
生化黄腐酸对仙居鸡生产性能、屠宰性能、血清生化指标、肠道消化酶活性以及盲肠微生物菌群和代谢组的影响
2022-09-01 03:21冯佩诗揭晓泸卢立志
动物营养学报 2022年8期
冯佩诗 揭晓泸 叶 轩 田 勇 卢立志* 王 平*
(1.浙江工业大学药学院,杭州 310014;2.浙江省仙居种鸡场,仙居 311719;3.浙江省农业科学院畜牧兽医研究所,杭州 310021)
蛋鸡在产蛋后期由于生理机能的衰退,疾病抵抗力的下降,容易受病原微生物的侵袭而发生疾病。传统上,会在畜禽饲粮中加入亚治疗水平的抗生素以促进畜禽生长和预防疾病。然而,抗生素的使用是一把双刃剑,其过量使用和滥用容易造成细菌耐药性的提高。故自2020年1月始,我国开始全面禁用饲用抗生素。因此,亟需开发一种绿色、安全、有效的饲用抗生素替代品来保障畜禽生产。
生化黄腐酸(biochemical fulvic acid,BFA)是腐植酸的一种,是通过人工模拟自然界腐植酸的形成过程,以秸秆、糖蜜等为主要原料,通过特定的微生物发酵产生的一类含有多种活性基团的有机化合物[1-2]。近年研究指出,在奶牛和肉牛饲粮中添加适量的BFA可增加奶牛产奶量,提高机体免疫能力和抗应激能力,降低饲养场的氨排放量[3]。另外,BFA可提高小鼠巨噬细胞的吞噬指数、碳粒廓清指数和血细胞凝集效价,促进机体非特异性免疫[4]。陆秀玉[5]研究发现,饲粮中添加0.03%的BFA可提高肉仔鸡的平均日增重(ADG),降低料重比(F/G),促进肉仔鸡的生长。但是,目前国内外关于BFA用作饲料添加剂的适宜剂量和研究结论存在较大的争议。因此,本试验以仙居鸡为研究对象,研究不同剂量的BFA对仙居鸡生产性能、屠宰性能、血清生化指标、肠道消化酶活性以及盲肠微生物菌群和代谢组的影响,以期为BFA在家禽生产上的应用提供参考依据。
1 材料与方法
1.1 试验材料
试验所用BFA为棕黄色粉末状,为河北某科技有限公司产品,纯度为99%。
1.2 试验设计
试验选用平均体重为(1.83±0.33)kg、524日龄健康的仙居母鸡240羽,由浙江省仙居种鸡场提供。试验鸡随机分为4组,每组6个重复,每个重复10羽。对照组饲喂基础饲粮,试验组分别在基础饲粮中添加500(BFA500组)、1 000(BFA1000组)、1 500 mg/kg(BFA1500组)的BFA。在参考NRC(1994)和《鸡饲养标准》(NY/T 33—2004)的基础上配制基础饲粮,其组成及营养水平见表1。预试期3 d,正试期42 d。

表1 基础饲粮组成及营养水平(风干基础)
1.3 饲养管理
试验在浙江省仙居种鸡场进行,鸡舍为砖混结构,采用3层笼养,自然光照加人工光照,光照制度为18 h光照:6 h黑暗,相对湿度为55%~65%。试验鸡自由采食和饮水,按照常规方法进行饲养管理。
1.4 样品采集
