
产α-环糊精葡萄糖基转移酶的菌株筛选、鉴定与酶学性质的初步研究
2021-04-01 03:27陶大炜宁喜斌
食品与发酵工业 2021年6期
陶大炜,宁喜斌,2,3,4*
1(上海海洋大学 食品学院,上海,201306)2(上海水产品加工及贮藏工程技术研究中心,上海,201306)3(农业部水产品贮藏保鲜质量安全风险评估试验室(上海),上海,201306)4(国家淡水水产品加工技术研发分中心(上海),上海,201306)
环糊精 (cyclodextrin,CD)通常由6个以上D-吡喃葡萄糖基通过α-1,4-糖苷键相连而成的,由于葡萄糖基个数的不同,又被分为α-CD、β-CD和γ-CD这3种,它们是应用最广泛的环糊精,也称基础环糊精,其中α-CD有6个葡萄糖基,β-CD有7个,而γ-CD有8个[1]。环糊精分子结构的外面部分具有亲水的特点,因为含有亲水基团,分子结构的内部相对就是疏水的,正因为其分子结构特殊,从而可以包络不同化合物,以改变它们物理和化学等性质,因此环糊精可以被广泛并且有效的利用,并运用于如食品工业、化妆品、药品、环保、化学分析与检测等诸多领域里[2-5]。
环糊精葡萄糖基转移酶(cyclodextrin glycosyltransferase,CGTase)可作用于淀粉等,使底物的葡萄糖基团发生转移,从而获得环糊精。CGTase可以催化4种不同的反应:歧化反应、环化反应、偶合反应和水解反应(其中前3种为3种转糖基反应)[6]。在工业应用中,主要是利用CGTase的环化反应作用,与淀粉等发生反应后生产环糊精。
目前大致可归纳为筛选产酶野生菌株进行发酵生产、结合基因库对产酶菌株的基因进行克隆表达、使用蛋白质工程技术对已知的基因片段进行改良这3种方法获取CGTase,其中产酶野生菌株的筛选是直接有效的,同时也是其他方法的基础[7]。在自然界中有很多菌株可生产CGTase,它们的分布很广泛,既存在于火山、沙石、温泉、油井等[8-11]环境,还存在于富含淀粉土壤的玉米、薯类、土豆、南瓜、甘蔗等[12]的种植地,同时这些野生菌株的种类众多。
国内外学者主要致力于筛选高产且性质稳定的野生菌株、提高酶的热稳定性及pH稳定性、优化菌株的产酶条件提高酶活性、物理化学诱变与在工程菌株中的异源表达提高酶活性等。MARTINS等[8]筛选出的包含黏琼脂芽孢杆菌(Bacillusagaradhaerens)所产CGTase酶活性较低,仅有0.31 U/mL,需要通过产酶条件优化等方式提高其酶活性。COELHOSL等[12]筛选出的芽孢杆菌(Bacillussp.)产β-CGTase的热稳定性较差,限制了其在工业上的应用能力。张佳瑜等[13]将来自于浸麻类芽孢杆菌(Paenibacillusmacerans)的产α-CGTase酶的基因进行克隆,并优化产酶条件,酶活性从1.9 U/mL提高到4.5 U/mL,说明基因的克隆表达和产酶条件优化有助于提高酶活性。雷新辉[5]筛选出的产酸克雷伯氏菌(Klebsiellaoxytoca)产α-CGTase酶活性为0.64 U/mL,经复合诱变处理后,活性提高到5.68 U/mL,说明诱变育种对于菌株的产酶活性有很大影响。
为了使环糊精的工业化生产能够实现,同时降低成本,找到产酶活性高、稳定性良好的野生菌株是关键。本试验从上海浦东新区新场镇某农场马铃薯、红薯、玉米种植田分离、筛选产α-CGTase的菌株,对其中1株酶活性较高的菌株进行形态学特征、生理生化试验鉴定和16S rRNA序列比对分析,对其酶学性质进行了初步研究,以期为进一步工业化开发利用性状稳定且高产α-CGTase的菌株提供借鉴,旨在为工业生产α-CGTase及其应用提供参考依据。
