
拉萨河流域亏组山植物物种丰富度和群落特征研究
2021-06-10 09:01古桑群宗拉多郭应杰向明学武俊喜潘影张燕杰李石胜达文彦
广西植物 2021年3期
古桑群宗 拉多 郭应杰 向明学 武俊喜 潘影 张燕杰 李石胜 达文彦
摘 要:為研究草地植物群落、物种丰富度及其分布格局与影响因子之间的关系,该文以拉萨河流域林周县卡孜乡亏组山为研究地点,对山体垂直样带 (3 900~5 100 m) 植物群落特征、植物物种丰富度与各影响因子之间的关系进行了研究。研究区域共设置了13个样带 (每隔100 m设一个样带),每样带设置0.5 m × 0.5 m的5个样方进行植被调查,运用主成分分析 (PCA) 和双向指示种分析法 (TWINSPAN) 对植物群落进行排序和分类,运用冗余分析法 (RDA) 对群落及其分布格局与影响因子之间的关系进行分析,对植物物种丰富度与各因子之间的关系进行了回归分析。结果表明:该山体植物可分为3个群落类型,含7个群丛;影响区域植物群落物种组成和分布格局的主要环境因子为海拔,其次是坡度;物种丰富度与海拔、分种盖度呈单峰关系,与总盖度、坡度、地上生物量呈正相关。该研究结果为区域植物物种多样性和山地植物资源的保护和可持续利用提供了科学依据。
关键词: 物种丰富度, 环境因子, 分类, 排序, 群落结构, 亏组山
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2021)03-0372-12
收稿日期:2019-09-15
基金项目:国家重点研发计划项目 (2016YFC0502004-5);西藏大学研究生高水平人才培养项目 (2017-GSP-111);西藏科技重大专项 (XZ201801NA02) [ Supported by the National Key Research and Development Program of China (2016YFC0502004-5);High-Level Talents Training Program in Tibet University (2017-GSP-111); Key Program of Science and Technology for Tibet, China (XZ201801NA02) ]。
作者简介: 古桑群宗(1995-),硕士研究生,研究方向为草地植物生态学,(E-mail)1992899926@qq.com。
通信作者:拉多,博士,教授,硕士研究生导师,研究方向为植物生态学、植物生理学、古生态学,(E-mail)lhaduo@hotmail.com。
Plant species richness and community characteristics at Kuizu Mountain, Lhasa River basin
GUSANG Qunzong1, LA Duo1*, GUO Yingjie1,4, XIANG Mingxue1,2, WU Junxi2,3, PAN Ying2,3, ZHANG Yanjie3, LI Shisheng1, DA Wenyan1
( 1. Department of Biology, Tibet University, Lhasa 850000, China; 2. Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China; 3. Institute of Geographic Sciences and Resources, Chinese Academy of Sciences, Beijing 100101, China; 4. Yunnan Provincial Academy of Agricultural Sciences of Alpine Economic Plant Research Institute, Lijiang 674100, Yunnan, China )
Abstract:In order to study the relationship between grassland plant community, species richness, its distribution pattern and their influencing variables. Taking the Kuizu Mountain in Lhasa Basin as the study area, the plant community structure, species richness along the altitudinal gradient and their relationships with influential variables were studied. A total of 13 sample plots were set up at every 100 m altitude with five samples of 0.5 m × 0.5 m at each plot in the study area for vegetation survey. The plant community structure and distribution were analyzed by using TWINSPAN (two-way indicator species analysis) and PCA (principal component analysis). RDA (redundancy analysis) was played to analyze the correlations between plant community structure, distribution and environmental variables (altitude, slope). The regression analysis was used to analyze the correlations between species richness and abiotic (altitude, slope) and biotic (total coverage, separate coverage and aboveground