试验第42天,每个重复随机选择接近平均体重的2只试验鸡,使用不含抗凝剂的采血管从颈静脉采血10 mL,经静置2 h后,3 000 r/min离心10 min,收集血清后分装于离心管中,-20 ℃保存,用于后续血清生化指标测定。放血致死后,1只试验鸡用于测定屠宰性能指标,另1只试验鸡分别收集十二指肠、空肠、回肠和盲肠内容物,液氮速冻后,-80 ℃保存,用于后续肠道消化酶活性以及盲肠微生物菌群和代谢组测定。
1.5 测定指标和方法
1.5.1 生产性能测定
每天观察鸡的生长发育、发病及死亡情况,准确记录产蛋总数、蛋重、死淘数和采食量。于试验第41天的21:00禁食,次日08:00以重复为单位称重,计算平均日采食量(ADFI)、ADG、F/G、产蛋率和料蛋比。
1.5.2 屠宰性能测定
参照《家禽生产性能名词术语和度量统计方法》(NY/T 823—2020)测定活重、屠体重、半净膛重、全净膛重、两侧胸肌重、两侧腿肌重、腹脂重和脾脏重,计算屠宰率、半净膛率、全净膛率、胸肌率、腿肌率和腹脂率,计算公式如下:
屠宰率(%)=(屠体重/活重)×100;半净膛率(%)=(半净膛重/活重)×100;全净膛重(%)=(全净膛重/活重)×100;胸肌率(%)=(两侧胸肌重/全净膛重)×100;腿肌率(%)=(两侧腿肌重/全净膛重)×100;腹脂率(%)=[腹脂重/(全净膛重+腹脂重)]×100。
1.5.3 血清生化指标测定
血清谷丙转氨酶(ALT)、谷草转氨酶(AST)活性和总蛋白(TP)、尿酸(UA)含量采用宁波美康生物科技股份公司试剂盒,使用全自动生化分析仪(HITACHI-7020,日本)测定,测定方法分别为丙氨酸底物法、天门冬氨酸底物法、双缩脲法和脲酶-谷氨酸脱氢法。
血清白细胞介素-2(IL-2)、白细胞介素-6(IL-6)、白细胞介素-10(IL-10)和肿瘤坏死因子-α(TNF-α)含量使用酶联免疫吸附测定(ELISA)试剂盒(上海赫澎生物科技有限公司)测定,并按照试剂盒说明书进行操作。
1.5.4 肠道消化酶活性测定
采集后的肠道内容物于低温下称取0.1 g,使用试剂盒(南京建成生物工程研究所)分别测定糜蛋白酶、胰蛋白酶和淀粉酶活性,并按照试剂盒说明书进行操作。
1.5.5 盲肠微生物菌群测定
对照组和BFA150组的盲肠微生物总DNA使用E.Z.N.A.®粪便DNA试剂盒(D4015,Omega公司,美国)提取。所提取的总DNA由上海美吉生物有限公司使用338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)对细菌16S rDNA V3~V4可变区进行测序。测序平台为Illumina NovaSeq PE250,原始数据(raw reads)经拼接、质控和过滤后,使用美吉生物云平台(https://cloud.majorbio.com)进行数据分析。
1.5.6 盲肠代谢组测定
