
龙眼果实脱落特性参数与果柄分离力的相关性分析
2021-12-08 06:40杨子琴,李建国,张蕾,李松刚,洪继旺,黄旭明
热带作物学报 2021年10期
杨子琴,李建国,张蕾,李松刚,洪继旺,黄旭明

摘 要:海南反季节龙眼采前落果普遍发生,严重制约了反季节龙眼产业的发展。而果柄分离力可以反映果实离层活动的状况,成为果实脱落进程的重要指标,但未见在龙眼上使用的报道。本研究以DS2-1000 gf和DS2-5000 gf型推拉力仪检测果柄分离力,并分析果實脱落过程中各项生理特性参数,旨在弄清龙眼果实脱落过程中果柄分离力与果实碳水化合物、呼吸速率和离层细胞壁代谢酶活性的关系。数据表明反季节龙眼果实脱落过程中,果柄分离力的变化范围为0~3000 gf,且果柄分离力低于1000 gf会导致果实呈现脱落趋势;落果率与果柄分离力呈现显著负相关性(r=–0.984,P=0.000);果实总糖和淀粉含量与果柄分离力呈现显著正相关性,相关系数分别为0.942(P=0.005)和0.952(P=0.003);果柄呼吸耗氧速率与果柄分离力呈负相关性(r=–0.807,P=0.099);果柄离层纤维素酶和β-甘露聚糖酶活性也与果柄分离力呈显著分负相关性,相关系数分别为–0.936(P=0.019)和–0.954(P=0.002)。结果表明反季节龙眼果实脱落进程可用果柄分离力的变化体现,果实脱落进程伴随着果柄分离力的不断降低,与果柄细胞壁降解酶活性的增加呈线性关系;脱落的发生与碳水化合物含量关系密切,而高的果实呼吸消耗,可能加快果实脱落进程。
关键词:龙眼;果柄分离力;果实;脱落
中图分类号:S667.1 文献标识码:A
Relativity Analysis Between Characteristic Parameters of Off-season Longan Abscission and Its Fruit Removal Force
YANG Ziqin1, LI Jianguo2, ZHANG Lei1, LI Songgang1, HONG Jiwang1, HUANG Xuming2*
1. Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences / National Center for Crops Varieties Improvement, Haikou, Hainan 571101, China; 2. College of Horticulture, South China Agricultural University / State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources, Guangzhou, Guangdong 510642, China
Abstract: Pre-harvest fruit abscission phenomenon is intensive in off-season longan in Hainan, China, which seriously restricts the development of the off-season longan. Pedicel detaching force or fruit removal force, reflecting the separation status of the abscission zone, has been adopted as a direct index reflecting the process of fruit abscission. However, there has been no report about its use in longan. The purpose of our study was to evaluate the use of pedicel detaching force to reflect fruit abscission in off-season longan and clarify its correlation with availability of carbohydrates, fruit respiration rate and activities of wall-degrading enzymes in the abscission zone. Small clusters of fruit were harvested and pedicel detaching force was measured using DS2-1000 gf or DS2-5000 gf force gauges every day. At the same time, correlation of various physiological parameters with pedicel detaching force was analyzed. The pedicel detaching force of the off-season longan of ‘Chuliang ranged between 0 and 3000 gf. When it became less than 1000 gf, the fruit was doomed to abscise. The cumulative abscission rate was significantly and negatively correlated to pedicel detaching force, with a coefficient of –0.948 (P=0.000). Total sugar and starch content in fruit was significantly and positive correlated to pedicel detaching force with a correlation coefficient of 0.942 (P=0.005) and 0.952 (P=0.003), respectively. A moderate positive bud insignificant correlation was found between respiratory oxygen consumption and pedicel detaching force (r=–0.807, P=0.099). Activity of cellulase and β-mannanase in the abscission zone was negatively correlated to pedicel detaching force, with a correlation coefficient of ?0.936 (P=0.019) and –0.954 (P=0.002), respectively. The results showed changes in pedicel detaching force could be used to reflect the process of fruit abscission. Fruit abscission was accompanied by significant decrease in pedicel detaching force, which was highly correlated with carbohydrate availability and cell wall degrading enzyme activity in the abscission zone, and high fruit respiration consumption may accelerate the process of fruit abscission.