biomass) variables. PCA and TWINSPAN results showed that the vegetation in the study area was divided into three communities including seven plant associations. The results of RDA showed that altitude was the main variable influencing the plant community composition and its distribution pattern followed by slope in the study area. The results of regression analysis showed that the species richness had a single-peak relationship with altitude and separate coverage, and was positively correlated with total coverage, slope and aboveground biomass. This study provides a scientific baseline for the conservation of plant species diversity and sustainable utilization of mountain plant resources in the study area.
Key words: species richness, environmental factor, TWINSPAN, PCA, community structure, Kuizu Mountain
青藏高原具有独特的高寒气候和复杂的自然地理条件,是一个具有全球性重要意义的生物多样性热点地区和生态脆弱区 (钟祥浩等,2006)。植物物种多样性的研究对区域生态系统的保护和其资源的永续利用以及环境评价等方面具有很重要的意义。影响物种多样性的因素很多,在一个特定的地点,海拔主导综合性环境因子产生垂直分异 (宋长宏,2013),它关联着温度、降雨量、土壤、气压、辐射能等多种环境因子 (赵海珍,2010),海拔对物种分布有深刻的影响,相关的研究可以为预测全球气候变化背景下的物种分布和生态系统的响应提供科学依据。现有的研究表明,高山草地植物丰富度与海拔之间存在三种不同的变化趋势,即呈正相关、负相关和单峰分布 (Jií & Miroslav,2002;杨元合等,2004;李素清和张金屯,2007;朱源等,2007;段敏杰,2011;栗文瀚等,2017)。可见,在不同研究区域、海拔跨度,研究结果有一定的差异,并且在影响物种丰富度与群落结构因子中,除了目前研究较为多的环境因子海拔以外,还有地形因子坡度等与物种丰富度和群落结构有重要的影响 (宋永昌,2001;金樑等,2007;王娟,2013;刘冠成等,2018)。植物盖度与生物量作为衡量植物群落保持水土功能 (郭忠升,2000) 和地面植物群落直观定量化的功能指标,能够反映区域生境质量的好坏 (Harris et al., 2014)。因此,在生态系统的结构和功能的形成上有重要的作用。在目前的研究中,关于生物量和植被盖度与植物物种丰富度关系的研究发现,物种丰富度与总盖度呈正显著相关关系,与分种盖度(唐志尧和方精云,2004;拉琼等,2014) 和生物量 (刘立斌等,2018) 呈单峰变化趋势。
區域草地植物物种分布格局及群落结构的研究能够为区域草地资源合理利用与保护提供科学依据。从已有的文献来看,拉萨河流域山地植物物种丰富度及群落结构在垂直梯度上的研究较多。但是,由于垂直尺度选择不同而出现不同的研究结果,因此对植被因子 (植物盖度、生物量等)、地形因子 (坡度) 与群落分布格局之间关系的研究却相对较少。然而,在地形多样化的青藏高原地区,关于地形因子与植物多样性的相关研究显得尤为重要。本文以拉萨河流域林周县卡孜乡亏组山作为研究地点,在海拔3 900~5 100 m的垂直梯度上进行植被调查,对植物物种丰富度与植被因子、地形因子之间的相互关系进行了初步研究,其结果对研究拉萨河流域大尺度山地植物物种丰富度与影响因子之间的关系具有一定的参考价值,为保护研究区域植物物种多样性和山地植物资源可持续利用提供科学依据。
1 研究区域与研究方法
1.1 研究区域
卡孜乡亏组山 (91°07′33″—91°07′19″ E、29°52′23″—29°49′50″ N) 位于西藏拉萨河支流澎波藏曲流域西南部,隶属于西藏自治区拉萨市林周县,距林周县10 km左右,其山体海拔跨度3 900~5 100 m之间,年日照时数大于3 000 h,年均温7.5~7.6 ℃,年均降水量在440 mm左右,主要集中在6—9月 (张晓庆等,2017;金艳梅等,2018), 研究区域常见植物有高山嵩草 (Kobresia pygmaea)、钉柱委陵菜 (Potentilla saundersiana)、青藏苔草 (Carex moorcroftii)、肉果草 (Lancea tibetica)、草沙蚕 (Tripogon bromoides) 等。
1.2 研究方法
1.2.1 样方调查 2018年7—9月,在海拔3 900~5 100 m的研究区域,每隔100 m设置一个样地,共13个样地,由于该区域的灌丛较少(只有2个样方灌丛盖度较大,分别为30%、46%,其余灌丛盖度在0.5%~22%之间),并且需要长期监测样地,为了减少人为破坏对样地的影响,每个样地设置0.5 m × 0.5 m的5个样方 (王长庭等,2005;郑伟等,2010),共65个样方,样方序号与海拔带相对应,如1~5号为3 900 m,6~10号为4 000 m,以此类推;样方设置时尽可能避开特殊小生境 (如大块石头、人和牲畜干扰严重的地方)。记录每个样方内的植物种类,并估算每个样方植被的总盖度与分种盖度 (样方内各物种盖度的总和)。用坡度仪记录每个样方的坡度,齐地面剪下植物,在80 ℃烘箱内烘干24 h后称干重,即为植物地上生物量,以下简称为生物量。