对照组和BFA150组的盲肠内容物的制备、代谢物鉴定和定量由上海美吉生物有限公司测定,仪器平台为AB SCIEX公司的超高效液相色谱串联飞行时间质谱系统。原始数据经去噪平滑、基线校正、重叠识别获得的数据矩阵用80%规则来去除缺失值,再进行填补空缺值,用总和归一化法对样本质谱峰的响应强度进行归一化,同时删除质量控制(QC)样本相对标准偏差(RSD)>30%的变量,并进行log10对数化处理,得到最终用于后续分析的数据矩阵。同时将质谱信息与代谢公共数据库HMDB(http://www.hmdb.ca/)和Metlin(https://metlin.scripps.edu/)数据库进行匹配,得到代谢物信息。预处理后的数据上传美吉生物云平台(https://cloud.majorbio.com)进行数据分析。
1.6 统计分析
数据经Excel 2019初步整理后,用SPSS 22.0统计分析软件进行单因素方差分析,用Scheffe法进行多重比较,数据以“平均值±标准差”表示,P<0.05为差异显著。
2 结果与分析
2.1 BFA对仙居鸡生产性能的影响
由表2可知,饲粮中添加BFA对仙居鸡的ADG、ADFI、F/G、产蛋率和料蛋比无显著影响(P>0.05)。但是,随着饲粮中BFA添加量的提高,ADG有所提高,而F/G有所降低。

表2 BFA对仙居鸡生产性能的影响
2.2 BFA对仙居鸡屠宰性能的影响
由表3可知,饲粮中添加BFA对仙居鸡的半净膛率、全净膛率、胸肌率、腿肌率、腹脂率和脾脏重无显著影响(P>0.05)。BFA1000组的屠宰率显著低于对照组(P<0.05)。

表3 BFA对仙居鸡屠宰性能的影响
2.3 BFA对仙居鸡血清生化指标和细胞因子含量的影响
由表4可知,饲粮中添加BFA对血清ALT、AST活性和TP、UA含量无显著影响(P>0.05)。

表4 BFA对仙居鸡血清生化指标的影响
由表5可知,BFA500组、BFA1000组和BFA1500组的血清TNF-α含量显著低于对照组(P<0.05)。BFA500组的血清IL-6含量显著低于BFA1500组(P<0.05),但与其余2组无显著差异(P>0.05)。饲粮中添加BFA对血清IL-2和IL-10含量无显著影响(P>0.05)。

表5 BFA对仙居鸡血清细胞因子含量的影响
2.4 BFA对仙居鸡肠道消化酶活性的影响
由图1可知,饲粮中添加BFA对十二指肠、空肠、回肠消化酶活性的影响有一定差异。在十二指肠,BFA500组、BFA1000组和BFA1500组的糜蛋白酶活性显著低于对照组(P<0.05);饲粮中添加BFA对胰蛋白酶和淀粉酶活性无显著影响(P>0.05)。在空肠,BFA500组、BFA1000组和BFA1500组的糜蛋白酶活性显著高于对照组(P<0.05),饲粮中添加BFA对胰蛋白酶和淀粉酶活性无显著影响(P>0.05)。在回肠,BFA1000组的糜蛋白酶活性显著高于对照组(P<0.05),BFA500的淀粉酶活性显著高于对照组(P<0.05),BFA500组、BFA1000组和BFA1500组的胰蛋白酶活性显著高于对照组(P<0.05)。