1 材料与方法
1.1 材料与试剂
1.1.1 菌种来源
2019年8月于上海浦东新区新场镇某农场马铃薯、红薯、玉米种植田(121.658 766 °E,31.052 923 °N)采集样品,用无菌采样器采集3~10 cm深的土壤样品,样品采集后放入无菌取样袋,并在2 h内送回试验室进行处理分析。
1.1.2 试剂
PCR扩增试剂盒、酵母浸粉、DNA提取试剂盒、可溶性淀粉、蛋白胨等, 上海生工生物工程股份有限公司;α-CD, Sigma(上海)公司;NaCl、K2HPO4·3H2O、MgSO4·7H2O,国药集团化学试剂有限公司;Na2CO3,上海柯灵斯试剂有限公司;酚酞、甲基橙,上海麦克林生化科技有限公司。
1.1.3 培养基
初筛培养基(g/L):可溶性淀粉10,蛋白胨5,酵母浸粉5,琼脂15,NaCl 5,Na2CO31,K2HPO4·3H2O 1,MgSO4·7H2O 0.2,酚酞 0.3,甲基橙 1,自然pH,121 ℃,0.1 MPa灭菌20 min。
复筛培养基(g/L):可溶性淀粉10,蛋白胨5,酵母浸粉5,NaCl 1,Na2CO30.5,K2HPO4·3H2O 1,MgSO4·7H2O 0.2,pH 7.0,121 ℃,0.1 MPa灭菌20 min。
发酵培养基及种子培养基,同复筛培养基。
1.2 仪器与设备
UV-1800PC紫外可见分光光度计,上海美谱达仪器有限公司;立式高压蒸汽灭菌锅,上海申安医疗器械厂;超净工作台,苏净集团安泰公司;THZ-300C恒温培养摇床、GHP-9 270隔水式恒温培养箱,上海一恒科学仪器有限公司;HWT-6B恒温水浴箱,天津市恒奥科技发展有限公司;CX41RF显微镜,东京奥林匹斯公司;高速台式离心机,上海安亭科学仪器厂;PTC-200 PCR仪、GelDoc XR凝胶成像系统,美国Bio-Rad公司。
1.3 试验方法
1.3.1 产α-CGTase菌株的初筛
分别取土豆、红薯、玉米种植田土样10 g加入到90 mL含玻璃珠的无菌水中,充分振荡40 min。取1 mL 上述混合液加入99 mL营养肉汤,37 ℃,170 r/min条件富集培养24 h。使用生理盐水进行梯度稀释,并将稀释液(100 μL)均匀涂布于初筛培养基,37 ℃培养48 h。以出现黄色透明圈作为筛选标准,将筛选出的菌株分离纯化,4 ℃条件下进行保藏。
1.3.2 产α-CGTase菌株的复筛
粗酶液的制备:发酵培养基装液量100 mL/250 mL,菌种接种量为4%的种子培养基,37 ℃、170 r/min摇床培养48 h,于10 000 r/min离心10 min后所得上清液即为粗酶液。
α-CGTase活力的测定:采用甲基橙褪色法测定酶的环化活力。在酸性(pH 1.1~1.4)条件下,α-CD与甲基橙发生反应,形成络合物,使溶液的吸光度下降。在一定范围内,吸光度的下降与α-CD的浓度存在着线性关系[4,14]。用50 mmol/L Na2HPO4-NaH2PO4缓冲液(pH 6.0)配制40 g/L的可溶性淀粉溶液做底物,取0.9 mL于试管中,加入适当稀释的粗酶液0.1 mL,均匀混合。50 ℃水浴反应10 min,加入1 mol/L盐酸溶液1 mL中止反应。再加入0.1 mmol/L 甲基橙溶液1 mL,于20 ℃静置15 min, 507 nm测定吸光度[15]。对应α-CD标准曲线计算浓度,1个酶活单位(U)定义为上述条件下1 min生成1 μmol α-CD所需的酶量。
