
干旱胁迫对细穗柽柳幼苗生长和生理生化指标的影响
2021-06-28 00:35胡杨李钢铁李星贾守义
中国农业科技导报 2021年6期
胡杨, 李钢铁*, 李星, 贾守义
(1.内蒙古农业大学沙漠治理学院, 呼和浩特 010018; 2.巴彦淖尔市林业科学研究所, 内蒙古 巴彦淖尔 015000 )
水分匮乏是我国西北干旱、半干旱地区抑制植物生长的主要因素[1],频发的高温天气和水资源短缺严重影响植物正常发育[2]。因此,研究植物抗旱性、筛选抗旱植物对于干旱、半干旱地区植物资源的发展尤为重要。
细穗柽柳(TamarixleptostachysBunge)属于柽柳科柽柳属灌木,生长于我国西北部荒漠、半荒漠地区,主要分布于新疆、青海、甘肃、宁夏和内蒙古等地。细穗柽柳的根系发达,防风固沙效果显著;因其具有极强的耐旱和耐盐碱能力,是荒漠、半荒漠盐碱和沙漠等地的理想造林树种;同时,种植细穗柽柳有良好的生态效益。细穗柽柳除具备以上特点外,还是柽柳种中艳丽的多花柽柳,是盐碱地造林的优质绿化及水土保持树种[3]。
目前,对细穗柽柳的研究大多集中在播种育苗技术[4]及药用化学成分[5-6]等方面,但关于细穗柽柳的抗旱机理探究尚未见报道。鉴于此,本研究通过设置不同梯度的干旱胁迫处理,分析探讨了干旱胁迫下细穗柽柳幼苗若干生长指标及主要生理生化特性对干旱胁迫的响应,旨在探明细穗柽柳幼苗的抗旱机理,综合分析其幼苗耐旱性潜力,为其在干旱地区适地适树大面积栽培提供依据。
1 材料与方法
1.1 试验材料
试验材料选用内蒙古磴口立地条件相同的2 a生细穗柽柳扦插苗。2018年7月10日将长势整齐、生长健壮、高度基本一致的细穗柽柳嫩枝小苗移栽于盆中。每盆定植健壮幼苗1株,试验所用土壤为采种地土壤(沙壤土),土壤经烘干(105 ℃)至恒重,每盆均装入等量过筛的混合土6.2 kg。土壤田间持水量为19.69%,容重1.46 g·cm-3。栽培容器为上口径25 cm、下口径15 cm、高19 cm的塑料花盆(带底盘)。从移栽当日至进行试验时,为确保幼苗健康生长,促进根系恢复,在温室内将土壤水分含量维持在田间持水量的80%[7]。
1.2 试验设计
