
基于脂质组学技术探究热处理和发酵对乳脂质的影响
2022-08-02 03:10高文浩尹乾隆王筱迪滕翔宇任皓威
食品科学 2022年14期
高文浩,尹乾隆,王筱迪,滕翔宇,2,任皓威,2,刘 宁,2,*
(1.东北农业大学食品学院,乳品科学教育部重点实验室,黑龙江 哈尔滨 150030;2.哈尔滨腾凝科技有限公司,黑龙江 哈尔滨 150030)
酸乳是世界上最受欢迎的发酵乳制品之一,作为健康食品已获得消费者的广泛认可。酸乳不仅能提供全面营养,还具有许多健康益处,其生产是一个长期的、不断变化的过程。酸乳的质量特性受诸多因素影响,其中热处理和发酵是最重要两方面。在20世纪后期,酸乳实现了生产工业和生产过程标准化,其中热处理一般采用高温巴氏杀菌,要求温度在85 ℃持续20~30 min或90~95 ℃持续5 min,在此条件下除孢子外,大多数微生物被杀死,大多数酶失活,从而确保了产品的安全性。在商业化酸乳制作中最常用的发酵剂是嗜热链球菌和德氏乳杆菌保加利亚亚种,在42 ℃左右条件下发酵,这一阶段诱导凝乳的形成,促使酸乳产生质地特征和独特风味。然而,目前关于酸乳加工过程中热处理和发酵会对产品脂质影响的研究较少。
脂质组学是组学的重要分支,作为食品科学领域中高效、敏感和强大的分析工具应用范围十分广泛,如食品加工及转化和食物对人类健康影响等研究。然而,以酸乳为中心的脂质组学研究有限。章丽等基于脂质组学技术鉴定了嗜热链球菌发酵乳的脂质构成;Lordan等基于超高效液相色谱(ultra-high performance liquid chromatography,UPLC)结合高分辨率质谱分析发酵对羊乳极性脂质的影响,并探究极性脂质对血小板活化因子和凝血酶的抑制作用;Furse等用脂质组学技术分析市售牛乳、酸乳和乳酪的脂质特性,但并未深入研究发酵对脂质的影响。因此,本研究基于非靶向脂质组学技术系统阐明牛酸乳和羊酸乳制造过程中脂质组成及动态变化,深入探究热处理-巴氏杀菌(物理因素)和发酵-菌种脂质代谢(生物化学因素)对牛乳和羊乳脂质的影响,并筛选出可作为鉴别产品是否经热处理和发酵的潜在标志物,用于商业化生产酸乳过程中加工工艺终点及产品状态的判断,并在分子水平上探索热处理和发酵对不同乳基产生的共同脂质的变化,以期推动对酸乳最终产品质量和营养特性的理解。
1 材料与方法
1.1 材料与试剂
全脂牛乳和全脂羊乳分别购自中国黑龙江省哈尔滨市当地牧场,品种分别为荷斯坦乳牛和萨能山羊,饲喂日粮均为玉米、黄豆和青贮饲料。采集10 头乳牛和乳山羊挤出的新鲜牛乳和羊乳,分别混合后立即转移至实验室进行酸乳制作。
发酵剂MY105(嗜热链球菌和德氏乳杆菌保加利亚亚种按1∶1混合) 法国丹尼斯克公司;乙腈、异丙醇、甲醇 美国Thermo Fisher Scientific公司;其余试剂均为色谱级。
1.2 仪器与设备