Keywords: longan; fruit removal force; fruits; abscission
DOI: 10.3969/j.issn.1000-2561.2021.10.032
龍眼(Dimocarpus longan Loureiro)是我国南方主栽热带亚热带果树之一,正常情况下,其坐果力强,素有“爱果不惜树”之印象[1]。海南以生产反季节龙眼为主,但是反季节龙眼采前落果现象普遍发生,严重制约了产业的发展。如何促进反季节龙眼保果增产是亟待解决的产业难题。前人的研究表明龙眼果实的脱落除了和授粉受精不良有关外,主要由营养供应不足及营养竞争导致[2]。海南反季节龙眼夏秋季节花、果、梢同时生长对营养的竞争极大,恰逢高温、高湿、多雨天气,光合产物积累不足;秋冬季果实发育期营养供给后力不足。因此针对光合产物供应不足条件下龙眼果实脱落调控机理的研究具有重要的理论意义,对海南当地龙眼产业来说,有地域特殊性。
果实是一个大量消耗养分的异养生殖器官。在果实发育过程中,落果是果树减少果实负载量,集中树体资源保证留树果实个体发育的正常现象。这种现象被视为果树果实负载量的自我调节机制。果柄分离力大小是决定果实是否脱落的重要衡量指标。为了更好的了解果柄分离力及果实脱落相关参数的关系,前人在研究果实早期脱落的同时对果实生理生化变化与其果柄分离力之间的关系进行了研究[3-7]。Ferrara等[8]利用乙烯利处理刺激成熟葡萄浆果的脱落,降低了果实分离力,并促进了柄端疤痕的形成,进一步促进高品质无柄鲜食葡萄的生产。用茎腐病Diplodia侵染柑橘加剧采前落果,PCR鉴定Diplodia CT值与果实脱离力呈正相关(R=0.855)[9]。乙烯利和CMNP(水果特异性脱落剂)诱导柑橘果实分离力降低,进而诱发果实早期脱落[10]。
目前对龙眼果实脱落过程中生理特性参数相关研究较多,还没有龙眼果实脱落过程中果柄分离力变化的报道。龙眼果实脱落程度还没有判定标准,至今只还停留在“落或不落”层面。结合脱落过程中果实落果率、总糖和淀粉含量、果柄呼吸耗氧量、果柄离层纤维素酶和β-甘露聚糖酶活性等指标,研究龙眼果实脱落各项生理特性参数变化,分析果实脱落相关的生理指标与果柄分离力之间的关系可有效的确立果柄分离力对果实脱落程度的诠释。
反季节龙眼果实发育过程中极易发生落果,果柄分离力的大小决定着果实附着于树体的寿命长短,分析龙眼各项生理特性与果柄分离力的相关性,不仅为研究落果与果柄分离力的关系,也为研究反季节龙眼科学保果提供参考。
1 材料与方法
1.1 材料
试验于2020年6月,在中国热带农业科学院热带作物品种资源研究所国家热带果树品种改良中心龙眼种质圃内进行。反季节龙眼为5年生‘储良龙眼,3月初株施0.4 kg氯酸钾催花处理,于4月底盛花。盛花后50 d,选取树势一致、坐果量相近、果实大小一致的4株龙眼树做试验用树。田间常规肥水管理。
1.2 方法
1.2.1 样品处理 受光照、养分等因素的影响,果树长势会有差异,尤其内膛果长势弱。因此取样时选择外周中上部坐果量相近的果穗进行。每株选取5~8 mm直径的30个结果母枝,在距果穗基部约10 cm处环剥,宽度2 mm,深达木质部,同时摘除环剥口以上的叶片,中断果实碳水化合物供应,人为制造果实饥饿胁迫。处理后每日跟踪调查果实脱落情况,同时采集果穗做检测。为了方便测量果柄分离力,样品果实随果穗一起剪下,冰盒带回实验室检测。