试验在内蒙古自治区呼和浩特市内蒙古农业大学苗圃温室大棚内进行,土壤干旱胁迫试验于2018年9月10日开始,于11月8日结束。采用盆栽方法人工模拟土壤干旱条件,设置4个水分胁迫处理,对照(CK):土壤含水量为田间持水量的80%,实际土壤含水量保持在15.75%±0.98%;轻度干旱胁迫(T1):土壤含水量为田间持水量的60%,实际土壤含水量保持在11.81%±0.35%;中度干旱胁迫(T2):土壤含水量为田间持水量的40%,实际土壤含水量保持在7.88%±0.69%;重度干旱胁迫(T3):土壤含水量为田间持水量的20%,实际土壤含水量保持在3.94%± 0.41%。每个处理3次重复,每个重复4盆植株。采用称重法控制土壤实际含水量,每日下午17∶30用电子计价秤(正常载量15 kg,感知重量5 g)称重并补水,调节土壤含水量至各干旱胁迫处理范围内。试验材料干旱胁迫60 d后,采集新梢部自顶端向下3~4节上的功能叶片用于叶片各生理指标的测定。
1.3 测定指标及方法
1.3.1苗高及地径测定 分别于干旱胁迫处理前和处理后60 d,用卷尺测量地面至植株顶部的株高,用游标卡尺测定地径(植株地基部茎秆的直径),每个处理组测定12株。
1.3.2生理指标测定 相对含水率采用水饱和法进行测定;水势用小液流法进行测定;叶绿素(chlorophyll,Chl)含量用95%的酒精提取叶绿素,再用紫外可见分光光度计(UV759、上海奥普勒仪器有限公司)测定;测定丙二醛(malondialdehyde,MDA)含量用硫代巴比妥酸法;用电导仪测定叶片细胞膜透性;叶片游离脯氨酸(proline,Pro)含量用磺基水杨酸法测定;过氧化氢酶(catalase,CAT)活性用滴定法测定;过氧化物酶(peroxisome,POD)活性用愈创木酚法测定;超氧化物酶(superoxide dismutase,SOD)活性用氮蓝四唑法测定;可溶性糖(soluble sugar,SS)含量的测定采用蒽酮比色法;可溶性蛋白质(soluble protein,SP)含量采用G-250比色法测定[8]。各生理指标测定均设置3次重复。
1.4 数据处理
用SPSS 24.0软件中的One-way ANOVA比较不同干旱胁迫处理间细穗柽柳幼苗各性状指标的差异显著性;Microsoft Excel 2007分析平均数、标准差和百分比;采用SigmaPlot 12.5作图。
2 结果与分析
2.1 干旱胁迫对细穗柽柳幼苗株高和地径的影响