UPLC Nexera LC-30A系统 日本Shimadzu公司;Q-Exactive Plus质谱(mass spectrometry,MS)仪美国Thermo Scientific公司;ACQUITY UPLC CSH C色谱柱(2.1 mmh100 mm,1.7 µm) 美国Waters公司。
1.3 方法
1.3.1 酸乳制作
参照前期实验进行酸乳制作。牛乳和羊乳经过多次纱布过滤去除固体杂质,然后加热至55 ℃采用二段法均质(20/5 MPa),均质后牛乳和羊乳在90 ℃下高温巴氏杀菌5 min,冷却至45 ℃准备接菌。按照56 mg/L的比例接种丹尼斯克发酵剂MY105,接种后牛乳和羊乳于42 ℃恒温培养,待牛酸乳和羊酸乳pH值达到4.6左右立即于4 ℃冷却停止发酵。生鲜牛乳(CM)、生鲜羊乳(GM)、巴氏杀菌牛乳(PC)、巴氏杀菌羊乳(PG)、牛酸乳(FC)、羊酸乳(FG)和质控(QC),等量取各组样品混合制备。样品于-80 ℃冰箱冷冻保存,备用。每组样品各6 个重复。
1.3.2 脂质提取
采用改良的甲基叔丁基醚(methyl tert-butyl ether,MTBE)提取法,取样本于4 ℃缓慢解冻后,将100 µL样本加入200 µL超纯水,混合,随后加入240 µL预冷甲醇,涡旋混合,再加入800 µL MTBE,涡旋混合,室温放置30 min后,14 000h、10 ℃离心15 min,取上层有机相,氮气吹干。检测分析时加入200 µL异丙醇复溶、涡旋,14 000h、10 ℃离心15 min,取上清液进样分析。
1.3.3 色谱及质谱条件
UPLC条件:ACQUITY UPLC CSH C色谱柱(2.1 mmh100 mm,1.7 µm);流动相:A为含10 mmol/L甲酸铵的体积分数60%乙腈溶液;B为含10 mmol/L甲酸铵的乙腈-异丙醇(1∶9,/)。梯度洗脱程序:0~2 min,70% A、30% B;2~25 min,70%~0% A、30%~100% B;25~35 min,70% A、30% B;柱温45 ℃,流速300 μL/min;进样口温度10 ℃。为避免仪器检测信号波动造成的影响,采用随机顺序进行样本连续分析。
MS条件:采用电喷雾电离源,采用正负离子模式。正离子模式参数设定:离子源温度300 ℃;喷雾电压3.0(正离子模式)、2.5 kV(负离子模式);离子传输管温度350 ℃;鞘气流速45 arb;辅助器流速15 arb;吹扫气流速1 arb;一级MS扫描范围:200~1 800 Da。每次全扫描后采集10 个碎片图谱,一级MS在/200时的分辨率为70 000,二级MS在/200时的分辨率为17 500。
采用LipidSearch进行峰的识别和提取、脂质鉴定(二级鉴定)等。
1.4 数据统计分析

2 结果与分析
2.1 方法学验证
将所有实验样品和QC样本提取到的数据进行PCA,如图1A所示,6 个QC样本紧密聚集在一起,相对位置位于6 组样品中间并且接近原点,表明实验具有良好的重复性。Hotelling通过多元变量建立模型对总体样本进行检验,红色虚线定义了99%置信区间,可用于离群样本的诊断,如图1B所示,所有样本均在99%置信区间内,再次说明实验重复性好。

