的几件头骨化石进行了研究,发表了对其中一件来自于孙家沟组底部保存的近完整头骨及部分下颌的研究成果,根据化石产地及发现者将其命名为白氏桃河兽(Taoheodon baizhijuni),这一新类群有下列特征区别于其他属种:犁骨腹面光滑,缺乏翼骨龙骨突,翼骨中板窄,泪骨与隔颌骨接触,翼骨不与上颌骨接触,枕髁宽、弧形,侧齿骨架细、但有明显的背腹面。系统发育分析研究表明,白氏桃河兽与在老挝发现的两种二齿兽类——背目古尼龙兽及强壮雷佩林龙组成一个单系类群,被包含在一个核心的二齿兽支系中。此前研究认为老挝的两种二齿兽是最基干的肯氏兽形类,其生活时代为早三叠世,而本次研究结果则支持老挝的二齿兽类生活时代可能为二叠纪晚期,并且是由华南板块东缘迁徙而去。(Journal of Vertebrate Paleontology,2020,e1762088)

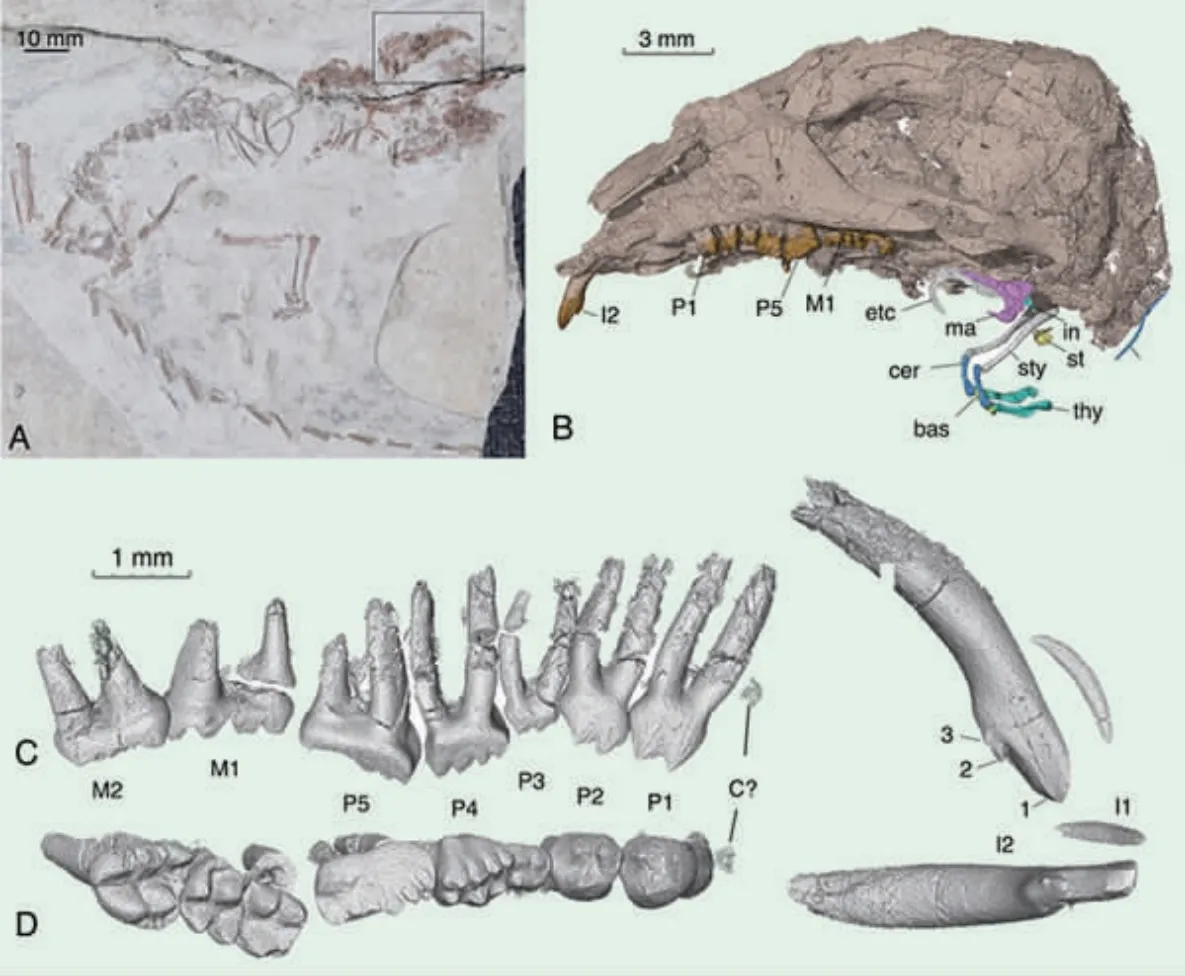
中耳演化发育在热河哺乳动物中的祖先表型
哺乳动物中耳的演化,是从达尔文时代就在广泛探讨的一个问题,也是脊椎动物渐进演化的经典案例。爬行类具原始颌关节(由下颌关节骨与头骨上的方骨构成),中耳只有一块听骨(镫骨=耳柱骨)。与此相比,哺乳动物中耳则由锤骨、砧骨、镫骨形成听骨链,外鼓骨支撑鼓膜收集声音,下颌由一块齿骨构成,形成齿骨-鳞骨颌关节。解剖学、发育生物学、遗传学的大量研究证明,哺乳动物的听骨和爬行动物的齿骨后骨和方骨具有同源关系。古生物学也早已提出中耳渐进演化假说,即从爬行类向哺乳动物的典型哺乳动物中耳的演化过程中,齿骨后骨逐步缩小,并与齿骨分离,最终移入颅基部,成为专司听觉功能的结构。但纤细的听骨很难保存为化石,近两百年来发现的中生代哺乳动物中耳化石极少,其形态细节和关节方式一直不清楚,对其演化过程和机制的解释存在极大的人为性,各种推论、假说存在较大争议。产自辽宁朝阳大平房下白垩统九佛堂组的多瘤齿
兽——潘氏中华俊兽 (Sinobaatar pani)保存了完好的听骨,高精度CT 扫描重建首次揭示了多瘤齿兽5 块听骨确切的三维形态和接触关系。此外,研究者对辽尖齿兽、源掠兽,以及现生哺乳动物中形态较为原始的单孔类针鼹、有袋类负鼠和真兽类刺猬的中耳进行了高精度CT扫描和重建,以期进行系统的对比研究。研究结果表明,辽尖齿兽、源掠兽、中华俊兽这三种中生代哺乳动物体现了中耳演化三个不同的渐进阶段,作为祖先表型不同形态,填补了似哺乳的下颌中耳与典型哺乳动物中耳之间的形态空缺。这些祖先表型体现的演化程度差别,与现生哺乳动物中耳个体发育过程可以对应。三种热河生物群中生代化石中,听骨前端和腹部变化较大,而锤骨和砧骨关节相对保守,表现出听骨前、后端的异时演化发育现象。可能的解释是,在演化过程中,砧骨虽然也在不断缩小,但它一直位于颅基部,与岩骨等颅基结构的位置关系变动不大。而听骨与齿骨的分离或者说听觉功能和咀嚼功能的分离,可以看成是听骨(齿骨后骨)不断缩小、向后退缩,离开齿骨进入颅基部的演化过程,因而变化较大。砧骨-锤骨关节相对于头骨,位置较为稳定,并一直起着一定的辅助颌关节的作用,等同于听骨与颅骨连接的锚点,所以演变速率较低。(National Science Review,2020,nwaa188)