1.2.2 数据分析 采用双向指示种分析 (TWINSPAN-v3.2) 法对植物群落进行分类;运用CANOCO for Windows 4.5的主成分分析 (principal components analysis,PCA) 法对植物群落进行排序分析;采用冗余分析 (redundancy analysis,RDA) 法对植物群落与环境因子之间的关系进行了分析,同时采用SPSS19.0对植物物种丰富度与各因子间进行了回归分析。
2 结果与分析
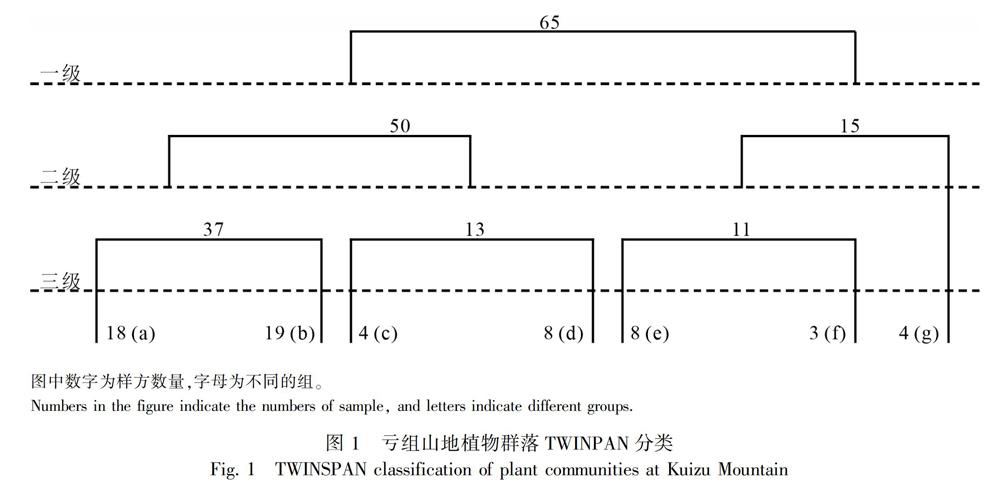
2.1 亏组山植物群落TWINSPAN 分类
TWINSPAN分类分析将亏组山65个样方分成7组,按照“外貌-生态学分类方法” (中国科学院青藏高原综合考察队,1980),并以依据群落生境特征的优势种和次优势中命名群丛类型,将其划分为7个群丛 (图1),具体如下:
青藏苔草+高山嵩草+藏西嵩草 (Kobresia deasyi) 群丛 (a):包括18个样方,主要分布海拔为4 800~5 100 m。在群丛水平上,以青藏苔草为优势种,次优势种有高山嵩草和藏西嵩草;主要的伴生种有钉柱委陵菜、圆穗蓼 (Polygonum macrophyllum)、中亚早熟禾 (Poalitwinowiana)、麻花艽 (Gentiana straminea)、弱小火绒草 (Leontopodium pusillum) 等。
雪层杜鹃 (Rhododendron nivale)+樱草杜鹃 (Rhododendron primuliflorum)+高山嵩草群丛 (b):共包括19个样方,主要分布海拔为4 400~4 800 m。在群丛水平上,以雪层杜鹃为优势种,次优势种为高山嵩草,灌木层的次优势种为樱草杜鹃;主要的伴生种有钉柱委陵菜、藏西嵩草、圆穗蓼、弱小火绒草、高山唐松草 (Thalictrum alpinum)、甘松 (Nardostachys jatamansi) 等。
丝颖针茅 (Stipa capillacea) +高山嵩草群丛 (c):共包括5个样方,主要分布海拔为4 300~4 400 m。在群丛水平上,丝颖针茅为优势种,次优势种为高山嵩草;伴生种有钉柱委陵菜、肉果草、藏橐吾 (Ligularia rumicifolia)、西伯利亚蓼 (Polygonum sibiricum) 等。
藏西嵩草+高山嵩草群丛 (d):包括8个样方,主要分布海拔为4 200~4 400 m。在群丛水平上,藏西嵩草为优势种,次优势中为高山嵩草;主要的伴生种有丝颖针茅、二裂委陵菜(Potentilla bifurca)、青藏苔草、肉果草等。
草沙蚕+藏橐吾群丛 (e):包括8个样方,主要分布在海拔3 900~4 000 m的干暖河谷底部。在群丛水平上,以草沙蚕为优势种,次优势种为藏橐吾;主要的伴生种有笔直黄芪 (Astragalus strictus)、肉果草、丝颖针茅等。
白草+小蓝雪花群丛 (f):包括3个样方,主要分布在海拔3 900~4 100 m的温度较高、较干旱的环境中。在群丛水平上,以白草 (Pennisetum centrasiaticum) 为优势种;主要的伴生种有笔直黄芪、二裂委陵菜、狼毒 (Stellera chamaejasme)、美叶藏菊 (Stellera chamaejasme)、蜜花毛果草 (Lasiocaryum densiflorum) 等。
蕨麻委陵菜 (Potentilla anserina) +二裂委陵菜群丛 (g):包括4个样方,分布海拔在4 100 m左右。在群叢水平上,以蕨麻委陵菜为优势种;主要的伴生种有青藏苔草、平车前 (Plantago depressa)、西伯利亚蓼、蜜花毛果草等。
2.2 亏组山植物物种和样方排序
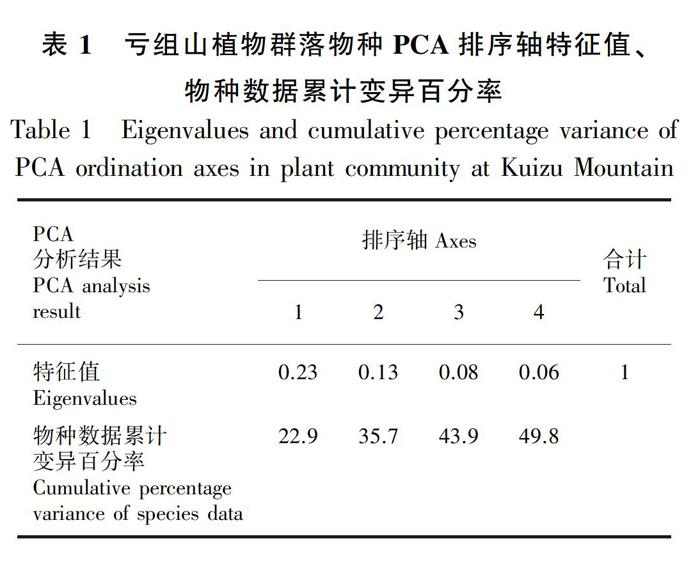
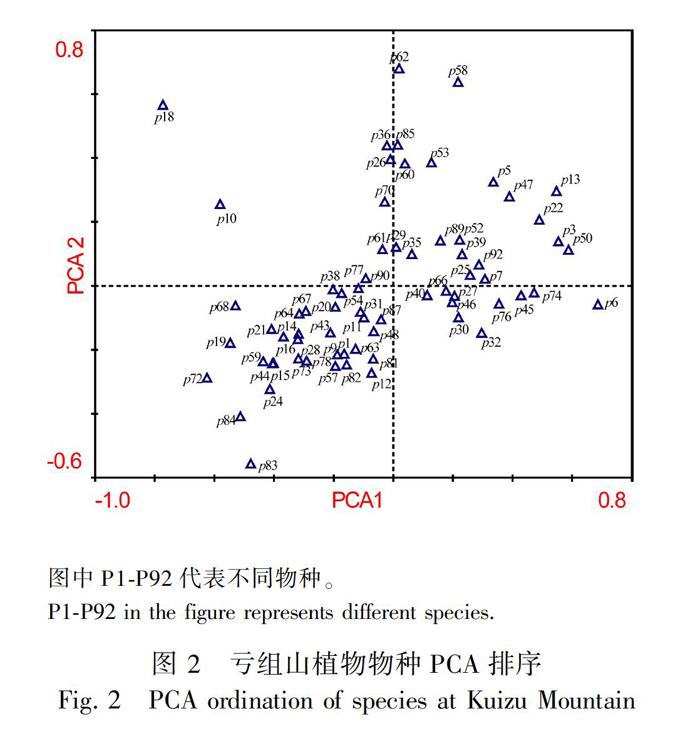
2.2.1 植物物种PCA排序 PCA分析结果 (表1) 显示,排序轴1的特征值λ=0.23,物种数据累计变异百分率为22.9%,排序轴1解释22.9%的物种变化;排序轴2的特征值λ=0.13,物种数据累计变异百分率为12.8%,与排序轴1共同解释了物种组成变化为35.7%;排序轴3和排序轴4分别解释了物种组成变化的8.2%、5.9%。这表明排序轴1和排序轴2为最能反映植物群落物种组成变化的影响因子梯度。
物种PCA排序结果 (图2)显示,高山嵩草、钉柱委陵菜、藏西嵩草、美丽风毛菊 (Saussurea pulchra)、线叶龙胆 (Gentiana farreri)、缘毛紫菀 (Aster souliei) 圆穗蓼等大多排在轴1的左下方,这些物种属高山草甸成分,分布于高海拔区域;而拉萨狗娃花 (Heteropappus bowerii)、藏橐吾、密花毛果草 (Lasiocaryum densiflorum)、二裂委陵菜、 白草、草沙蚕等大多排在轴1的右上方,这些物种通常出现在海拔较低、湿度较小的干暖河谷区域。丝颖针茅、肉果草、西藏荨麻 (Urtica tibetica) 等排在轴2的最上方,这些物种通常出现在干扰和退化程度较大的区域;而多刺绿绒蒿 (Meconopsis horridula)、樱草杜鹃、优越虎耳草 (Saxifraga egregia) 等排在轴2的下方,这些物种通常出现在湿度大、海拔较高的地方。
2.2.2 植物群落样方PCA排序 亏组山65个植物样方的PCA排序结果 (图3) 与TWINSPAN分类分析结果 (图1) 存在一定的差异。在PCA排序分析中将样方划分为3个类型,在TWINSPAN分类中分为7个类型,TWINSPAN分类中的a和b群丛在PCA排序中合为组成类型 (Ⅰ);c和d群丛合为组成类型 (Ⅱ);e、f和g群丛合为组成类型 (Ⅲ)。从PCA划分的3个类型内植物群丛的组成来看,Ⅰ、Ⅱ和Ⅲ分别为灌丛草甸、高寒草甸和温性草原的群丛组成成分。
2.2.3 植物群落样方RDA排序 从亏组山植物样方RDA分析结果 (图4,表2) 可以看出,RDA排序轴1的特征值λ1=0.16,物种与影响因子相关系数r=0.89,轴1解释了物种组成与影响因子之间关系的16.4%;轴2 的特征值λ2=0.03,物种与影响因子的相关系数r=0.72,与轴1累计解释了物种组成与影响因子之间关系的13.3%;轴3的特征值λ3=0.14,物种与影响因子的相关系数r=0,与轴3累计解释了物种组成与影响因子之间关系的33.1%;轴4的特征值λ4=0.09,物种与影响因子的相关系数r=0,与轴3累计解释了物种组成与影响因子之间关系的41.6%。第1轴与海拔和坡度相关系数分别为-0.88和-0.68,呈负相关;轴2与坡度和海拔相关系数分别为-0.46和0.07,与坡度负相关,与海拔正相关,但相关程度很小;轴3和轴4与环境梯度没有相关性。