图1 所有样本的PCA(A)和Hotelling T2检验(B)Fig.1 PCA plot (A) and Hotelling T2 test plot (B) for all samples
2.2 牛乳和羊乳在加工过程中脂质特性变化分析
基于UPLC-Q-Exactive-MS对牛乳、羊乳、巴氏杀菌牛乳、巴氏杀菌羊乳、牛酸乳和羊酸乳进行非靶向脂质组学分析。如图2所示,实验样本中检测到脂质亚类共计27 类,其中检测到数目最多的脂质分子为三酰基甘油酯(triglyceride,TG)共731 种,其次为379 种二酰基甘油酯(diglyceride,DG)和110 种己糖基神经酰胺(hexa-ceramide,HexCer)(其中包括83 种Hex1Cer、25 种Hex2Cer和2 种Hex3Cer),之后依次为82 种磷脂酰胆碱(phosphatidylcholine,PC)、74 种磷脂酰乙醇胺(phosphatidylethanolamine,PE)、66 种鞘磷脂(sphingomyelin,SM)、43 种神经酰胺(ceramides,Cer)、31 种磷脂酰肌醇(phosphatidylinositol,PI)、21 种磷脂酰丝氨酸(phosphatidylserine,PS)、14 种鞘氨醇(sphingosine,Sph)、9 种单酰基甘油酯(monoglyceride,MG)、6 种溶血性磷脂酰胆碱(lysophosphatidylcholine,LPC)、6 种酵母甾醇(zymosterol,ZyE)、5 种溶血性磷脂酰乙醇胺(lysophosphatidylethanolamine,LPE)、5 种单半乳糖二酰基甘油(monogalactosylmonoacylglycerol,MGDG)、5 种磷脂酰甘油(phosphatidylglycerol,PG)、4 种豆甾醇酯(stigmasterol ester,StE)、4 种蜡酯(wax exters,WE)、3 种磷酸神经酰胺(ceramides phosphate,CerP)、3 种心磷脂(cardiolipin,CL)、2 种磷脂酰肌醇一磷酸(phosphatidylinositol(4)phosphate,PIP)、1 种神经节苷脂(gangliosides,GD1a)、1 种溶血性磷脂酰甘油(lysophosphatidylglycerol,LPG)、1 种磷脂酰肌醇(lysophosphatidylinositol,LPI)、1 种植物鞘氨醇(phytosphingosine,phSM)。牛乳是一种复杂的脂质混合物,含有数千种脂质,虽然研究众多,但许多低丰度的脂类仍然未知。Li Mohan等对牛初乳和成熟乳脂质进行全面表征,共检测出13 亚类335 种脂质分子;Liu Zhiqian等对牛乳中TG进行综合表征共鉴定出3 454 种脂质分子,并对牛乳中磷脂、鞘脂、糖脂、Cer进行综合表征,共鉴定出15 亚类514 种脂质分子。本研究在正负离子模式下共检测到1 607 种脂质分子,对牛乳和羊乳在加工过程中各阶段的脂质分子及动态变化进行表征,结果表明牛乳和羊乳在加工制备酸乳过程中脂质家族具有巨大的多样性和复杂性。