1.2.2 果柄分离力 使用智取DS2-1000 gf、DS2-5000 gf推拉力试验机。
测试前将上端夹具卸除,将底座夹具螺旋固定在原上端夹具位置。果实连同一段果枝夹在上端固定装置上,按下ZERO和PEAK键,手动向下拉果柄,记录在离层部位断裂的最大拉力值。每处理果柄分离力取5次测量的平均值。
所有测量果柄分离力手动拉力方向均为竖直向下,因此要求果柄与夹具在同一垂直线。
1.2.3 果实落果率 以处理当天为初始值(落果率0%)计算落果率动态,直至处理果穗果实全部脱落。每株选10个处理(环剥+去叶)的果穗分别挂牌编号记录。
1.2.4 总糖和淀粉含量 蒽酮法测定总糖含量[11];KI-I比色法测定淀粉含量[12]。
1.2.5 果柄呼吸耗氧量 采用氧电极(Hansatech Oxygraph)测定,反应体系2 mL,反应底物为溶解有饱和氧的超纯水,零氧线建立用高纯氮。反应温度25 ℃,反应时间取斜率稳定的一段计数,设3次重复,计算果柄呼吸耗氧量。
1.2.6 果柄离层纤维素酶和β-甘露聚糖酶活性 果柄离层纤维素酶和β-甘露聚糖酶活性采用琼脂糖凝胶扩散法[13-14],设6次重复。
1.3 数据处理
果柄分离力与龙眼各项生理参数测试数据采用SPSS 21.0统计分析软件进行分析。根据方差分析结果,对不同龙眼品种的果柄分离力相对于果实落果率、总糖和淀粉含量、果柄呼吸耗氧量、果柄离层纤维素酶和β-甘露聚糖酶活性进行回归分析,并建立回归模型,分析回归模型的相关系数,确定回归模型的可靠性。
2 结果与分析
2.1 龙眼果实各项参数的测定结果
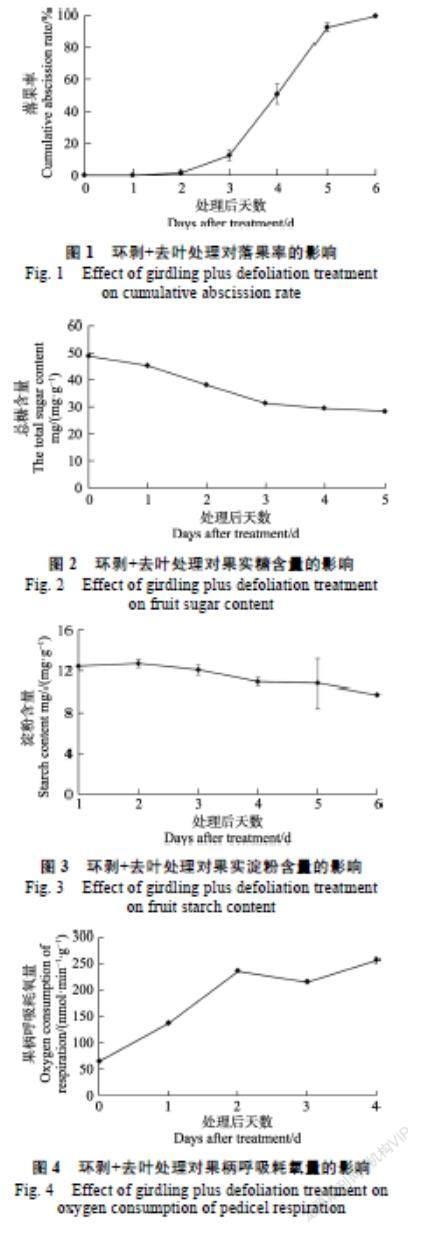
龙眼果实各项参数连续测量结果曲线如图1所示,脱落过程持续6 d,总糖和淀粉含量28.34~ 48.84 mg/g、9.80~12.76 mg/g;果柄呼吸耗氧量58.90~258.94 nmol/(ming)、果柄离层纤维素酶相对活性0~1.84(活性值取对数)和β-甘露聚糖酶活性0.47~2.48(活性值取对数)。从图1可以看出,3 d开始启动大量落果,4 d落果率激增至51.10%。随着果实脱落进程的推进,果实总糖含量不断下降(图2),淀粉含量小幅下降(图3),果柄呼吸耗氧量持续上升(图4),果柄离层纤维素酶和β-甘露聚糖酶相对活性持续上升(图5、图6),果柄分离力不断下降(图7)。
2.2 龙眼各项参数对果柄分离力的影响
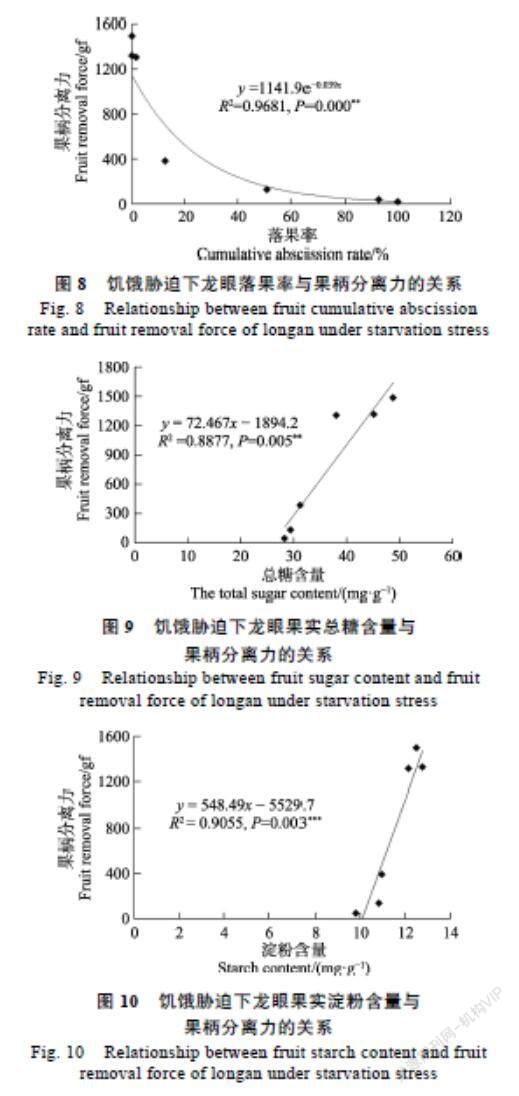
2.2.1 落果率与果柄分离力的关系 落果率越高,树体产量越低,直接影响了经济效益,因此保果对反季节龙眼生产尤为重要。随着落果率的上升,果柄分离力逐渐下降。通过每日监测果实落果动态计算落果率,相关性检验在0.01显著水平下果柄分离力与落果率显著相关。为确定显著性相关规律,对果柄分离力进行曲线回归分析表明:试验中龙眼果柄分离力与落果率多项式曲线回归方程为:y=1141.9e?0.039x;R?=0.9681,拟合度很高。从图8可以看出:随着果柄分离力不断降低,落果率逐渐上升。