从各因子之间的相关关系结果 (表3) 可以看出,在所调查的5个因子之间存在一定的相关性。其中,海拔与总盖度 (r=0.71)、分种盖度 (r=0.54)、生物量相关性不显著;坡度与总盖度 (r=0.55)、分种盖度 (r=0.41)、生物量 (r=0.27) 正显著相关。
2.3 物种丰富度分布格局
通过回归分析结果 (图5:a-e)发现,亏组山植物物种丰富度与海拔、分种盖度存在显著相关 (P<0.01),丰富度从低海拔到中、高海拔 (3 900~4 500 m) 逐渐上升,在4 600~4 700 m达到最高点之后开始缓慢下降,呈中间高、两边低的“单峰”格局,且该趋势解释量R2=0.447,能够解释44.7%的物种丰富度变化 (图5:a)。植物物种丰富度随着分种盖度先逐渐上升,当分种盖度在80%~100%之间时丰富度达到最大值,在分种盖度超过100%之后物种丰富度开始下降,也呈单峰变化趋势,并且该趋势能够解释36.1%的物种丰富度变化 (图5:c)。植物物种丰富度与坡度和总盖度存在显著相关 (P<0.01),呈直线上升趋势 (图5:b, d),并且该趋势能够分别解释41.6%和33.3%的物种丰富度变化。植物物种丰富度与生物量存在显著相关关系 (P<0.01),呈先上升后稳定的变化趋势 (图5:e),且每样方内地上生物量在10~30 g时物种丰富度值较高。
3 讨论与结论
3.1 群落特征和植被类型
TWINSPAN分类将亏组山65个样方分为7个组,分别为7种群丛。PCA样方排序分析将亏组山植物群落分为Ⅰ、Ⅱ和Ⅲ 3个组。对亏祖山的65个样方用不同的分类方法进行分析时发现,两种方法的分类结果有些差异。但是,从PCA排序进行分类的3个组和TWINSPAN分类的7个组间进行对比时发现两结果间却没有很大的冲突。如PCA分类的Ⅰ组里包括TWINSPAN分类里的两个群丛(a和b),为典型的灌丛草甸,Ⅱ为高山草甸(c和d),Ⅲ为高山草原(e,f和g) 三种不同的草地群落类型。
将对不同类型群丛所对应的样方信息进行分析发现,类型Ⅲ分布于海拔3 900~4 100 m,该区域温度较高,蒸发量大,人和动物的干扰较大的河谷底部;类型Ⅱ分布在海拔4 200~4 400 m之间,该区域水热条件处于比较理想的状态;类型Ⅰ分布的海拔范围为4 400~5 100 m,这一植被类型为山体的主要植被类型,是小型灌丛分布较普遍的区域。可见,在不同的区域,生境与环境条件不同,植被类型也不同。对每个植被类型样方对应的物种信息进行分析发现,Ⅰ内总计68物种,共有种为38种;Ⅱ内总计45个物种,共有种为20种;Ⅲ内总计43物种,共有种为10种。在PCA分析中,Ⅰ类型内a和b之间很难区分开来,呈连续状的分布,Ⅱ和Ⅲ也是如此。高山草地植物群落在垂直样带上的分布有明显的组成差异,但不同的群落类型之间有过渡区,群落之间存在连续性分布;在相对同一个样带内,由于人为、地形和地表径流等因子的影响,出现草地植物群落的斑块化,使得植物群落在不同生境间呈间断性分布。可见,PCA和TWINSPAN分类很好地揭示了植物群落在不同环境梯度上的连续性与间断性的分布特征。
物种PCA排序分析结果表明,轴1的左下方分布着以高海拔高山草甸物种成分为主,而轴1的右上方是以低海拔的干暖河谷出现的物种为主,结合样方的PCA排序结果,可见轴1明显的代表海拔梯度;轴2上方分布着丝颖针茅、肉果草、西藏荨麻等通常出现在干旱生境、干扰和退化较大的地方出现的物种,而轴2下方分布着多刺绿绒蒿、樱草杜鹃、优越虎耳草等湿度较大生境中出现的物种,可见轴2反映出以湿度为主的干扰等复合的环境梯度。这与拉多等(2016) 在拉萨河谷流域做的植被研究结果一致。综上所述,能够推断海拔、湿度和干扰梯度是影响该区域群落分布的主导因素。
RDA冗余分析结果显示,海拔梯度和坡度的变化对区域植物群落和物种分布的影响。其中,海拔对群落和物种分布格局的影响比坡度大,在藏北高原草地植物群落研究(王景升等,2016)中指出,湿度是影响该区域群落分布的主导因素;杨小林等(2010)在对拉萨河流域半干旱河谷地带植物群落研究和姚帅臣等(2018) 对同区域草地植物群落研究表明,海拔是影响群落分布的主导环境因子之一。除了海拔和湿度以外,还在本次研究中发现,坡度对群落组成与物种分布也有很重要的影响。坡度对群落的影响体现在不同的坡度所涉及的土壤厚度、土壤水分含量和单位面积内接受的能量等的差异影响着群落组成与物种的分布(宋永昌,2001)。
3.2 物种丰富度分布格局
植物物种丰富度在垂直样带上随环境梯度的变化而变化,由于研究区域、海拔跨度大小、坡度等环境因子的不同,因此呈现不同的结果 (彭闪江等,2003; Rahbek, 2005)。本研究表明,卡孜乡亏组山植物物种丰富度随海拔梯度大致呈中间高、两边低的单峰变化格局。与藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局类似 (段敏杰等,2011);在拉萨河谷山地灌丛草地物种多样性随海拔变化特征中也得到了相同的变化规律 (罗黎鸣等,2016)。低海拔带 (3 900~4 200 m),由于温度高、降雨量低、蒸散量高,在一定程度上,植物的生长主要受湿度的限制,且低海拔处还受人为干扰和牲畜放牧等影响较大,出现了笔直黄芪、藏橐吾、车前草等,因此植被稀疏、群落组成较为单一、植被呈现明显的退化状态。加之,在低海拔区域高原鼠兔的干扰强烈,不仅改变了高寒草甸植物群落内的物种数,而且还改变了原有物种在植物群落内的地位和作用 (金少红等,2017)。中海拔区域 (4 300~4 800 m),物种丰富度逐渐上升且达到最大值,这可能与该区域的水热条件与灌丛的分布有关,中海拔段相对于低海拔和高海拔区域,水热条件相对良好,给植物提供了相对理想的生长环境,并且该区域分布有小型灌木丛,灌丛既能隔离牲畜的采食和踩踏,间接地对灌丛下的植物起到了保护作用,又能有效地降低风速以及很好地截留掉落物,從而促进土壤有机质含量及其他物质在灌丛下的积累 (魏巍,2013;瞿王龙等,2015),导致物种丰富度的增高。在做调查时发现,中海拔段存在径流水,与低海拔区域相比湿度较大,土壤湿度作为植物生长的主要环境因子,对植被生长和分布起着很重要的作用 (武建双等,2012)。高海拔区域 (4 900~5 100 m),物种丰富度开始有降低的趋势,随着海拔的升高,温度逐渐成为草地植物生长的主要限制因子 (徐广平等,2005),在本研究区域的最高海拔5 100 m处,物种丰富度虽有下降的趋势,但维持在较高的水平,这可能与样方地理位置有关,最高的样点所处位置地势比较平缓,土壤比较厚,并且被周围的小山丘围绕着,可能形成了特殊的生境,导致维持较高的物种丰富度。