揭秘5 亿年前的小“口板”
节肢动物是显生宙以来地球上物种多样性最高的动物门类。在漫长的地球历史时期,大多数曾经在地球上生活过的节肢动物物种都灭绝了,幸存的则逐渐演化为今天的螯肢动物(蜘蛛、蝎子等)、多足动物(蜈蚣、马陆等)和泛甲壳动物(虾、蟹、昆虫等)。1984年澄江生物群的发现,为研究众多动物门类的早期演化、“寒武纪大爆发”的过程和机制等重要科学问题提供了大量珍贵素材。一直以来,研究者对澄江化石的形态学观察,主要局限于使用光学显微镜等传统成像技术,对化石表面所保存的二维结构的观察上,而保存在化石标本内部的动物身体结构信息,只能通过耗时且具破坏性的钢针修理等方法进行研究。正因为如此,研究者对早期节肢动物身体许多重要的精细结构(如口板)是否真正存在这一科学问题,只能通过分支分析等方法进行推测。即便推测准确,也无法对这类精细结构的真实形态进行展示。近期,在高精度显微CT 的帮助下,研究者用无损方式,对两枚呈三维立体保存的迷人林乔利虫幼虫的精细结构,进行任意角度的高精度复原与观察,使用计算机三维软件对它们的身体模型进行“虚拟解剖”并分离出身体的各部分结构进行研究。这两枚迷人林乔利虫幼虫体长分别为5 毫米及7 毫米。研究发现,迷人林乔利虫在体长仅为5 毫米的幼虫期,眼睛后长有一个明显的口板,口板表面还长有一对小突起。通过使用计算机三维软件复原,研究者对一枚体长仅为7 毫米的迷人林乔利虫幼虫化石的大附肢进行擦除后发现,在这一生长期,动物口板在其头部所占的比例有所减小,说明口板结构在该物种的个体发育进程中有可能存在不完全发育的现象。将将化石数据与现生节肢动物胚胎发育数据进行对比后,发现迷人林乔利虫作为螯肢类节肢动物(如蜘蛛、蝎子等)在寒武纪的代表,已经进化出明显的、与现生节肢动物口板同源的结构。这说明现生节肢动物中普遍存在的这一口前的突起构造,早在5.18 亿年前的干群螯肢动物中就已出现。(Current Biology,2020,30:3057-3061.e2)
猜你喜欢
心理学报(2021年6期)2021-06-10
中华耳科学杂志(2021年5期)2021-01-02
安徽史学(2020年6期)2020-12-07
湛江文学(2020年5期)2020-11-18
中华养生保健(2020年3期)2020-11-16
决策(2019年12期)2019-12-27
黄河·黄土·黄种人(华夏文明)(2018年11期)2018-12-05
文学少年(小学版)(2009年1期)2009-01-22
百家讲坛(2008年17期)2008-09-28
为了孩子(孕0~3岁)(2001年12期)2001-07-23