图2 牛乳和羊乳的脂质亚类分析Fig.2 Analysis of lipid subclasses in cow and goat milk
2.3 热处理对牛乳和羊乳的脂质组学影响
如图3A所示,TG(相对含量90%)是牛乳中检测到最丰富的脂质成分,其次是DG(8%),这表明牛乳脂肪主要以甘油酯形式存在,约占总脂质含量的98%,而其他脂质(2%)主要以磷脂、鞘脂、固醇类和糖脂形式存在,这些脂质是乳脂肪球膜(milk fat globule membrane,MFGM)的主要成分,确保了乳的水包油乳液的稳定性。在2%的其他脂质中,磷脂含量最为丰富,约占68%,其中亚类及相对含量分别为PC(26%)、PE(15%)、SM(14%)、PG(10%)、PI(2%)和PS(1%),这与Zhang Xinhao等测定的牛乳磷脂组成较为相似,但PS含量相差较大,这可能是由地域、泌乳期、气候、季节、饲料以及乳牛个体差异导致,这些因素均会对乳脂成分产生显著影响;鞘脂相对含量约占24%,包括Hex1Cer(20%)、CerP(2%)、Cer(1%)和Hex2Cer(1%);固醇类约占4%,包括ZyE(3%)和StE(1%);糖脂主要以MGDG(3%)形式存在;其余12 种亚类含量极微量,共计1%。如图3B所示,与生牛乳相比,热处理后的巴氏杀菌牛乳DG相对含量下降1%,TG相对含量没有改变,PC、SM、PI和MGDG相对含量有极小幅度上调,PE、PG、Hex1Cer和ZyE极小幅度下调。如图3C、D所示,与生羊乳相比,巴氏杀菌羊乳DG相对含量也降低1%,TG相对含量不变,PC和SM相对含量分别上调1%,Hex1Cer相对含量下调2%。表明热处理对牛乳和羊乳脂质特性的影响基本趋同,即热处理对PC、SM相对含量有轻微上调作用,这与Gallier等探究生牛乳和巴氏杀菌后牛乳的极性脂质变化一致;而DG、Hex1Cer相对含量轻微下降、TG相对含量基本保持不变,这一结果与Xu Qingbiao等研究结果一致;其他磷脂、鞘脂、糖脂在热处理后没有变化。总的来说,高温巴氏杀菌处理未导致牛乳和羊乳脂质亚类的显著变化。Bezie等研究表明牛乳中的脂肪基本不会受到热处理影响。


图3 热处理前后牛乳和羊乳的脂质特性比较Fig.3 Comparison of lipid properties of cow and goat milk before and after heat treatment
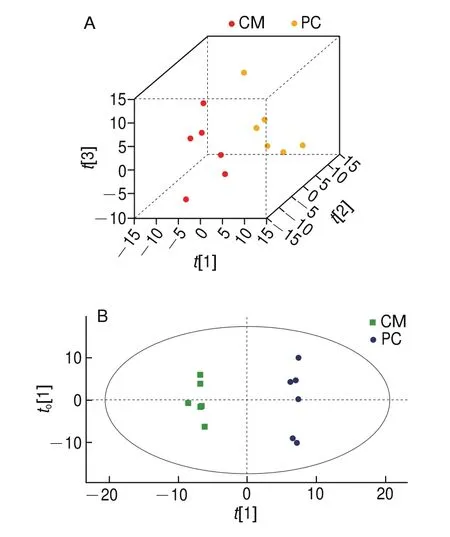