2.2.2 果实糖含量与果柄分离力的关系 果实是一个库力强大的异养器官,主要依赖外源营养供给[15]。光合产物的供求平衡在坐果/脱落平衡中扮演重要的调节作用[16]。果实糖含量是衡量果实光合产物积累量的重要指标。前期的研究表明对正造龙眼结果母枝环割+去叶处理引发果实大量脱落,且果实糖含量可能存在一个阈值,低于该阈值,便触发果实的脱落[14],即碳水化合物的缺乏是导致龙眼更快速脱落的关键诱因。果柄分离力的检测是将脱落过程这一抽象的进程进行量化研究,更科学的判断脱落的发展阶段。通过对处理后果实糖含量变化检测,相关性检验在0.01显著水平下果柄分离力与果实糖含量显著相关。为确定显著性相关规律,对果柄分离力进行曲线回归分析表明:试验中龙眼果柄分离力与果实糖含量多项式曲线回归方程为:y=72.467x?1894.2;R2=0.8877,拟合度较高。从图9可以看出,随着果实糖含量的下降,果柄分离力不断降低。
2.2.3 果实淀粉含量与果柄分离力的关系 果实淀粉含量是果实碳水化合物积累量的另一的重要形式。晚熟脐橙正常果中葡萄糖、蔗糖和淀粉含量均显著高于脱落果,碳水化合物向果实的富集,对减轻第一次和第二次生理落果具有重要影响[17]。通过对处理后果实淀粉含量变化检测,相关性检验在0.01显著水平下果柄分离力与果实淀粉含量显著相关。曲线回归分析表明:试验中龙眼果柄分离力与果实淀粉含量多项式曲线回归方程为:y=548.49x?5529.7;R2=0.9055,拟合度较高。从图10可以看出:随着果实淀粉含量的下降,果柄分离力不断降低。
2.2.4 果柄呼吸耗氧量與果柄分离力的关系 呼吸耗氧量在一定程度上反应了果柄所处的状态,呼吸耗氧量越高,则对碳水化合物消耗越快,进而加速果实脱落进程。通过对每日果柄呼吸耗氧量的检测,相关性检验在0.01显著水平下,果柄分离力与果柄呼吸耗氧量显著相关。曲线回归方程为:y = ?6.270 2x + 2079;R2= 0.650 9,拟合度较高。从图11可以看出:随着果柄呼吸耗氧量的上升,果柄分离力不断降低。
2.2.5 离层纤维素酶相对活性与果柄分离力的关系 纤维素酶在植物器官脱落中起重要作用[18]。通过对处理后果柄离层纤维素酶相对活性变化检测,相关性检验在0.01显著水平下果柄分离力与果柄离层纤维素酶相对活性显著相关。曲线回归分析表明:试验中龙眼果柄分离力与果柄离层纤维素酶活性多项式曲线回归方程为:y=?787.8x+ 1425.1;R2=0.8767,拟合度高。从图12可以看出:随着果柄离层纤维素酶活性的上升,果柄分离力不断降低。
2.2.6 β-甘露聚糖酶相对活性与果柄分离力的关系 通过对处理后果柄离层β-甘露聚糖酶相对活性变化检测,相关性检验在0.01显著水平下果柄分离力与果柄离层β-甘露聚糖酶活性显著相关。曲线回归分析表明:试验中龙眼果柄分离力与果柄离层β-甘露聚糖酶活性多项式曲线回归方程为:y=?736.6x+1815;R2=0.9108,拟合度极高。从图13可以看出:随着果柄离层β-甘露聚糖酶活性的上升,果柄分离力不断降低。