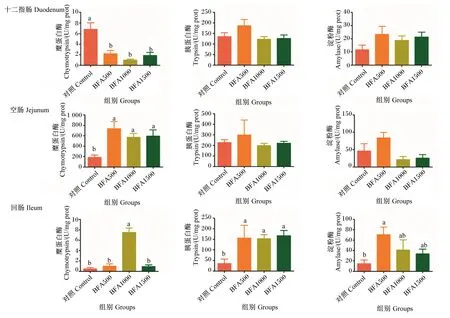
数据柱标相同小写字母表示差异不显著(P>0.05),不同小写字母表示差异显著(P<0.05)。
2.5 BFA对仙居鸡盲肠微生物菌群的影响
2.5.1 BFA对盲肠微生物群落丰富度和多样性的影响
Illumina NovaSeq PE250高通量测序结果表明,从对照组和BFA1500组盲肠内容物样本中共获得1 210 413条clean reads,平均每个样品121 041条clean reads,平均序列长度为420 bp。由表6可知,α-多样性结果显示,对照组和BFA150组的Sobs指数、Shannon指数、Simpson指数、Ace指数、Chao指数、Shannoneven指数、Simpsoneven指数和覆盖率无显著差异(P>0.05)。
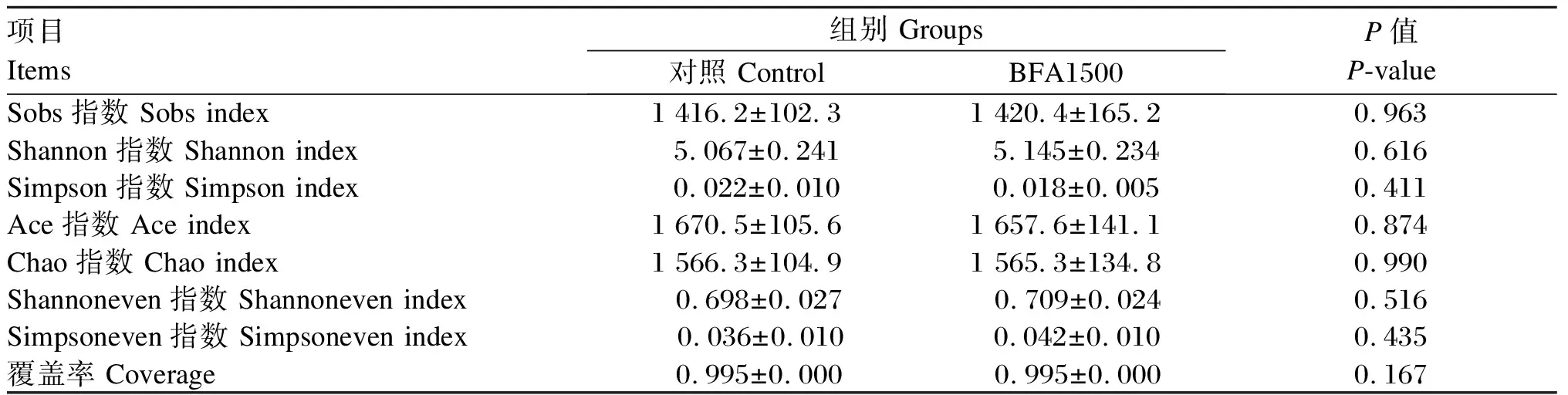
表6 BFA对仙居鸡盲肠微生物α-多样性的影响
2.5.2 BFA对盲肠微生物菌群组成及结构的影响
基于97%相似度进行操作分类单位(OTU)聚类,共获得4 243个OTU,其中对照组独有的OTU数量为1 174个,BFA150组独有的OTU数量为1 156个(图2-A)。采用RDP classifier贝叶斯算法对97%相似水平的OTU序列进行分类学分析,在门水平上共鉴定出20个菌门,并且以厚壁菌门、拟杆菌门、放线菌门和脱硫杆菌门等为优势菌门(图2-B)。在属水平上共鉴定出229个菌属,其中优势菌属为拟杆菌属、乳杆菌属、理研菌科RC9肠道群和考拉杆菌属等(图2-C)。
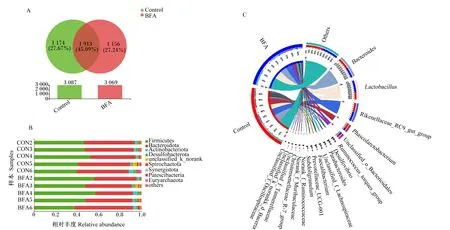
Control、CON:对照组 control group;BFA:BF150组 BF150 group。下图同 the same as below。
2.5.3 微生物富集差异
进一步用LEfSe并以线性判别分析(LDA)值>2为阈值分析了对照组和BFA150组之间的微生物富集差异。由图3可知,在属水平上,对照组中富集的生物标记物为肠球菌属、草酸杆菌属、奇异菌属;BFA1500组富集的生物标记物为罕见小球菌属、Catenibacillus、蛋白胨链球菌科未知属和毛螺菌科-UCG-002(图3)。

g_Enterococcus:肠球菌属;g_Oxalobacter:草酸杆菌属;g_Atopobium:奇异菌属;g_Subdoligranulum:罕见小球菌属;g_Lachnospiraceae_UCG-002:毛螺菌科-UCG-002;g_norank_f_Peptococcaceae:蛋白胨链球菌科未知属。
2.6 BFA对仙居鸡盲肠代谢组的影响
2.6.1 差异代谢物的筛选与鉴定