表1结果表明,干旱胁迫下,细穗柽柳幼苗的生长随干旱胁迫程度的增加而减缓。轻度干旱胁迫下(T1),细穗柽柳幼苗的株高和地径生长量均与CK无显著差异;随着胁迫增强、土壤水分含量的减少,T2和T3处理细穗柽柳幼苗的株高和地径生长量均显著低于CK,T2和T3两处理间无显著差异。
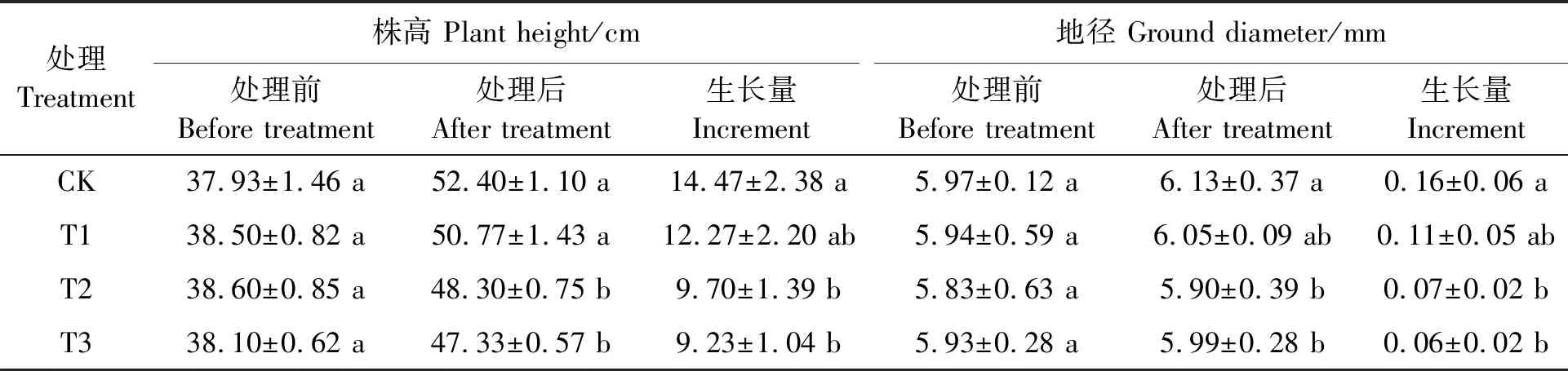
表1 干旱胁迫对细穗柽柳幼苗生长指标的影响Table 1 Effects of drought stress on growth indexes of Tamarix leptostachys Bunge seedlings
2.2 干旱胁迫对细穗柽柳幼苗叶片相对含水量的影响
图1显示,随着干旱胁迫强度增加,细穗柽柳幼苗叶片相对含水量呈持续下降趋势。与CK相比,各处理的叶片含水量均显著降低,且处理间也存在显著差异。随着干旱胁迫程度的增强,叶片含水量的降幅也逐渐增大,即与CK相比,轻度干旱胁迫(T1)导致叶片含水量降低6.35%;与轻度胁迫(T1)处理相比,中度干旱(T2)胁迫导致叶片含水量降低8.72%;与中度胁迫(T2)处理相比,重度干旱胁迫(T3)导致叶片含水量降低11.98%。
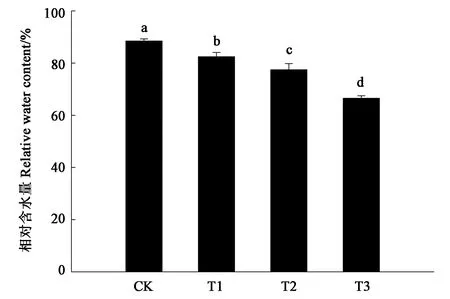
注:不同小写字母表示差异在P<0.05水平具有显著性。Note:Different small letters indicate significant difference at P<0.05 level.图1 干旱胁迫胁迫对细穗柽柳幼苗叶片相对含水率的影响Fig.1 Effect of drought stress on relative water content of Tamarix leptostachys Bunge seedling leaf
2.3 干旱胁迫对细穗柽柳幼苗叶片叶绿素含量的影响
从图2可以看出,随着干旱胁迫程度的增加,细穗柽柳幼苗叶片叶绿素含量呈下降趋势。叶绿素主要有叶绿素a和叶绿素b组成。干旱胁迫对叶绿素a含量的影响较大;轻度干旱胁迫(T1)下,细穗柽柳幼苗叶片的叶绿素a含量与CK无显著差异;随着胁迫增强、土壤水分含量的降低,T2和T3处理叶绿素a含量显著低于CK,而两处理间无显著差异。但干旱胁迫对叶绿素b的影响较小,各处理与CK均无显著差异。
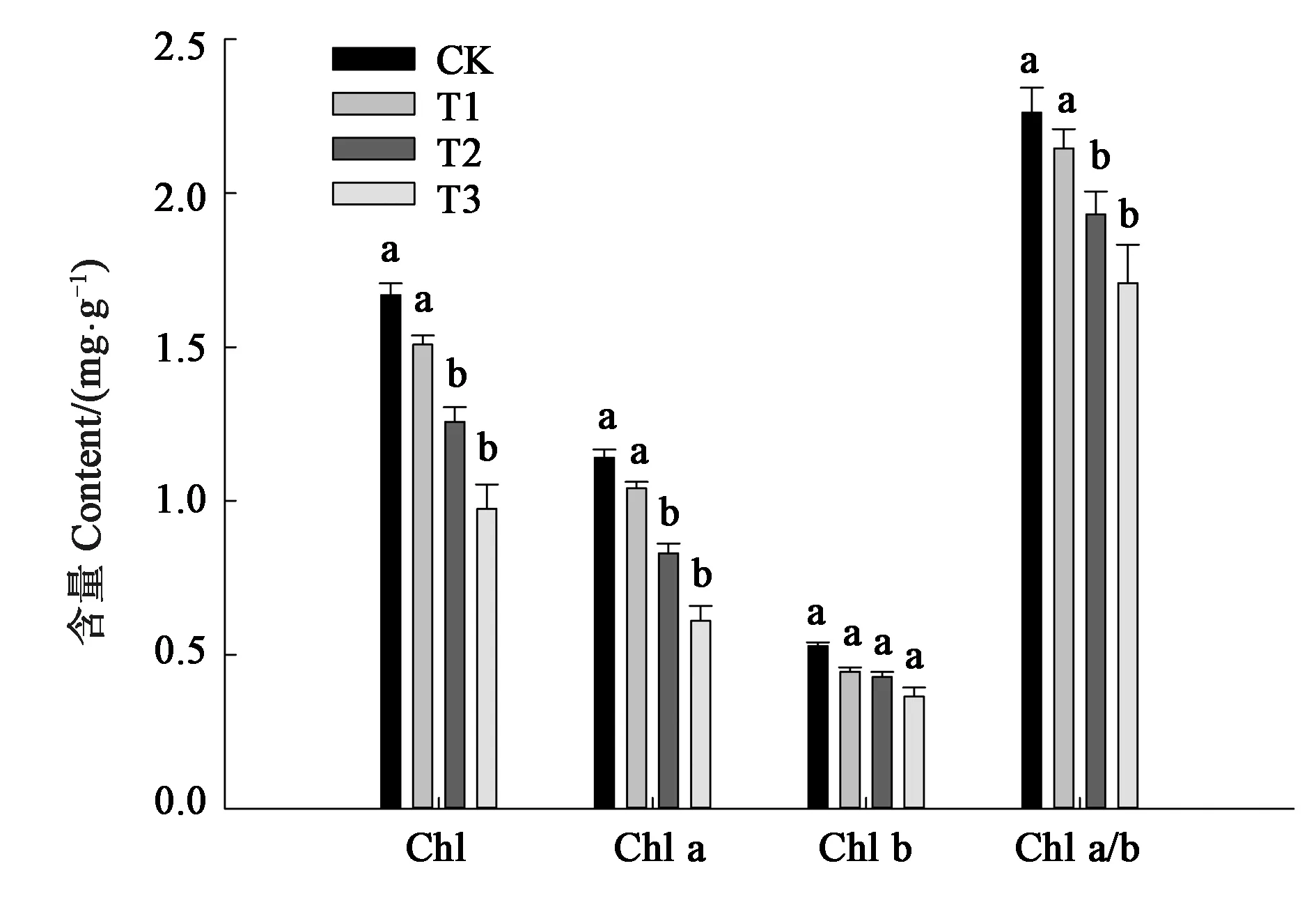