PCA根据样本的相似性或不同性对样本进行分组。如图4A所示,[1]、[2]和[3]是对模型贡献率最大的3 个PC,前3 个PC累计贡献率达到49.4%,CM组和PC组之间观察到较明显差异,表明高温巴氏杀菌对牛乳的脂质分子产生一定影响。进一步使用有监督的OPLSDA,如图4B所示,CM组和PC组明显分离,再次验证了两组样本脂质分子的差异性。如图4C、D所示,GM组和PG组明显分离,表明高温巴氏杀菌使羊乳脂质分子也发生变化。

图4 热处理前后牛乳和羊乳的脂质特性PCA和OPLS-DAFig.4 PCA and OPLS-DA plots for lipid properties of cow and goat milk before and after heat treatment
选择满足<0.05并且VIP>1的脂质分子,分别在CM组与PC组、GM组与PG组中筛选出65 种和170 种显著差异脂质分子。为使表征高温巴氏杀菌对乳脂脂质分子的影响更具普遍性和科学性,将上述牛乳和羊乳组的显著差异脂质分子进行交集,根据丰度变化趋势,进一步筛选出27 种脂质分子作为判别热处理乳脂的潜在生物标志物。如图5所示,热处理后,这27 种脂质分子在牛乳组和羊乳组中的变化趋势相同,其丰度均呈下降趋势,表明高温巴氏杀菌虽然未对牛乳和羊乳的脂质亚类特性产生显著影响,但对乳脂分子层面产生影响。

图5 热处理前后牛乳(A)和羊乳(B)中27 种潜在标志物热图变化Fig.5 Heat map showing changes in 27 potential markers in cow milk (A) and goat milk (B) before and after heat treatments
27 种脂质分子以TG和PE为主,这可能反映了热处理主要破坏了乳脂中TG和PE的脂质分子结构,而其他脂质亚类分子受热处理影响很小。这些脂质分子丰度的显著减少可能是因为在高温巴氏杀菌条件下,MFGM成分改变,蛋白质含量升高而脂质含量降低,热处理对MFGM和脂肪球的损害导致部分游离TG和磷脂暴露;此外,牛乳中存在大量脂蛋白脂肪酶,具有脂肪分解活性,且对热处理具有抗性,经过高温巴氏杀菌处理后,残存的脂肪酶与从MFG(M)中游离出的TG和磷脂直接接触并进行脂解,生成游离脂肪酸,从而导致27 种脂质分子丰度的显著降低。事实上,比巴氏杀菌更高温度的杀菌工艺对牛乳中的脂解程度更剧烈,因为更高的温度导致MFGM的脂质表面暴露更多,从而可能导致更多的脂质分子暴露。
27 种脂质分子中脂肪酸链大多为C~C、不饱和脂肪酸链多为C和C,以及极少数的奇链脂肪酸链(C和C),它们经热处理后被脂解生成游离脂肪酸,而游离脂肪酸较酯类更易氧化,尤其是短链和中链脂肪酸(C~C),这些游离脂肪酸和氧化的脂质产物导致风味缺陷,这有可能是高温巴氏杀菌后牛乳和羊乳产生“熟味”的原因,表明高温巴氏杀菌处理使脂质分子产生一定程度的脂解和氧化,对牛乳和羊乳风味产生不利影响。
2.4 发酵对牛乳和羊乳的脂质组学影响
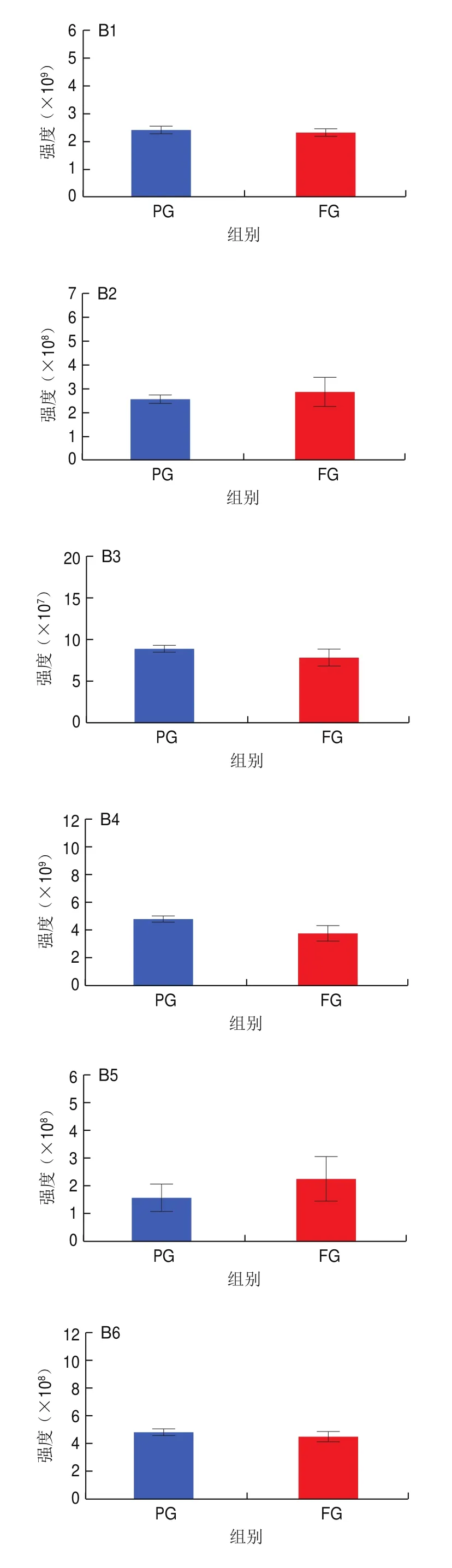