1.4 菌种鉴定
1.4.1 形态学鉴定
观察纯化后菌株的菌落形态。挑取单菌落进行革兰氏染色镜检,观察菌体形态。
1.4.2 生理生化鉴定
根据相关文献[16-17],对菌株进行生理生化鉴定试验,主要包括水解淀粉试验、吲哚试验、硝酸盐还原、接触酶、分解酪氨酸、甘露醇、葡萄糖产酸产气、阿拉伯糖、棉籽糖利用等。
1.4.3 16S rRNA基因序列分析和构建系统发育树
根据细菌总DNA提取试剂盒说明提取菌株DNA。PCR扩增采用通用引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1 492R(5′-TACGGCTACCTTGTTACGACTT-3′)。PCR扩增产物送至上海生工生物公司测序,登入NCBI(National Center for Biotechnology Information),BLAST比对测序结果,获取同源性高的菌株的16S rRNA基因序列,使用MEGA-X软件中的邻接法(neighbor-joining,NJ)构建系统发育树,并对其分析鉴定[18]。通过Bankit将菌株鉴定的16S rRNA基因的部分序列向GenBank申请登录号(GenBank Accession)。
1.5 酶学性质初步研究
1.5.1 酶的最适作用温度
用Na2HPO4-NaH2PO4缓冲液(pH 6.0)稀释粗酶液和甲基橙溶液,设定20、30、40、45、50、55、60、70、80 ℃,9个反应温度。按照上述不同反应温度的条件,对各反应温度条件下的样品分别测定其α-CGTase活性,将酶活性最高者定义为100%,并计算相对酶活性。
1.5.2 酶的热稳定性
取适量无菌粗酶液分装于离心管,分别在40、45、50、55、60 ℃,5个保温温度条件下分别保温不同时间。按照上述不同保温温度和时间的条件,对各保温条件下的样品分别在不同保温时间下分别测定其α-CGTase活性,将未进行保温处理的酶活性定义为100%,并计算相对酶活性。
1.5.3 酶的最适作用pH
分别配制浓度为50 mmol/L的柠檬酸盐缓冲液(pH 3.0~5.0)、Na2HPO4-NaH2PO4缓冲液(pH 6.0~8.0)、甘氨酸-NaOH缓冲液(pH 8.0~10.0),Na2HPO4-NaOH缓冲液(pH 11.0~12.0)[15]。设定pH 3.0~12.0,10个反应pH。按照上述不同反应pH的条件,对各反应pH条件下的样品分别测定其α-CGTase活性,将酶活性最高者定义为100%,并计算相对酶活性。
1.5.4 酶的pH稳定性
先将粗酶液与不同pH值(3.0~12.0)的缓冲液混合,放置处理1、2 h。按照上述不同反应pH和处理时间的条件,对各pH处理条件下的样品分别在1、2 h下分别测定其α-CGTase活性,将未经过不同pH缓冲液处理的酶活性定义为100%,并计算相对酶活性。
1.5.5 金属离子对酶活力的影响
配制浓度为1 mmol/L的不同金属离子(K+、Zn2+、Mn2+、Cu2+、Mg2+、Ca2+、Fe2+、Co2+)溶液,分别取适量与粗酶液混匀。对添加不同金属离子的样品分别测定其α-CGTase活力,将未添加金属离子的酶液作为对照组,定义其酶活力为100%,并计算各组相对酶活力。
2 结果与分析
2.1 菌种筛选