为了研究BFA对盲肠微生物代谢的影响,我们利用非靶向代谢组学同时检测分析了盲肠内容物的小分子代谢物。由图4可知,与对照组相比,在BFA1500组共检测到了1 586个差异离子峰,并筛选出27种差异代谢物[变量重要性投影(VIP)>2且P<0.05]。在这些差异代谢物中,16种被下调,11种被上调。其中,得分最高的前5种代谢物为磷脂酰乙醇胺PE[14∶1(9Z)/16∶1(9Z)](VIP=4.278 5)、硫代硫酸盐(VIP=3.388 3)、蕨素E(VIP=3.386 6)、咖啡因(VIP=3.373 0)和单取代基甘油糖脂MG(0∶0/16∶0/0∶0)(VIP=3.169 3)。
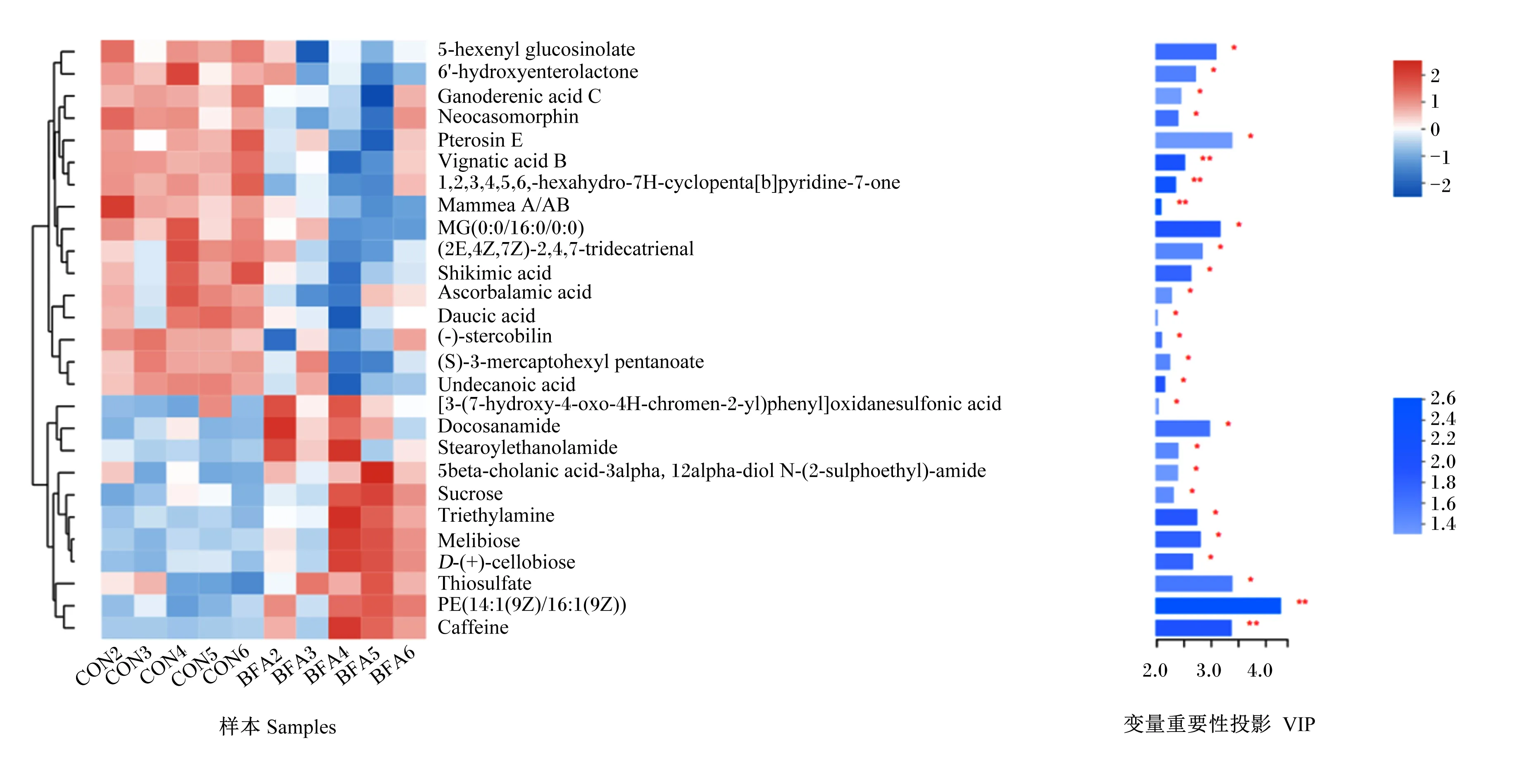
5-hexenyl glucosinolate:5-己烯基硫苷;6’-hydroxyenterolactone:6-羟基肠内酯;Ganoderenic acid C:灵芝烯酸C;Pterosin E:蕨素E;1,2,3,4,5,6,-hexahydro-7H-cyclopenta[b]pyridine-7-one:1,2,3,4,5,6-六氢-7H-环戊并[b]吡啶-7-酮;MG(0∶0/16∶0/0∶0):单取代基甘油糖脂MG(0∶0/16∶0/0∶0);(2E,4Z,7Z)-2,4,7-tridecatrienal:(E,Z,Z)-2,4,7-十三碳三烯醛;Shikimic acid:莽草酸;Ascorbalamic acid:阿斯考巴拉酸;(-)-stercobilin:粪胆色素;(S)-3-mercaptohexyl pentanoate:3-巯基己醇戊酸酯;Undecanoic acid:十一酸;[3-(7-hydroxy-4-oxo-4H-chromen-2-yl)phenyl]oxidanesulfonic acid:[3-(7-羟基-4-氧代-4H-铬-2-基)苯基]烷氧磺酸;Docosanamide:二十二酰胺;Stearoylethanolamide:硬脂酸单乙醇酰胺;5beta-cholanic acid-3alpha:去氧胆酸;12alpha-diol N-(2-sulphoethyl)-amide:甘氨去氧胆酸;Sucrose:蔗糖;Triethylamine:三乙胺;D-(+)-cellobiose:D-(+)-纤维二糖;Thiosulfate:硫代硫酸盐;PE[14∶1(9Z)/16∶1(9Z)]:磷脂酰乙醇胺PE[14∶1(9Z)/16∶1(9Z)];Caffeine:咖啡因。
由图5可知,利用KEGG数据库对差异代谢物进行注释,共获得8个差异的代谢通路,分别为苯丙氨酸、酪氨酸和色氨酸的生物合成,咖啡因代谢,碳水化合物的消化和吸收,味觉传导,硫代谢,半乳糖代谢,植物次生代谢产物的生物合成以及ABC转运蛋白。