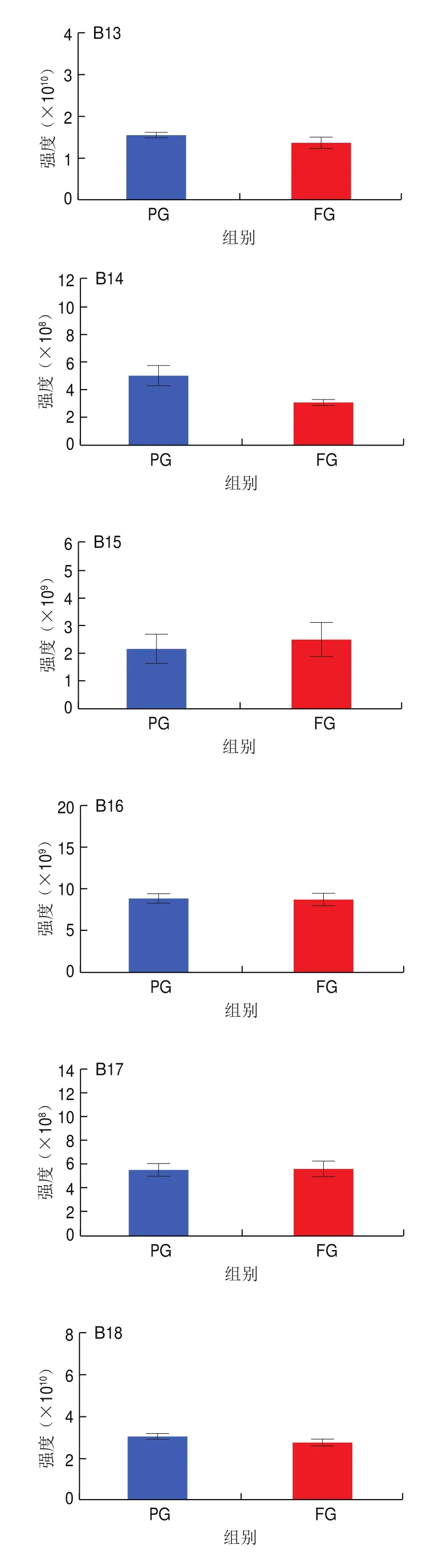
如图6A所示,巴氏杀菌牛乳经发酵后,WE含量上升,CerP和MGDG含量基本一致,其余24 种脂质亚类含量均明显下降,尤其是CL、GD1a、Hex2Cer、LPC、LPE、LPG、LPI、MG、PC、PE、phSM、PI、SM、StE和ZyE的含量下降了50%以上,表明发酵显著改变了牛乳的脂质特性。巴氏杀菌羊乳经发酵后脂质特性也表现出显著变化。如图6B所示,巴氏杀菌羊乳在发酵后CerP、GD1a、PG、PIP、StE和ZyE含量上升,MG、MGDG和Hex3Cer含量基本没有变化,其余18 种脂质亚类含量均下降,尤其是LPC、LPE、LPG和LPI含量大幅下降。这些结果共同证明了发酵显著改变了乳脂特性,值得注意的是,发酵对牛乳和羊乳在一些脂质亚类上表现出一致的变化趋势,TG、DG、PC、PE、SM、PS、PI、CL、LPC、LPE、LPG、LPI、Cer、Hex1Cer、Hex2Cer、phSM和SPH均表现出下降趋势,表明嗜热链球菌和保加利亚乳杆菌代谢消耗这些脂质,使其含量降低,值得注意的是,这两种乳酸菌对LPC、LPE、LPG和LPI的代谢程度最剧烈,这可能是由于溶血型磷脂的结构由一条烷基链或酰基链和极性头基形成,更易被嗜热链球菌和保加利亚乳杆菌分解利用。溶血型磷脂是一类生物活性脂质,LPC是大脑发育和生长过程中脂肪酸传递的重要途径,在大脑生长中扮演着重要角色;LPE具有抗细胞凋亡的作用,被证实可以诱导PC12细胞丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPK)活化,从而激活MAPK级联反应抑制血清剥夺诱导的细胞凋亡;LPG参与心血管疾病的发病机制;LPI可以上调脂肪生成基因的表达,与人类肥胖呈正相关。这些结果表明发酵降低了乳中溶血型磷脂含量,从而影响了其功能特性。

图6 发酵前后牛乳和羊乳中27 种脂质亚类含量变化Fig.6 Changes in the contents of 27 lipid subclasses in cow and goat milk before and after fermentation







对PC组和FC组脂质亚类进行PCA,如图7A所示,前3 个PC累计贡献率达到65.5%,在[1](51.6%)上观察到两组存在明显差异,OPLS-DA得分图(图7B)中两组样本点分离良好,表明发酵对牛乳脂质产生显著影响。如图7C所示,PG组和FG组PCA中前3 个PC累计贡献率为45.2%,PG组和FG组样本点相互包含,两组样本无明显分离,但在有监督的OPLS-DA得分图(图7D)中,PG组和FG组有效分离。

图7 发酵前后牛乳和羊乳的脂质特性PCA和OPLS-DAFig.7 PCA and OPLS-DA plots for lipid properties of cow and goat milk before and after fermentation

选择满足<0.05且VIP>1的脂质分子,在PC组和FC组中筛选出191 种显著差异脂质分子,在PG组和FG组中筛选出46 种显著差异脂质分子。根据发酵前后的变化趋势,在上述191 种和49 种显著差异脂质分子中进一步筛选出23 种脂质分子作为表征发酵对乳脂影响的潜在生物标志物。如图8所示,经发酵后,这23 种潜在生物标志物丰度在牛酸乳和羊酸乳中均呈下降趋势。这23 种潜在生物标志物由7 种PE、4 种溶血型磷脂、4 种PI、3 种TG、2 种SM、2 种PS、1 种PC构成,这一结果与发酵对乳脂亚类的影响结果一致,再次验证了发酵降低了TG、溶血型磷脂、磷脂(PE、PC、PS、PI和SM)的含量。磷脂对脂类代谢失调、肠道生态失调、炎症、心血管疾病、肠道健康和神经发育有积极影响,其损失可能影响食物的营养价值和生物活性。这些潜在标志物的脂肪酸链大多由C和C家族组成,而且基本不包括短中链脂肪酸(C~C)和奇链脂肪酸(C和C),表明嗜热链球菌和保加利亚乳杆菌基本不代谢短中链脂肪酸和奇链脂肪酸,它们更偏向于代谢C和C家族脂质。Furse等的研究也表明牛乳和酸乳中的奇链脂肪酸含量相似。

图8 发酵前后牛乳(A)和羊乳(B)中23 种潜在标志物热图变化Fig.8 Heat map showing changes in 23 potential markers in cow milk (A) and goat milk (B) before and after fermentation
3 结 论
用UPLC-Q-Exactive-MS联用技术表征了“牛乳-巴氏杀菌牛乳-牛酸乳”和“羊乳-巴氏杀菌羊乳-羊酸乳”的脂质特性动态变化,共检测出27 种脂质亚类1 607 种脂质分子。结果表明,巴氏杀菌热处理对牛乳和羊乳的脂质亚类基本无影响,但对脂质分子产生显著影响,筛选出27 种脂质分子可作为鉴定生与巴氏杀菌牛乳和羊乳的潜在生物标志物;而发酵对牛乳和羊乳的脂质特性具有显著影响,大部分脂质亚类表现出下调趋势,筛选出23 种脂质分子可作为鉴定巴氏杀菌与发酵牛乳和羊乳的潜在生物标志物。本研究为酸乳加工过程中热处理和发酵阶段对乳脂的影响提供了分子基础,有利于加深对不同阶段对酸乳终产品质量和营养特性影响的理解,并且潜在生物标志物可用于商业化生产酸乳过程中判断加工工艺的终点及产品状态。
猜你喜欢
中国果业信息(2021年7期)2021-12-01
中国食品(2021年11期)2021-06-23
大众健康(2020年11期)2020-11-13
食品工业科技(2020年18期)2020-09-23
乳业科学与技术(2019年5期)2019-12-24
乳业科学与技术(2019年4期)2019-12-23
中国中医基础医学杂志(2019年9期)2019-10-21
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年13期)2014-03-11