3 讨论
龙眼的早期落果严重制约了反季节龙眼产业的发展,保果技术的研发成为目前最迫切的技术需求。龙眼果实脱落伴随着一系列的特性参数的变化,对其变化与果柄分离力变化进行相关性分析,对保果技术的研究具有重要的指导意义。果树树体碳水化合物营养储备不足,常出现果实发育障碍,落果现象十分严重[19]。荔枝在碳水化合物供应不足的情况下也发生类似情况的大量落果,如:阴雨连绵、光照不足和光合叶片数量不足[20-21]。树干注射蔗糖溶液明显促进柑橘幼果中蔗糖、淀粉含量的积累,有利于坐果[22]。温州蜜柑和日本甜夏橙的淀粉含量与幼果相对脱落率均呈负相关[23]。本研究也有相同结论,龙眼果实糖和淀粉含量与果柄分离力正相关,与落果率呈负相关。
植物呼吸作用为生物合成提供能量,其与光合作用的平衡决定了植物生物量的积累率[24]。阿月混子的花芽在脱落前会出现呼吸上升现象[25];佘小平等[26]报道GA诱导大叶黄杨离体茎段的叶柄脱落,同时可诱导呼吸显著提高。本研究中伴随着龙眼果实脱落,果柄呼吸耗氧量也出现上升现象,且与果柄分离力呈负相关。
大量研究表明纤维素酶(EG)在组织脱落中具有重要作用[27-29]。本研究证实纤维素酶在龙眼果实脱落过程中活性上升。
β-甘露聚糖酶是降解植物细胞壁半纤维素的主要酶类,前人的研究表明该酶在植物种子萌发、果实成熟和花粉发育等过程中有作用,但对于该酶在器官脱落中的作用还存在争议。任艳芳等[30]在脐橙离区克隆了β-甘露聚糖酶基因片段,进一步肯定了β-甘露聚糖酶在植物脱落器官离区中的存在。Belfield等[31]认为β-甘露聚糖酶虽在脱落中活性水平和基因水平上有一定表达,但它不是引起脱落的关键酶。在大豆叶柄脱落的研究中发现内切-β-甘露聚糖酶GmMAN1与叶脱落无关,但可能参与了对伤害的反应[32]。本研究发现了果实脱落过程中β-甘露聚糖酶活性上升。
目前还没有通过果柄分离力来判断龙眼果实脱落程度的报道,通过直接观测的方法具有很大的局限性,因此采用推拉力试验机测量果柄分离力的变化范围,能够更精确地判断果实脱落的程度。将果实脱落这一抽象参数进行量化的研究,更加科学地判断果实是否有脱落的趋向,进而为新型保果技术的研发提供参考。
4 结论
龙眼果柄分离力是直观反映果实分离动态的一个重要参数。本研究结果表明:果实脱落相关的特性参数和果柄分离力之间有着很高的相关性,其中落果率与果柄分离力相关系数为?0.984;果实总糖和淀粉含量与果柄分离力相关系数分别为0.942和0.952;果柄呼吸耗氧量与果柄分离力相关系数为?0.807;果柄离层纤维素酶和β-甘露聚糖酶活性与果柄分离力相关系数分别为?0.936、?0.954;通过对果柄分离力与生理特性参数进行曲线拟合分析,得到拟合方程和曲线。果实脱落过程龙眼果柄分离力的变化范围为0~3000 gf,且果柄分离力低于1000 gf会导致果实呈现脱落趋势,低于400 gf时果实极易脱落。
参考文献
[1] 杨子琴, 李 茂, 章笑赟, 等. 饥饿胁迫对龙眼果实脱落及糖代谢的影响[J]. 果树学报, 2011, 28(3): 428-432.