Tropane, piperidine and pyridine alkaloid biosynthesis:托烷、哌啶和吡啶生物碱的生物合成;Taste transduction:味觉传导;Sulfur metabolism:硫代谢;Steroid hormone biosynthesis:类固醇激素生物合成;Starch and sucrose metabolism:淀粉和蔗糖代谢;Phosphotransferase system(PTS):磷酸转移酶系统;Phenylalanine, tyrosine and tryptophan biosynthesis:苯丙氨酸、酪氨酸和色氨酸的生物合成;Lysine degradation:赖氨酸降解;Galactose metabolism:半乳糖代谢;Carbohydrate digestion and absorption:碳水化合物的消化和吸收;Caffeine metabolism:咖啡因代谢;Biosynthesis of plant secondary metabolites:植物次生代谢物的生物合成;Biosynthesis of plant hormones:植物激素的生物合成;Biosynthesis of phenylpropanoids:苯丙烷生物合成;Biosynthesis of alkaloids derived from shikimate pathway:莽草类生物碱的生物合成;Biosynthesis of alkaloids derived from ornithine, lysine and nicotinic acid:鸟氨酸、赖氨酸和烟酸生物碱的生物合成;Biosynthesis of alkaloids derived from histidine and purine:组氨酸和嘌呤类生物碱的生物合成;Aminobenzoate degradation:氨基苯甲酸酯降解;ABC transporters:ABC转运蛋白。
2.6.2 代谢物组成与微生物群落组成的相关性分析
将样品中相对丰度前20的菌属与上述VIP前20的差异代谢物进行相关性分析。结果显示,罕见小球菌属与蕨素E、单取代基甘油糖脂MG(0∶0/16∶0/0∶0)、(E,Z,Z)-2,4,7-十三碳三烯醛、莽草酸、灵芝烯酸C、Vignatic acid B呈负相关,而与硬脂酰基乙醇酰胺、二十二酰胺、D-(+)-纤维二糖、三乙胺、咖啡因、磷脂酰乙醇胺PE(14∶1(9Z)/16∶1(9Z))、蜜二糖和脱氧胆酸呈正相关。此外,考拉杆菌属恰好相反,与蕨素E、单取代基甘油糖脂MG(0∶0/16∶0/0∶0)、Vignatic acid B和Mammea A/AB呈正相关,而与二十二酰胺、D-(+)-纤维二糖、蔗糖、三乙胺、磷脂酰乙醇胺PE[14∶1(9Z)/16∶1(9Z)]和蜜二糖呈负相关。
3 讨 论
3.1 BFA对仙居鸡生产性能和屠宰性能的影响