从物种丰富度随总盖度和分种盖度变化趋势可以看出,物种丰富度与总盖度呈正显著相关关系,与分种盖度呈单峰变化趋势,这与唐志尧和方精云(2004)以及拉琼等(2014)的研究结果一致,并且方精云等(2004)认为这种关系的出现可能与生境质量有关。植被盖度作为衡量植物群落保持水土功能 (郭忠升,2000) 和地面植物群落直观定量化的功能指标,能够反映区域生境质量的好坏 (Harris A et al., 2014)。在研究区域植物物种丰富度随总盖度一直上升,并且还有进一步的上升趋势,而物种丰富度较高的值都集中在盖度80%~100%,说明在这个盖度范围内物种以一种最合理的资源分配方式存在,当分种盖度超过100%时物种丰富度开始下降,可能是由于物种之间存在对光、水分、土壤养分等环境资源的竞争,而竞争能力小的物种就可能开始消失 (Bonser & Reader, 1995),从而导致物种丰富度减少。有关研究报道,植被盖度不仅是土壤侵蚀与水土流失的主要因子,而且还在区域尺度上影响着土壤水分的平衡 (赵文智,2002;Xin et al., 2004);植被通过根系吸水和水力提升起到水分调节作用 (李小雁,2011)。在不同等级植被盖度对土壤水分的影响研究中发现,植被盖度过大,土壤过度耗水,导致植被的严重演替与退化(孙祯元,1984)。
坡度在某种程度上决定了植物群落的分布格局 (刘金根和薛建辉,2010)。根据喻庆国等 (2007) 对坡度等级的划分,本研究区域在平坡至陡坡范围内 (3°~34°),坡度作为影响水土流失的主导因子 (曹梓豪等,2017),其大小会影响土壤水分含量和植物在单位面积接受的能量 (车明轩等,2016; 刘建华等,2017),并且在垂直样带上,影响着土壤养分含量,进而成为影响草本和灌木植物分布的主要地形因子 (袁铁象等,2014)。本研究区域,由于各样方之间和各样带之间的坡度差异较大,导致小环境条件的差异,从而影响着植物物种丰富度。本研究中,物种丰富度随坡度变化呈直线上升趋势,坡度较大的地方海拔相对较高,受到人畜干扰较小,并且在坡度较大的区域,低矮灌丛分布相对较多,灌丛具有一定的增加土壤肥力的作用 (Wezel et al., 2000),随着坡度的增加灌丛的多样性也增加 (韩杰等,2016),灌丛的出现为流苏虎耳草 (Saxifraga wallichiana)、甘松 (Nardostachys chinensis)、多刺绿绒蒿 (Meconopsis horridula) 等喜湿喜阴类丛下伴生植物种提供了适合的生境条件,从而对物种丰富度的提高具有积极作用。坡度较小的区域与村民居住地较近,牲畜及人为活动较为频繁,部分区域出现草地退化,禾草和杂类草优势较为明显,加之鼠兔的分布明显比坡度大的区域多,且水源的分布主要集中在低中海拔段(3 900~4 500 m),饮水点附近作为高山草地牲畜干扰较强的区域,其草地植物物种多样性与干扰强度呈正相关 (唐明坤等,2011;吕曾哲舟,2019)。
植物生物量作为生态系统的功能指标和获取能量的集中体现,在生态系统的结构和功能的形成上有重要作用。目前关于生物量与植物物种丰富度之间的关系研究结果较为多样,在群落分布格局研究中,植物物种多样性的变化由于群落类型的不同,其功能群以及植物生活型和种间竞争等差异对生物量有不同的影响 (陈加际等,2018)。本研究中,物种丰富度随生物量先增后稳定的变化趋势,生物量达到一定的值后,物种丰富度变化缓慢。这与杨元合等(2004)的研究结果一致,研究认为是生境条件差异及物种间的共生作用强所导致的。在本研区域之所以出现这一变化趋势可能是由于每个样带海拔和坡度等环境因子的不同而导致不同的小生境,且本研究区域为当地自然放牧地,每个区域的放牧强度也有所差异。此外,在垂直样带上草地有3种群落类型和7个群丛,不同的群落类型间,种间竞争和植物物种的组成等有所差异(刘立斌等,2018),其生物量也存在差异。如在样方内出现藏橐吾、黄苞南星 (Arisaema flavum) 等株高较高、叶片面积大和茎秆粗的物种,会直接影响样方内整体植物的生物量,因此生物量大的区域物种丰富度不一定高。随着放牧强度的增加,植物生物量会减少,但是物种丰富度的变化还得考虑具体的放牧压大小。高寒草地通过物种之间的信息与其功能间的联系来适应变化的环境,从而维持生产力的相对稳定 (朱桂丽等,2004)。因此,这两种变量间不是直接的因果关系,而是由环境因子、草地类型、放牧干扰强度等因子共同作用导致。
亏组山体植被在群落水平上可分为3个类型,各类型间具有连续性分布特点,群丛水平上分为7个类型,具有间断性分布特点;群落在垂直样带上的分布格局主要受海拔的影响,其次为坡度。在研究区植物物种丰富度随海拔变化呈单峰分布格局,与坡度呈正相关。植物物种丰富度对两种盖度变化的响应存在一定的差异,其与分盖度呈单峰分布格局,与总盖度呈正相关,植物物种丰富度与生物量呈正相关。山体植物群落组成,物种丰富度及其分布,不仅受环境因子的影响,而且还与总盖度、分盖度和生物量等生物因子之间存在显著的关系。由于本研究中未考虑到土壤养分、降雨量、坡向等对区域植被的影响,因此在PCA排序分析推断的湿度因子未能在RDA冗余分析中得到验证,有待于今后进一步地研究。
致谢 感谢中国科学院地理研究所国家自然科学基金面上項目(31570460)和“复旦大学钟扬教授基金”对本研究的资助,以及卡孜乡当地农牧民对本研究的大力支持与配合,特此致谢。
参考文献:
BONSER S, READER RJ, 1995. Plant competition and herbivory in relation to vegetation biomass [J]. Ecology,76(7): 2176-2183.