[2] 韩冬梅, 郭栋梁, 潘学文, 等. 不同品種龙眼果实发育进程对其生理落果和熟性的影响[J]. 广东农业科学, 2011, 38(7): 59-62.
[3] Rizzuti A, Aguilera-saez L M, Gallo V, et al. On the use of ethephon as abscising agent in cv. crimson seedless table grape production: combination of fruit detachment force, fruit drop and metabolomics[J]. Food Chemistry, 2015, 171(15): 341-350.
[4] Goldental-cohen S, Burstein C, Biton I, et al. Ethephon induced oxidative stress in the olive leaf abscission zone enables development of a selective abscission compound[J]. BMC Plant Biology, 2017, 17(1): 87.
[5] Maule A, Henning G, Patterson S. Love me not meter: A sensor device for detecting petal detachment forces in Arabidopsis thaliana[J]. Methods in Molecular Biology, 2017(1573): 245-252.
[6] Garcia-rojas M, Meneses M, Oviedo K, et al. Exogenous gibberellic acid application induces the overexpression of key genes for pedicel lignification and an increase in berry drop in table grape[J]. Plant Physiology and Biochemistry, 2018(126): 32-38.
[7] Lanza F E, Marti W, Silva G J, et al. Characteristics of citrus canker lesions associated with premature drop of sweet orange fruit[J]. Phytopathology, 2019, 109(1): 44-51.
[8] Ferrara G, Mazzeo A, Matarrese A M, et al. Ethephon as a potential abscission agent for table grapes: effects on pre-harvest abscission, fruit quality, and residue[J]. Frontiers in Plant Science, 2016(7): 620.
[9] Zhao W, Bai J H, Mccollum G, et al. High incidence of preharvest colonization of huanglongbing-symptomatic citrus sinensis fruit by Lasiodiplodia theobromae (Diplodia natalensis) and exacerbation of postharvest fruit decay by that fungus[J]. Applied and Environmental Microbiology, 2015, 81(1): 364-372.
[10] Malladi A, Burns J K. CsPLDα1 and CsPLDγ1 are differentially induced during leaf and fruit abscission and diurnally regulated in Citrus sinensis[J]. Journal of Experimental Botany, 2008, 59(13): 3729-3739.
[11] 宁正祥. 食品成分分析手册[M]. 北京: 中国轻工业出版社, 1998: 26-27.
[12] 徐昌杰, 陈文峻, 陈昆松, 等. 淀粉含量测定的一种简便方法——碘显色法[J]. 生物技术, 1998, 8(2): 41-43.
[13] 杨子琴, 李建国, 王惠聪, 等. 一种测定龙眼果柄离层纤维素酶活性的方法[J]. 华南农业大学学报, 2012, 33(2): 175-177.
[14] 杨子琴. 饥饿胁迫诱导龙眼果实脱落的信号发生与调控机理研究[D]. 广州: 华南农业大学, 2011.
[15] Hieke S, Menzel C M, Lüdders P. Effects of leaf, shoot and fruit development on photosynthesis of lychee trees (Litchi chinensis)[J]. Tree Physiology, 2002, 22(13): 955-961.