饲料成本通常占家禽养殖成本的70%以上,因此,F/G是衡量效益的重要指标,F/G越小,利润越高。众多研究表明,BFA具有改善家禽的生产性能、增强机体免疫力和降低死亡率等作用。陆秀玉[4]研究指出,饲粮中添加0.03%的BFA可显著提高肉仔鸡的ADG。李素芬等[5]研究发现,饲粮中添加0.5%~1.0%的BFA可提高肉仔鸡的ADG,改善饲料报酬和经济效益。这可能与BFA改变肉鸡肠道黏膜形态,增加绒毛高度,从而提高饲料的消化和吸收有关[6]。在本试验中,虽然饲粮中添加BFA可改善仙居鸡的ADG和F/G,但是未达到显著水平。这可能与试验所选择的仙居鸡处于产蛋后期,生长速度放慢有关[7]。
屠宰率和全净膛率是衡量家禽产肉性能的重要指标,本试验结果显示,虽然BFA1000和BFA1500组的屠宰率分别为90.27%和90.67%,低于对照组的92.84%,但是在全净膛率上,各组均达68%以上,组间无显著差异。屠宰性能结果与黄玲玲[8]研究结果相当,表明BFA对屠宰性能无明显影响。
3.2 BFA对仙居鸡血清生化指标和细胞因子含量的影响
血清生化指标是反映动物健康状况的重要指标。本试验测定了仙居鸡的血清AST、ALT活性和TP、UA含量,各组之间血清生化指标无显著差异,表明饲粮中添加BFA对血清生化指标无负面影响。
IL-2和IL-10是免疫网络中重要的调节因子,它们在抗感染和遏制炎症反应上发挥重要的作用[9-10]。TNF-α和IL-6则是促炎反应中重要的细胞因子,它们可与其他免疫细胞结合以维持慢性炎症[11]。本试验结果显示,饲粮中添加BFA可在一定程度上提高仙居鸡血清IL-2含量,并且降低血清IL-6和TNF-α含量,提示BFA可通过调节机体炎症因子水平从而增强仙居鸡的细胞免疫功能。Liu等[12]的研究结果也表明,BFA可显著降低大鼠中性粒细胞、白细胞和单核细胞数量,显著提高血清免疫球蛋白A(IgA)、免疫球蛋白G(IgG)、免疫球蛋白M(IgM)和IL-2含量,从而增强机体防御能力和免疫水平。另外,BFA可提高小鼠巨噬细胞的吞噬能力,对机体的非特异性免疫有一定的促进作用[13]。BFA对畜禽免疫功能具有积极影响,表明其具有作为功能性饲料添加剂改善畜禽健康的价值。
3.3 BFA对仙居鸡肠道消化酶活性的影响
消化酶是饲料消化吸收的媒介,其活性是反映机体消化能力的重要指标。本试验结果显示,空肠中糜蛋白酶、胰蛋白酶和淀粉酶活性均高于十二指肠或回肠,可见空肠是营养物质消化和吸收的主要场所[14]。研究显示,BFA是一种安全的促生长添加剂,可通过提高肠道消化活性增强机体对营养物质的消化吸收,从而提高生产性能[15]。在本试验中,饲粮中添加BFA可提高空肠糜蛋白活性和回肠糜蛋白酶、胰蛋白酶和淀粉酶活性,这与侯少岩等[16]、朱荣生等[17]研究结果一致。乌兰等[18]报道,饲料中添加0.30%的黄腐酸有助于提高鲫鱼的肝胰脏蛋白质酶、淀粉酶和肠道淀粉酶活性。由于BFA本身不含有消化酶,但可在一定程度上提高肠道消化酶活性,提示BFA可能刺激肠道消化酶的分泌,或者刺激内源消化酶的合成,但具体机制仍需要进一步研究。
3.4 BFA对仙居鸡盲肠微生物菌群和代谢组的影响