CAO ZH, ZHAO QH, DING SY, et al., 2017. Effect of slope gradient and vegetation cover on sediment yielding characteristics of the riparian slope [J]. J Nat Resour,
32(11): 1892-1904. [曹梓豪, 赵清贺, 丁圣彦, 等, 2017. 坡度和植被盖度对河岸坡面侵蚀产沙特征的影响 [J]. 自然资源学报, 32(11): 1892-1904.]
CHE MX, GONG YB, NAEEM KM, et al., 2016. Effects of different rainfall intensities and slope gradients on water and soil conservation of straw mulch [J]. J Soil Water Conserv,30(2): 131-135. [车明轩, 宫渊波, MUHAMMAD NAEEM KHAN, 等, 2016. 不同雨强、坡度对秸秆覆盖保持水土效果的影响 [J]. 水土保持学报, 30(2): 131-135.]
CHEN JJ, CHANG SH, WANG ZF, et al., 2018, The relationship between plant species diversity and above-ground biomass in a typical steppe in Abaga Banner [J]. Pratacul Sci, 35(9): 2068-2078. [陳加际, 常生华, 王召锋, 等, 2018. 阿巴嘎旗典型草原植物物种多样性与地上生物量的关系 [J]. 草业科学, 35(9): 2068-2078.]
Comprehensive Scientific Expedition Team of Chinese Academy of Sciences in Qinghai-Tibet Plateau, 1988. Tibetan Vegetation [M]. Beijing: Science Press: 90-228. [中国科学院青藏高原综合考察队, 1980. 西藏植被 [M]. 北京:科学出版社: 90-228.]
DUAN MJ, GAO QZ, GUO YQ, et al., 2011. Effects of altitude on plant productivity and species diversity in alpine meadows of northern Tibet [J]. Pratacul Sci, 28(10):1845-1850. [段敏杰, 高清竹, 郭亚奇, 等, 2011. 藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局 [J]. 草业科学, 28(10): 1845-1850.]
GUO ZS, 2000. Three coverages in soil and water conservation vegetation construction: Potential coverage, critical coverage and effective coverage [J]. Bull Soil Water Conserv,(2): 60-62. [郭忠升, 2000. 水土保持植被的有效盖度、临界盖度和潜势盖度 [J]. 水土保持通报, (2): 60-62.]
HARRIS A, CARR A, DASH J, et al., 2014. Remote sensing of vegetation cover dynamics and resilience across southern Africa [J]. Int J Appl Earth Obs Geoinf, 28: 131-139.
JIN L, DU XG, HOU FJ, et al., 2007. Effects of hillside slopes on plant community succession of re-natural farmland in the Loess, Plateau [J]. Pratacul Sci, 24(7): 66-71. [金樑, 杜晓光, 侯扶江, 等, 2007. 黄土高原山地坡度对退耕农田生态系统自然植被演替初期的影响 [J]. 草业科学,24(7): 66-71.]
JIN SH, LIU T, PANG XP, et al.,(2017). Effects of plateau pika(Ochotona crzoniae) disturbances on plant species diversity and above ground plant biomass in a Kobresia pygmaea meadow in the Qinghai Lake Region [J]. Acta Pratacul Sin, 26(5): 29-39. [金少红, 刘彤, 庞晓攀, 等, 2017. 高原鼠兔干扰对青海湖流域高山嵩草草甸植物多样性及地上生物量的影响 [J]. 草业学报, 26(5):29-39.]
JIN YM, TIAN FY, WU JX, et al., 2018. Effects of indoor feeding on weight gain and wool mass of Pengbo semi-fine wool sheep during winter-spring period in Lhasa River Valley [J]. J Plat Agric, 2(4):347-353. [金艳梅, 田发益, 武俊喜, 2018. 冬春季舍饲对拉萨河谷彭波半细毛羊增重和产毛量的影响 [J]. 高原农业, 2(4): 347-353.]
JI D, MIROSLAV , 2002. Altitudinal changes in composition and structure of mountain temperate vegetation: a case study from the Western Carpathians [J]. Plant Ecol, 158(2): 201-221.
LA D, ZHANG YJ, LIU J, 2016. Spatial distribution patterns and environmental interpretation of plant species richness in the Lhasa Valley, Tibet [J]. Acta Pratacul Sci, 25(10): 202-211. [拉多, 张燕杰, 刘杰, 等, 2016. 拉萨河谷植物物种丰富度空间分布格局及其环境解释 [J]. 草业学报,25(10): 202-211.]
LA Q, ZHA XCR, ZHU WD, et al., 2014. Plant species-richness and association with environmental factors in the riparian zone of the Yarlong Zangbo River of Tibet, China [J]. Biodivers Sci, 22(3): 337-347. [拉琼, 扎西次仁, 朱卫东, 等, 2014. 雅鲁藏布江河岸植物物种丰富度分布格局及其环境解释 [J]. 生物多样性, 22(3): 337-347.]
LI SQ, ZHANG JT, 2007. Ecological analysis of subalpine meadow in Yunding Mountain, Shanxi [J]. Geogr Res,26(1): 83-90. [李素清, 张金屯, 2007. 山西云顶山亚高山草甸群落生态分析 [J]. 地理研究, 26(1): 83-90.]
LI WH, GAN ZZB, CAO XJ, et al., 2017. Effects of altitude on plant productivity and species diversity in alpine meadows of northern Tibet [J]. Acta Pratacul Sin, 26(9): 200-207. [栗文瀚, 干珠扎布, 曹旭娟, 等, 2017. 海拔梯度对藏北高寒草地生产力和物种多样性的影响 [J]. 草业学报, 26(9): 200-207.]
LI XY, 2011. Soil-vegetation-hydrology coupling, response and adaptation mechanism in arid areas [J]. Chin Sci Sin(Terrae), 41(12): 1721-1730. [李小雁, 2011. 干旱地区土壤-植被-水文耦合、响应与适应机制 [J]. 中国科学:地球科学, 41(12): 1721-1730.]
LIU GC, HUANG YX, WANG QG, et al., 2018. Effects of environmental factors on plant species diversity: Research progress [J]. Chin Agric Sci Bull, 34(13): 83-89. [刘冠成, 黄雅曦, 王庆贵, 等, 2018. 环境因子对植物物种多样性的影响研究进展 [J]. 中国农学通报, 34(13):83-89.]
LIU JG, XUE JH, 2010. Impacts of slope gradient on the characteristics of Vetiveria zizanioides community in early-stage [J]. Grassl Turf, 30(2): 56-61. [刘金根, 薛建辉, 2010. 坡度对香根草护坡植物群落早期特征的影响 [J]. 草原与草坪, 30(2): 56-61.]
LIU JH, LI ML, SU JB, et al., 2017. Effects of soil organic matter content and terrain slope on soil moisture, crop yield [J]. Hunan Agric Sci,(1): 16-18. [刘建华, 李铭亮, 苏剑波, 等, 2017. 有机质含量、地形坡度对土壤保水效果及作物产量的影响 [J]. 湖南农业科学,(1): 16-18.]
LIU LB, ZHONG QL, NIJ, et al., 2018. Allometric function-based root biomass estimate of woody plants in a karst evergreen and deciduous broadleaf and mixed forest in central Guizhou Province, southwestern China [J]. Acta Ecol Sin, 38(24): 8726-8732. [刘立斌, 钟巧连, 倪健, 等, 2018. 基于生物量回归方程估算黔中喀斯特常绿落叶阔叶混交林木本植物的根系生物量 [J]. 生态学报,38(24): 8726-8732.]
LIU ZZZ, HUANG XX, WANG XY, et al., 2019. Disturbance pattern of alpine meadow in Yak Meadow Park, Jade Dragon Mountain [J]. J Natur Resour, 34(6): 1223-1231. [吕曾哲舟, 黄晓霞, 王琇瑜, 等, 2019. 玉龙雪山牦牛坪高山草甸的干擾格局分析 [J]. 自然资源学报,34(6): 1223-1231.]
LUO LM, WU JS, YU CQ, et al., 2016. A comparative study of methods for monitoring plant diversity in montane shrub grassland in the Lhasa River Basin [J]. Acta Pratacul Sin, 25(3): 22-31. [罗黎鸣, 武建双, 余成群, 等, 2016. 拉萨河谷山地灌丛草地植物多样性监测方法的比较研究 [J]. 草业学报, 25(3): 22-31.]
PENG SJ, HUANG ZY, XU GL,et al., 2003. Effects of habitat heterogeneity on forest community diversity in Dinghushan Biosphere Reserve [J]. Guihaia, 23(5): 391-398. [彭闪江, 黄忠良, 徐国良, 等, 2003. 生境异质性对鼎湖山植物群落多样性的影响 [J]. 广西植物, 23(5): 391-398.]
QU WL, YANG XP, ZHANG CT, et al., 2015. Shrub-mediated “fertile island” effects in arid and semi-qrid grassland [J]. Acta Pratcacul Sin, 24(4): 201-207. [瞿王龙, 杨小鹏, 张存涛, 等, 2015. 干旱、半干旱地区天然草原灌木及其肥岛效应研究进展 [J]. 草业学报, 24(4): 201-207.]
RAHBEK C, 2005. The role of spatial scale and the perception of large-scale species-richness patterns [J]. Ecol Lett,8(2): 224-239.
SONG YC, 2001. Vegetation Ecology [M]. Shanghai: East China Normal University Press: 171-176. [宋永昌, 2001. 植被生態学 [M]. 上海:华东师范大学出版社:171-176.]
SONG ZH, 2013. Effect of different altitudes on kobresia pygmaea community [J]. J Anhui Agric Sci, 41(11):4709-4711. [宋长宏, 2013. 不同海拔梯度对小嵩草植物群落的影响 [J]. 安徽农业科学, 41(11): 4709-4711.]
SUN ZY, 1984. Preliminary discussion on the relationship between vegetation coverage and water in Yulin sandy land [J]. J Beijing For Univ,(3): 29-35. [孙祯元, 1984. 对榆林沙地植被盖度和水分关系的初步探讨 [J]. 北京林学院学报, (3): 29-35.]
TANG MK, LI MF, ZHAO J, et al., 2011. Disturbance pattern of alpine meadow in Yak Meadow Park, Jade Dragon Mountain [J]. Guihaia, 31(6): 775-781. [唐明坤, 李明富, 赵杰, 等, 2011. 四川若尔盖县不同退化程度高寒草地群落比较研究 [J]. 广西植物, 31(6): 775-781.]
TANG ZY, FANG JY, 2004. A review on the elevational patterns of plant species diversity [J]. Biodivers Sci, 12(1): 20-28. [唐志尧, 方精云, 2004. 植物物种多样性的垂直分布格局 [J]. 生物多样性, 12(1): 20-28.]
WANG CT, LONG RJ, SHI JJ, et al., 2005. Study on the change of vertical structure and productivity of artificial grassland community sowed in different year in alpine area [J]. Chin J Grassl, 27(5): 16-21. [王长庭,龙瑞军, 施建军, 等, 2005. 高寒地区不同建植期人工草地群落垂直结构和生产力变化的研究 [J]. 中国草地学报, 27(5): 16-21.]
WANG J, 2013. Study on the relation between geological background and plant diversity of Gongbu nature reserve in Tibet [D]. Chengdu: Chengdu University of Technology: 118. [王娟, 2013. 西藏工布自然保护区地质背景与植物多样性关系研究 [D]. 成都: 成都理工大学: 118.]
WANG JS, YAO SC, PU Q, et al., 2016. Quantitative classification and ordination of grassland communities on the Northern Tibetan Plateau [J]. Acta Ecol Sin, 36(21): 6889-6896. [王景升, 姚帅臣, 普穷, 等, 2016. 藏北高原草地群落的数量分类与排序 [J]. 生态学报, 36(21): 6889-6896.]
WEI W, 2013. Plant diversity pattern and interface characteristics of shrubland-grassland ecotone in Eastern Qilian Mountains [D]. Gansu: Gansu Agricultural University: 63. [魏巍, 2013. 东祁连山灌—草交错带植物多样性格局及界面因子特征研究 [D]. 甘肃: 甘肃农业大学: 63.]
WEZEL A, RAJOT JL, HERBRIG C, et al., 2000. Influence of shrubs on soil characteristics and their function in sahelian agro-ecosystems in semi-arid Niger [J]. J Arid Environ,44(4): 383-398.
WU JS, LI XJ, SHEN ZX, et al., 2012. Species diversity distribution pattern of alpine grasslands communities along a precipitation gradient across Northern Tibetan Plateau [J]. Acta Pratacul Sin, 21(3): 17-25. [武建雙, 李晓佳, 沈振西, 等, 2012. 藏北高寒草地样带物种多样性沿降水梯度的分布格局 [J]. 草业学报, 21(3): 17-25.]
XIN RL, FENG YM, HONG LX, et al., 2004, Long-term effects of revegetation soil water content of and dunes in arid region of Northern China [J]. J Arid Environ, 57(1):1-16.
XU GP, ZHANG DG, PU XP, et al., 2005. Two-dimensional polar ordination of plant community of alpine grassland in Eastern Qilian mountains [J]. Grassl Turf,(3): 42-45. [徐广平, 张德罡, 蒲小鹏, 等, 2005. 东祁连山高寒草地植物群落二维极点排序 [J]. 草原与草坪, (3): 42-45.]
YANG XL, ZHAO KT, MA HP, et al., 2010. Ecological studies on vegetation quantity in the Semi-Arid Valley region of Lasa [J]. Sci Silv Sin, 46(10): 15-22. [杨小林, 赵垦田, 马和平, 等, 2010. 拉萨半干旱河谷地带的植被数量生态研究 [J]. 林业科学, 46(10): 15-22.]
YANG YH, RAO S, HU HF, et al., 2004. Plant species richness of alpine grasslands in relation to environmental factors and biomass on the Tibetan Plateau [J]. Biodivers Sci, 12(1): 200-205. [杨元合, 饶胜, 胡会峰, 等, 2004.青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系 [J]. 生物多样性, 12(1): 200-205.]
YAO SC, WANG JS, DING LB, et al., 2018. Quantitative classification and ordination of grassland communities in Lhasa River Valley [J]. Chin Acta Ecol Sin,38(13): 4779-4788. [姚帅臣, 王景升, 丁陆彬, 等, 2018. 拉萨河谷草地群落的数量分类与排序 [J]. 生态学报,38(13):4779-4788.]
YU QG, CAO SW, DENG XQ, et al., 2007. Study on the spatial distribution of vegetation along the gradients in Nuozhadu Nature Reserve of Yunnan Province [J]. J Anhui Agric Sci, 35(28): 8994-8996. [喻庆国, 曹顺伟, 邓喜庆, 等, 2007. 云南糯扎渡自然保护区植被沿坡度空间分异研究 [J]. 安徽农业科学, 35(28): 8994-8996.]
YUAN TX, ZHANG HP, OU ZY, et al., 2014. Effects of topography on the diversity and distribution pattern of ground plants in karst montane forests in Southwest Guangxi, China [J]. Chin J Appl Ecol, 25(10): 2803-2810. [袁铁象, 张合平, 欧芷阳, 等, 2014. 地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响 [J]. 应用生态学报, 25(10): 2803-2810.]
ZHANG XQ, LUO LM, JIN YM, et al., 2017. Changes in nutrients of dominant species on montane shrub grassland in the Lhasa River Basin [J]. Chin J Grassl,39(3): 90-95. [張晓庆, 罗黎鸣, 金艳梅, 等, 2017. 拉萨河谷山地灌丛草地优势种营养物质含量变化规律 [J]. 中国草地学报,39(3): 90-95.]
ZHANG YF, ZHAO QH, DING SY, et al., 2017. Effects of slope gradient and vegetation coverage on hydrodynamic characteristics of overland flow on silty riparian slope [J]. Chin J Appl Ecol, 28(8): 2488-2498. [张祎帆, 赵清贺,丁圣彦, 等, 2017. 坡度和植被盖度对粉砂质河岸带坡面流侵蚀水动力特征的影响 [J]. 应用生态学报, 28(8):2488-2498.]
ZHAO HZ, LI WH, MA AJ, et al., 2010. Evaluation of ecological services value of mountain shrubby-grassland ecosystem in Lhasa-river valley region — A case study of Dazi County [J]. Pratacul Sci, 27(12): 27-31. [赵海珍, 李文华, 马爱进, 等, 2010. 拉萨河谷山地灌丛草地生态系统服务价值评价——以拉萨达孜县为例 [J]. 草业科学, 27(12):27-31.]
ZHAO WZ, 2002. Impact of plantation on spatial heterogeneity of soil moisture in Horqin sandy land [J]. Acta Pedol Sin,39(1): 107-113. [赵文智, 2002. 科尔沁沙地人工植被对土壤水分异质性的影响 [J]. 土壤学报, 39(1): 107-113.]
ZHENG W, ZHU JZ, PAN CD, 2010. Effect of grazing disturbance on plant functional group and community structure of meadow community in Kanas Scenic Area [J]. Chin J Grassl, 32(1): 92-98. [郑伟, 朱进忠, 潘存德, 2010. 放牧干扰对喀纳斯草地植物功能群及群落结构的影响 [J]. 中国草地学报, 32(1): 92-98.]
ZHONG XH, LIU SZ, WANG XD, et al., 2006. A research on the protection and construction of the state ecological safe shelter zone on the Tibet Plateau [J]. Mt Res, 24(2): 129-136. [钟祥浩, 刘淑珍, 王小丹, 等, 2006. 西藏高原国家生态安全屏障保护与建设 [J]. 山地学报, 24(2):129-136.]
ZHU GL, LI J, WEI XH, et al., 2017. Longitudinal patterns of productivity and plant diversity in Tibetan alpine grasslands [J]. J Natur Resour, 32(2): 210-222. [朱桂丽, 李杰, 魏学红, 等, 2017. 青藏高寒草地植被生产力与生物多样性的经度格局 [J]. 自然资源学报, 32(2): 210-222.]
ZHU Y, KANG MY, LIU QR, et al., 2007. Relationship between biodiversity and aboveground biomass in alpine meadow on Mt. Helan, China [J]. J Appl Environ Biol,13(6): 771-776. [朱源, 康慕谊, 刘全儒, 等, 2007. 贺兰山高山草甸生物多样性和地上生物量的关系 [J]. 应用与环境生物学报, 13(6): 771-776.]
(责任编辑 蒋巧媛)
猜你喜欢
科普童话·学霸日记(2020年1期)2020-05-08
小天使·一年级语数英综合(2019年2期)2019-01-10
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
中国市场(2016年36期)2016-10-19
河北渔业(2015年10期)2015-10-15
河北渔业(2015年7期)2015-07-21
河北渔业(2015年6期)2015-06-24