[16] Lakso A N, Greene D W, Palmer J W. Improvements on an apple carbon balance model[J]. Acta Hortic, 2006(707): 57-61.
[17] 姚珍珍. 晚熟脐橙落花落果生态影响因子及生理机制研究[D]. 重庆: 西南大学, 2012.
[18] Li C Q, Zhao M L, Ma X S, et al. Two cellulases involved in litchi fruit abscission are directly activated by an HD-Zip transcription factor LcHB2[J]. Journal of Experimental Botany, 2019.
[19] Yang W H, Zhu X C, Bu J H, et al. Effects of bagging on fruit development and quality in cross-winter off-season longan[J]. Scientia Horticulturae, 2009, 120(2): 194-200.
[20] Yuan R C, Huang H B. Litchi fruit abscission: its patterns, effect of shading and relation to endogenous abscisic acid[J]. Scientia Horticulturae, 1988, 36: 281-292.
[21] 黃永敬, 吴 文, 曾继吾, 等. 夏梢生长条件下树干供糖对‘砂糖橘幼果糖代谢及脱落的影响[J]. 热带作物学报, 2019, 40(8): 1522-1528.
[22] 徐昌杰, 张上隆. 柑橘幼果发育期碳水化合物代谢及其与生长发育的关系[J]. 果树学报, 2001, 18(1): 20-23.
[23] Millar A H, Whelan J, Soole K L, et al. Organization and regulation of mitochondrial respiration in plants[J]. Annual Review of Plant Biology, 2011, 62(1): 79-104.
[24] Vemmos S N. Pontikis C A, Tolza-marioli A P. Respiration rate and ethylene production in inflorescence buds of pistachio in relation to alternate bearing[J]. Scientia Horticulturae, 1994, 57: 165-172.
[25] Kalaitzis P, Hong S B, Solomos T, et al. Molecular characterization of a tomato endo-β-1,4-glucanase gene expressed in mature pistils, abscission zones and fruit[J]. Plant and Cell Physiology, 1999, 40(8): 905-908.
[26] 佘小平, 黄维玉. GA3处理后离区组织呼吸途径的改变与脱落的关系[J]. 西北植物学报, 1989, 9(1): 20-25.
[27] Libertini E, Li Y, Mcqueen-mason S J. Phylogenetic analysis of the plant endo-β-1,4-glucanase gene family[J]. Journal of Molecular Evolution, 2004, 58(5): 506-515.
[28] Trainotti L, Pavanello A, Zanin D. PpEG4 is a peach endo-β-1,4-glucanase gene whose expression in climacteric peaches does not follow a climacteric pattern[J]. Journal of Experimental Botany, 2006, 57(3): 589-598.
[29] Flors V, Leyva mde L, Vicedo B, et al. Absence of the endo-β-1,4-glucanases Cel1 and Cel2 reduces susceptibility to Botrytis cinerea in tomato[J]. The Plant Journal, 2008, 52(6): 1027-1040.
[30] 任艳芳, 刘厚宇, 何俊瑜. 脐橙离区β-甘露聚糖酶基因片段克隆和序列分析[J]. 中国农学通报, 2011, 27(2): 119- 122.
[31] Belfield E J, Ruperti B, Roberts J A, et al. Changes in expansin activity and gene expression during ethylene-promoted leaflet abscission in Sambucus nigra[J]. Journal of Experimental Botany, 2005, 56(413): 817-823.
[32] Yan M, Zhang Y F, Guo W J, et al. Soybean endo-β-mannanase GmMAN1 is not associated with leaf abscission, but might be involved in the response to wounding[J]. PLoS One, 2012, 7(11): e49197.
责任编辑:白 净
猜你喜欢
作文大王·低年级(2019年4期)2019-05-13
世界热带农业信息(2018年5期)2018-11-09
世界热带农业信息(2018年6期)2018-03-05
世界热带农业信息(2018年7期)2018-01-19
江苏农业科学(2016年8期)2017-02-15
天津农业科学(2016年12期)2017-01-11
学苑创造·A版(2016年9期)2016-10-10
山西果树(2015年1期)2015-03-31
安徽农学通报(2014年3期)2014-03-11
雕塑(1996年1期)1996-07-12