罕见小球菌属是近年来在人类排泄物中新分离的严格厌氧的革兰氏阴性菌,呈多样性的球菌形态[19]。罕见小球菌属广泛存在于健康成人肠道内,并与肠道通透性息息相关[20]。研究指出,罕见小球菌属可通过发酵反应产生丁酸、乳酸以及微量的乙酸和琥珀酸[19],这些短链脂肪酸是肠上皮细胞的主要能量来源,参与维持肠道黏膜屏障、调节肠道炎症以及增加胃肠动力等[21]。本试验发现,饲粮中添加BFA可增加仙居鸡盲肠罕见小球菌属相对丰度,并且罕见小球菌属与甘氨去氧胆酸呈显著正相关。目前已知只有极少数低相对丰度的菌属能够进行次级胆汁酸代谢[22]。这提示BFA可能通过提高罕见小球菌属相对丰度,增加肠道短链脂肪酸和次级胆汁酸含量,进而影响机体的能量和脂肪代谢。
Catenibacillus和毛螺菌科-UGG-002是在BFA组富集的2个属于毛罗杆菌科的菌属。其中,Catenibacillus是一种严格厌氧的革兰氏阳性菌,主要以链状出现[23]。研究指出,Catenibacillus不但可将葛根素去糖基化为大豆黄酮,还参与其他芳香族C-葡萄糖苷和类黄酮O-葡萄糖苷的去糖基化[24];其代谢产物原儿茶酸、3,4-二羟基苯乙酸和3,4-二羟基苯基丙酸已被证实可通过下调促炎细胞因子白细胞介素-1β(IL-1β)、IL-6、白细胞介素-8(IL-8)和TNF-α的表达来发挥抗炎作用,并且通过降低一氧化氮(NO)、过氧化氢(H2O2)和丙二醛含量来发挥抗氧化作用[25]。
本试验结果显示,饲粮中添加BFA可增加盲肠咖啡因水平。咖啡因是一种存在于植物中的天然生物碱,已被鉴定为糖基水解酶18家族(几丁质酶)的抑制剂[26]。郭会林等[27]研究发现,咖啡因长期刺激可增加小鼠小肠绒毛高度、降低隐窝深度,并且增强小鼠肠道对肠道激素胰高血糖素样肽-1(GLP-1)和胰岛素的敏感性。另外,咖啡因可改善硫酸葡聚糖钠(DSS)诱导的小鼠结肠炎症状,减少细菌易位的发生和降低促炎细胞因子含量[28]。
4 结 论
饲粮中添加1 500 mg/kg BFA对仙居鸡生产性能有一定的改善作用,并且提高了空肠糜蛋白酶以及回肠糜蛋白酶、胰蛋白酶和淀粉酶活性,降低了血清促炎细胞因子IL-6和TNF-α含量,其良好的促生长和抗炎效果可能与盲肠中短链脂肪酸产生有关的罕见小球菌属、Catenibacillus和毛螺菌科-UGG-002相对丰度增加有关。BFA可作为饲用抗生素的潜在替代品。
猜你喜欢
今日农业(2022年13期)2022-09-15
中国典型病例大全(2022年7期)2022-04-22
江苏农业科学(2019年8期)2019-08-20
特别健康·下半月(2019年7期)2019-07-29
东坡赤壁诗词(2017年3期)2017-07-05
中外医学研究(2017年15期)2017-06-29
特别健康·下半月(2017年5期)2017-06-15
旅游纵览(2017年3期)2017-03-16
扬子江(2016年1期)2016-05-19
浙江人大(2016年5期)2016-05-14