初筛培养基因加入酚酞而呈红色,菌株所产α-CGTase可降解培养基中的可溶性淀粉形成环糊精,环糊精可包埋酚酞,使菌株周围呈现黄色透明圈,透明圈越大,说明菌株产α-CGTase越多[19]。据此从土壤中筛选出117株产α-CGTase的菌株,从中挑选出透明圈直径与菌落直径比值较大的菌株30株进行发酵产酶培养复筛,通过甲基橙褪色法分别测定酶活性,获得酶活性较高(表1)菌株TLLY7。本试验将菌株TLLY7作为出发菌株,进行后续试验,其酶性活为1.35 U/mL,这与其他学者筛选出的野生菌株的酶活性相似,例如陈龙然[4]筛选出的地衣芽孢杆菌(Bacilluslicheniformis)产α-CGTase的酶活性(0.95 U/mL)、陈龙军等[20]筛选出的嗜热芽胞杆菌(Gebacilliussp.)产α-CGTase的酶活性(0.66 U/mL)、叶学军[21]筛选出的地芽孢杆菌(Geobacilluscaldoxylosilyticus)产α-CGTase的酶活性(1.29 U/mL),低于他们进行复合诱变、产酶条件优化和基因的克隆与表达等方法后的酶活性,分别为2.28、5.3和6.18 U/mL,这说明筛选出活性高的野生菌株只是完成了第1步,只依靠野生菌株发酵有很大的局限性,需要结合其他方法提高酶活性,才有更大的工业应用潜力。

表1 产α-CGTase部分菌株的筛选结果Table 1 Screening results of α-CGTase producing strains
2.2 菌种鉴定
2.2.1 菌种的形态观察
菌株TLLY7为革兰氏阳性,菌体成杆状,有芽孢产生。图1-a为菌株TLLY7在初筛培养基上的菌落透明圈形态。图1-b为菌株TLLY7平板菌落形态,菌落呈黄白色,圆形隆起,不光滑,不透明,大小2~5 mm,有褶皱,边缘整齐。图1-c为菌株TLLY7经革兰氏染色后在油镜下的观察图。

a-初筛培养基菌落透明圈形态;b-平板菌落形态;c-细胞显微形态图1 菌株 TLLY7 菌落形态和在油镜下的细胞形态Fig.1 The colony morphology and cell morphology under oil immersion lens of strain TLLY7
2.2.2 生理生化鉴定
根据相关文献[16-17],对菌株TLLY7进行生理生化鉴定。表2为菌株TLLY7的生理生化鉴定结果。

表2 菌株TLLY7的生理生化特征Table 2 Biochemical characteristics of strain TLLY7
2.2.3 16S rRNA鉴定及系统发育树构建
图2为菌株TLLY7的PCR产物电泳条带图。由图2可知菌株TLLY7经PCR扩增后,条带约1 500 bp,扩增产物经测序得到的16S rRNA序列为1 480 bp,登录NCBI官网上传序列并进行BLAST比对,结果显示高于Ident值96%的菌株有100多株,说明较多菌株与菌株TLLY7序列相似,将它们序列下载后通过MEGA-X的NJ法构建系统发育树(图3)[18, 22]。由图3可发现菌株TLLY7和P.maceransNBRC 15 307T(NR 112729.1)在同一分支上,其相似性高达98.91%,说明两者同为类芽孢杆菌属,且Bootstrap检验值为100,说明发育树所表示的2种菌株之间的进化关系可信[23]。结合形态学、生理生化鉴定可确定菌株TLLY7为浸麻类芽孢杆菌(P.macerans)。将菌株TLLY7的核酸序列通过Bankit向GenBank申请的登录号为MT 705866。

图3 基于16S rRNA序列分析的菌株TLLY7的系统发育树Fig.3 Phylogenetic tree of strain TLLY7 based on 16S rRNA sequence analysis
2.3 菌株TLLY7酶学性质初步研究
2.3.1 α-CD标准曲线的绘制
根据标准方法绘制α-CD标准曲线。线性回归方程为y=0.546 1x-0.000 8,R2=0.999 1,表明线性良好。
2.3.2 酶的最适作用温度
图4为温度对TLLY7所产α-CGTase活性的影响。当反应温度在20~50 ℃,酶活性不断升高,从30 ℃开始酶活性升高的很快。温度高于50 ℃,酶活性开始下降。尤其是温度高于60 ℃后,酶活性下降的很快,在80 ℃时酶活性仅15%。温度在40~60 ℃,酶活性保持在80%以上。所以酶的最适作用温度为50 ℃,是中温酶。该酶的最适反应温度低于张晓磊[19]从黄海海域中筛选出的包含黏琼脂芽孢杆菌所产α-CGTase的最适反应温度(55 ℃),低于张昌志[24]从罗汉果中筛选出的解淀粉芽孢杆菌(Bacillusamyloliquefaciens)所产β-CGTase的最适反应温度(60 ℃),具有节约工业应用的能源,提高发酵效率的潜力。
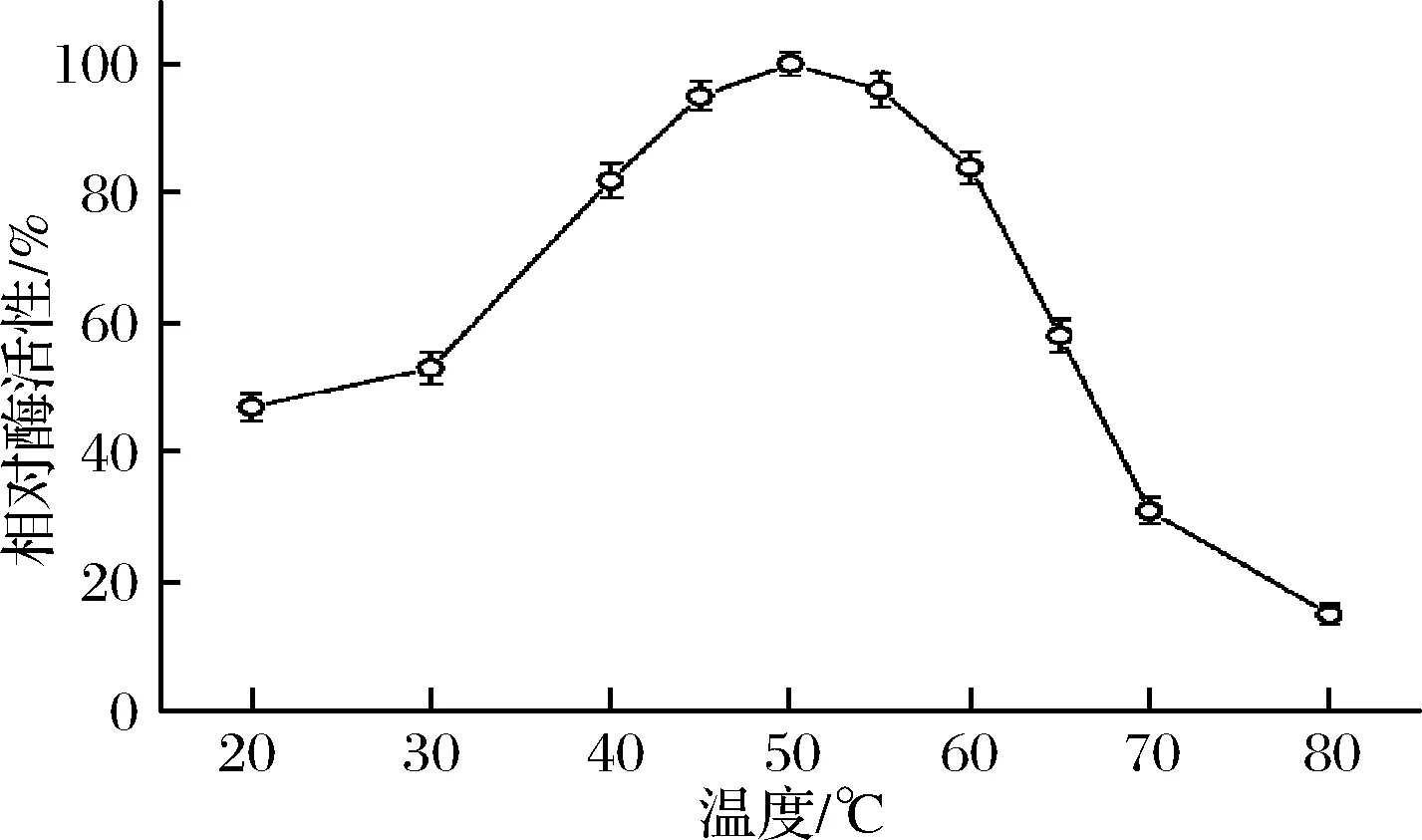
图4 温度对TLLY7所产α-CGTase活力的影响Fig.4 Effects of temperature on the α-CGTase produced by TLLY7
2.3.3 酶的热稳定性
图5为温度对TLLY7所产α-CGTase热稳定性的影响。在40 ℃下保温2 h酶活性保留82%,保温6 h保留30%的酶活性。在45 ℃保温1 h酶活性保留90%,保温5 h保留30 %酶活性。在50 ℃保温1.5 h,酶活性保留50%。在55 ℃保温0.75 h酶活性丧失50%。在60 ℃保温1 h酶活性保留28%。陈龙然[4]从淀粉厂泥土中筛选出的地衣芽孢杆菌所产α-CGTase在60 ℃保温1 h酶活性保留17%。本试验所产α-CGTase热稳定性相对较好,但热稳定性仍需要提高。因为酶的热稳定性不足会限制酶的工业化应用,所以还需要进行试验提高酶的热稳定性。

图5 温度对TLLY7所产α-CGTase稳定性的影响Fig.5 Effects of temperature on the stability of α-CGTase produced by TLLY7
2.3.4 酶的最适作用pH
图6为pH对TLLY7所产α-CGTase的影响。pH 3.0~4.0,酶活力很低。pH 4.0~6.0,酶活性升高的很快。pH>6.0,酶活性不断下降。pH 12.0时,酶活性仅5%。表明本试验所产α-CGTase的最适作用pH值为6.0,是弱酸性酶。

图6 pH对TLLY7所产α-CGTase活力的影响Fig.6 Effects of pH on the α-CGTase produced by TLLY7
2.3.5 酶的pH稳定性
图7为pH对TLLY7所产α-CGTase稳定性的影响。在pH 6.0~8.0,处理1 h酶活性保留95%以上,处理2 h酶活性保留90%以上。pH值<6.0或>8.0,酶活性明显下降。pH 3.0和12.0处理1 h分别保留9%和11%酶活性,处理2 h酶几乎失活。该酶在pH 10.0处理1 h保留77%酶活性。曹冬梅[25]从长白山温泉泥土中筛选出的高温芽孢杆菌所产β-CGTase在pH 10.0处理1 h保留60%酶活性。本试验的pH稳定性相对较好,具有更宽pH范围的应用前景。

图7 pH对TLLY7所产α-CGTase稳定性的影响Fig.7 Effects of pH on the stability of α-CGTase produced by TLLY7
2.3.6 金属离子对酶活的影响
由图8可知,对酶有不同程度的促进作用的是Zn2+、Mn2+和Ca2+,而对酶的促进作用最强的是Ca2+,相对酶活性约为空白对照的1.25倍。对酶有不同程度的抑制作用的是Cu2+、Fe2+和Co2+,其中对酶的抑制作用最强的是Cu2+,酶活性被抑制33%左右,可能由于离子结合蛋白的作用区域而抑制酶活。K+、Mg2+对酶活力的影响不是很明显。这与国内外学者筛选的菌株所产α-CGTase的性质相似,具有参考意义[26-27]。

图8 金属离子对α-CGTase活力的影响Fig.8 Effects of metal ions on the α-CGTase activity
3 结论与讨论
本试验利用甲基橙-酚酞平板法和甲基橙褪色法从上海浦东新区新场镇某农场土豆、红薯、玉米种植田分离出1株高产α-CGTase的菌株TLLY7,16S rRNA测序分析其基因序列,经BLAST比对后构建系统发育树,结合形态学特征、生理生化鉴定后确定该菌株为浸麻类芽孢杆菌,命名为P.maceransTLLY7。将菌株TLLY7的核酸序列通过Bankit向GenBank申请的登录号为MT 705866。
菌株TLLY7产α-CGTase的酶活性为1.35 U/mL,酶学性质的初步研究表明,50 ℃、pH 6.0为该酶的最适反应条件,为弱酸性中温酶。该酶在40~45 ℃保温1 h保留90%以上的酶活性,在60 ℃保温1 h保留27%酶活性,pH 6.0~9.0处理1 h保留95%以上的酶活性,处理2 h保留90%以上的酶活性,热稳定性和pH稳定性较好。Zn2+、Mn2+和Ca2+对酶有促进作用,Cu2+、Fe2+和Co2+对酶有抑制作用。其中Ca2+对酶的促进作用最强,Cu2+对酶的抑制作用最强。K+、Mg2+对酶活性的影响作用较小。
由于环糊精具有特殊的分子结构,可以包络不同化合物,改变它们理化性质,目前被应用到了多个领域,例如在食品领域中可以达到抗氧化、保护色素等效果,使使提高食品的贮藏时间、改善食品的保存;在医药领域中可以改变药物的性质使得药物刺激减少、提高药物的稳定性和降低毒副作用等效果,使药品更能被人们接受,还可被应用于化妆品、环保、化学分析与检测等诸多领域,因此对CGTase的产酶菌株进行分离纯化和酶学性质的研究是很有意义的[5]。本试验筛选出的产α-CGTase野生菌株可为未来的工作奠定基础,也为酶的分离纯化与工业化应用提供了依据。
猜你喜欢
食品安全导刊(2021年29期)2021-11-10
湖南饲料(2021年4期)2021-10-13
纺织科技进展(2021年3期)2021-06-09
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年8期)2019-11-23
新农业(2019年16期)2019-09-05
中国调味品(2019年3期)2019-03-18
中成药(2017年12期)2018-01-19
中成药(2017年5期)2017-06-13