注:同一指标不同小写字母表示差异在P<0.05水平具有显著性。Note:Different small letters of the same index indicate significant difference at P<0.05 level.图2 干旱胁迫对细穗柽柳幼苗叶片叶绿素含量的影响Fig.2 Effect of drought stress on Chl content of Tamarix leptostachys Bunge seedling leaf
2.4 干旱胁迫对细穗柽柳幼苗叶片细胞膜透性的影响
图3显示,随着干旱胁迫强度的增加,细穗柽柳幼苗叶片的相对电导率和丙二醛含量呈逐渐上升的趋势,各处理均显著高于对照;且处理间也存在显著差异。其中,干旱胁迫下的叶片相对电导率是对照的1.5~3.0倍;丙二醛含量相比对照增加38.79%~62.85%。
2.5 干旱胁迫对细穗柽柳幼苗酶促保护系统的影响
POD是植物体内重要的抗氧化酶之一且活性较高,可以清除植物体内的H2O2。图4结果显示,随着水分亏缺程度的加剧,细穗柽柳幼苗POD活性呈上升趋势;干旱胁迫后叶片中POD的酶活显著高于CK。轻度干旱胁迫(T1)下,POD活性较CK增加55.88%,与CK差异不显著;随着胁迫程度的增强,POD活性开始急剧上升,中度(T2)、重度(T3)胁迫下POD的酶活比对照升高了179.41%和352.94%,与CK存在显著差异。SOD是防护氧自由基对细胞膜系统伤害的一种很重要的保护酶。随着干旱胁迫的加剧,SOD的活性呈直线上升趋势,各处理均与CK存在显著差异。其中,重度干旱胁迫(T3)下,叶片中SOD的酶活性与CK相比增加了132.15%。细穗柽柳幼苗CAT活性随干旱胁迫加剧呈先升高后降低的趋势,但各处理下的CAT酶活性均显著高于CK。轻度干旱胁迫(T1)下,细穗柽柳幼苗叶片中CAT酶活性是CK的1.51倍;中度干旱胁迫(T2)下,CAT活性快速升高,达到120.83 U·g-1·min-1,是CK的2.48倍;重度干旱胁迫(T3)下,CAT活性略有降低,为105.83 U·g-1·min-1,是CK的2.17倍。即适度的干旱使叶片中CAT酶活性上升,但过度的干旱胁迫又会影响CAT酶活性。
2.6 干旱胁迫对细穗柽柳幼苗渗透调节的影响
图5结果表明,干旱胁迫下细穗柽柳幼苗叶片Pro、可溶性糖和可溶性蛋白含量均呈上升趋势,各处理均与CK存在显著差异。其中,轻(T1)、中度(T2)干旱胁迫下,细穗柽柳幼苗叶片中Pro含量分别是CK的1.2倍和1.6倍,可溶性糖含量比CK增加20.35%和31.47%,可溶性蛋白含量比CK增加45.89%和74.35%。随着干旱胁迫加剧,重度干旱胁迫(T3)下,细穗柽柳幼苗叶片中Pro含量显著增加,浓度为304.98 μg·g-1,是CK的2.1倍;可溶性糖含量为22.33 mg·g-1,比CK增加48.36 %;可溶性蛋白含量比CK增加129.94%。
3 讨论
干旱胁迫下植物的形态特征能直接反映植物受到伤害的程度。株高、茎粗、叶片形态、叶片数、叶面积和生长速度等植物地上部的形态特征会因干旱胁迫造成不同的影响,通常植物会通过放慢生长进度或缩小叶片面积等来缓解水分亏缺造成的危害。本研究表明,轻度干旱对细穗柽柳幼苗的,株高和地径生长无显著影响;而随着干旱胁迫的加剧,株高和地径的生长速度明显受到抑制,与仇云峰等[9]对小叶杨、谢志玉等[10]对文冠果的研究结果相一致,表明细穗柽柳能够通过降低株高和地径的生长速度来减少水分的消耗从而适应干旱环境。
干旱胁迫下,相对含水量降低幅度较小表明叶片保水能力较强[11]。本研究结果表明,干旱胁迫下细穗柽柳幼苗叶片的相对含水率降低,且随着胁迫增强,叶片相对含水率的降幅逐渐增大。由此表明,轻、中度干旱胁迫下,细穗柽柳幼苗的持水能力较强,与前人对柽柳和侧柏的研究结果相一致[11-12]。
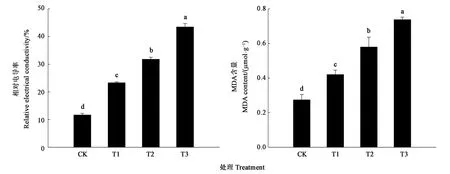
注:不同小写字母表示差异在P<0.05水平具有显著性。Note:Different small letters indicate significant difference at P<0.05 level.图3 干旱胁迫对细穗柽柳幼苗叶片相对电导率和MDA含量的影响Fig.3 Effect of drought stress on relative electrical conductivity and MDA content of Tamarix leptostachys Bunge seedling leaf
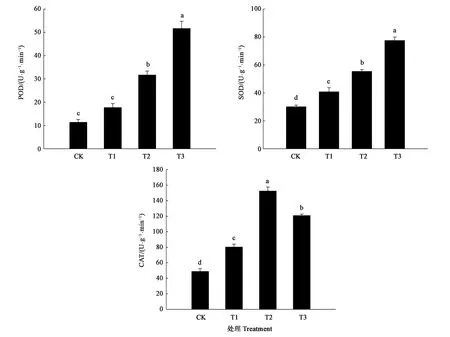
注:不同小写字母表示差异在P<0.05水平具有显著性。Note:Different small letters indicate significant difference at P<0.05 level.图4 干旱胁迫对细穗柽柳幼苗叶片POD、SOD和CAT活性的影响Fig.4 Effect of drought stress on POD, SOD and CAT activity of Tamarix leptostachys Bunge seedling leaf
叶绿素是植物光合作用中的基础色素[13-14],其含量在干旱胁迫环境下会降低[15]。本研究中,随着干旱胁迫程度的增加,叶绿素的含量显著降低,表明干旱胁迫影响了细穗柽柳幼苗叶片中叶绿素的生成。这与吴婧舒等[16]和刘遵春等[17]的研究结果一致。进一步分析表明,干旱胁迫主要降低了叶片中叶绿素a的含量,对叶绿素b含量无显著影响。
植物体内的抗氧化体系能缓解外界环境变化时对植物造成的伤害,由SOD、CAT和POD等抗氧化酶组成[18]。SOD、POD和CAT都是保护酶,可以清除活性氧以保持胞内的平衡状态,且保护酶之间的协作也可以保证膜系统不受侵害[19-20]。本研究表明,随着干旱胁迫的增强,细穗柽柳的SOD活性显著增强;POD活性在轻度干旱胁迫时变化不明显,但当胁迫增强时活性显著提高;CAT活性在轻度、中度胁迫下显著升高,而重度胁迫下活性略有降低。由此表明,细穗柽柳幼苗受到水分胁迫后能够启动SOD、POD和CAT等保护酶,提高酶活性来缓解干旱造成的伤害。在胁迫较重时,POD起关键作用;CAT在土壤干旱胁迫较轻时发挥作用;中间状态时,由两者共同起作用,表明细穗柽柳有较强的抗旱能力。
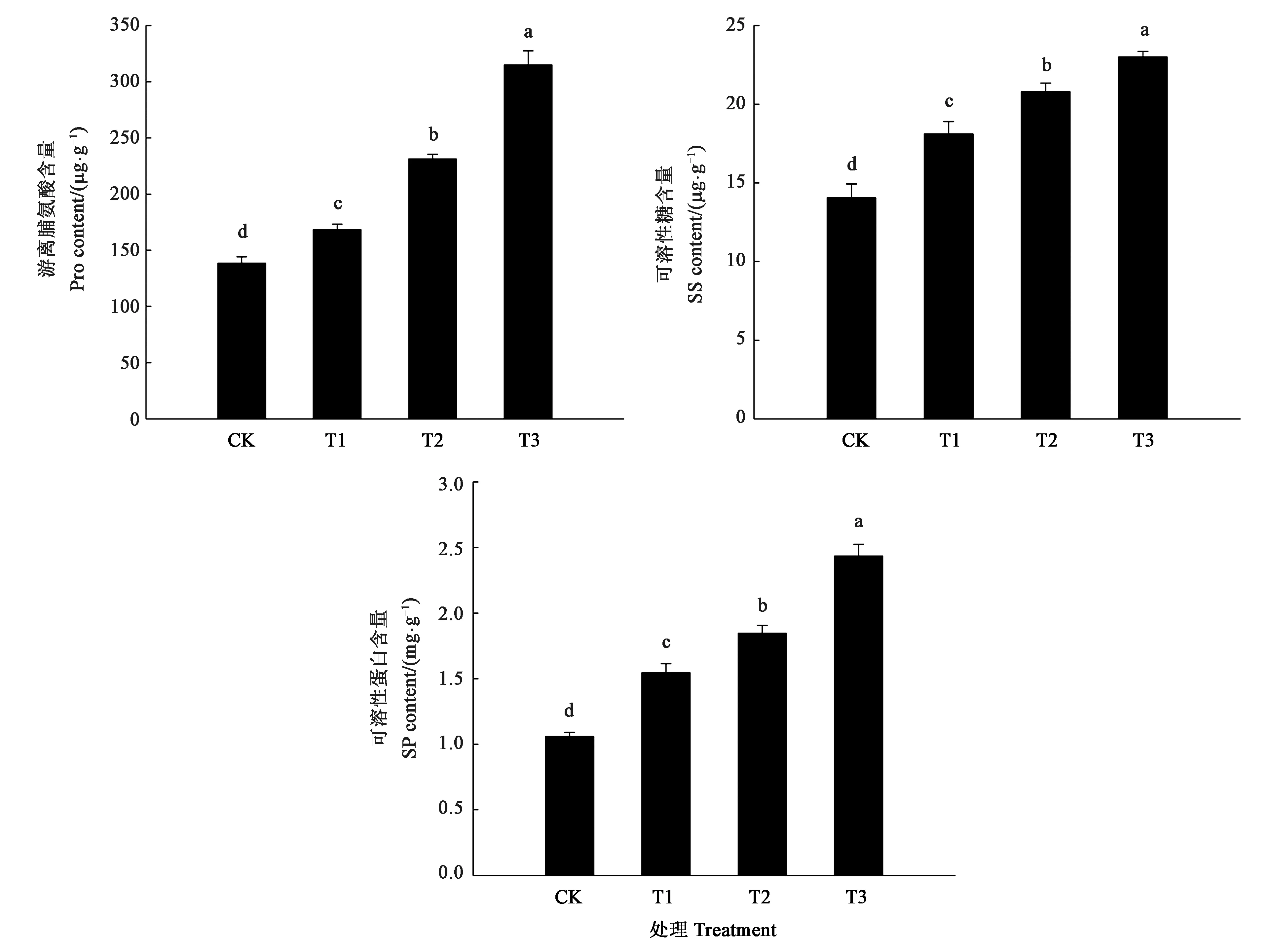
注:不同小写字母表示差异在P<0.05水平具有显著性。Note:Different small letters indicate significant difference at P<0.05 level.图5 干旱胁迫对细穗柽柳幼苗叶片Pro、SS和SP含量的影响Fig.5 Effect of drought stress on Pro, SS and SP content of Tamarix leptostachys Bunge seedling leaf
膜脂过氧化的重要产物之一是丙二醛(MDA)[21],当植物遭受环境胁迫时,内部平衡被打破,引发脂质过氧化作用,产生大量的MDA[22]。因此,MDA在许多研究中被用作判断植物抗旱能力的一个重要指标[23]。本研究中,随着干旱胁迫的增强,细穗柽柳幼苗叶片中MDA含量增多,细胞膜相对透性变大;轻度干旱胁迫时,细胞膜相对透性和MDA含量与对照无显著差异,表明细穗柽柳的抗旱性较强,轻度干旱未对细胞内环境造成严重伤害。在重度胁迫下,细穗柽柳的细胞膜相对透性和MDA大幅增长,说明此时膜脂过氧化作用变强,膜系统受到损伤。这与陈志成等[24]和曾艳等[25]的研究结果类似。
渗透调节是植物在逆境下降低渗透势,抵抗逆境胁迫的一种重要方式[26]。叶片游离脯氨酸、可溶性糖和可溶性蛋白含量的增加可以调节渗透势和保持平衡,提高植物的渗透调节能力,保持在干旱胁迫下植物生长所需要的水分,增强气孔的调节和持水能力,提升植物对干旱环境的适应能力[27]。本研究中,随着干旱胁迫程度的增强,细穗柽柳幼苗叶片游离脯氨酸、可溶性糖和可溶性蛋白含量均显著增加。由此表明,细穗柽柳幼苗在干旱胁迫时,通过提高叶片游离脯氨酸、可溶性糖和可溶性蛋白含量来增强对逆境环境的适应性。其中, 游离脯氨酸和可溶性蛋白含量的增幅较高,这可能是由于少量的糖转变为游离脯氨酸。在植物正常生长环境下,游离脯氨酸常常会转化为其他物质;而在干旱胁迫环境下,游离脯氨酸的转化效率降低,使其快速累积,参加蛋白质组成[28]。故在干旱胁迫下,细穗柽柳幼苗叶片中可溶性蛋白含量明显增加,这与胁迫加强时游离脯氨酸大量累积来维持细胞膨压和缓解干旱胁迫有着不可分割的联系[29]。
综上所述,细穗柽柳幼苗在干旱胁迫时能通过调整自身生长速度、保护酶活性及渗透调节物质含量来缓解干旱胁迫造成的伤害,即使在重度胁迫下,也未出现叶片凋落及幼苗死亡的现象。由此表明,细穗柽柳具有较强的抗旱性和适应性,在干旱、半干旱地区具有极高的推广价值。
猜你喜欢
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
恋爱婚姻家庭·养生版(2021年10期)2021-10-28
少儿科学周刊·少年版(2021年20期)2021-01-17
阅读(科学探秘)(2020年8期)2020-11-06
农家科技(2020年3期)2020-05-11
绿色科技(2019年2期)2019-05-21
女性天地(2016年10期)2017-04-25
哈尔滨理工大学学报(2016年2期)2016-09-12
农家顾问(2016年7期)2016-05-14
