
青藏高原老芒麦不同部位的营养价值及其分子结构特性研究
2022-09-01 03:21孙晓旭史海涛陈有军蔡晓蕊李胜利
动物营养学报 2022年8期
彭 婧 孙晓旭 史海涛* 陈有军 蔡晓蕊 王 茜 柏 雪 李胜利
(1.西南民族大学畜牧兽医学院,青藏高原动物遗传资源保护与利用教育部重点实验室,成都 610041;2.西南民族大学青藏高原研究院,青藏高原生态畜牧业协同创新中心,成都 610041;3.中国农业大学动物科学技术学院,动物营养学国家重点实验室,北京 100193)
青藏高原地区是我国主要牧区之一,但其高寒、缺氧和强辐射的环境特点又使其与其他牧区的生产模式明显不同。当前,超载过牧、草原退化以及饲草料严重匮乏已成为制约青藏高原地区畜牧业转型升级的瓶颈因素。培育适应性强、产量高和营养价值良好的牧草对高原地区草原生态恢复和畜牧业可持续发展至关重要。
老芒麦是禾本科(Poaceae)、披碱草属(Elymus)的多年生草本植物,具有抗旱、耐寒、耐盐碱等优良特性,能够有效改善高原地区表层土壤理化性质[1-2]。由于其高产优质和对寒冷干旱气候的良好适应性[3],老芒麦已经成为青藏高原地区栽培利用最为广泛的当家草种之一[1]。饲草的不同部位其营养成分含量及消化利用率通常也不同[4]。研究老芒麦不同部位的营养价值,对高原地区牧草育种和家畜饲粮配制具有重要的指导意义。
获取饲料原料的营养价值参数是精准配制动物饲粮的前提。传统的化学分析需要较长的分析周期且严重依赖刺激性化学药品,在分析加工过程中通常会破坏饲料的内在结构[5]。饲料的营养价值不仅与其营养成分含量有关,还与其分子结构特性有关[5-6]。植物的不同品种及植株的不同部位都有其独特的分子结构特征,研究其分子结构特性并将其与营养价值参数进行关联分析,有助于从分子构造层面揭示营养价值差异原因并实现营养价值的快速、无损检测。傅里叶变换红外光谱(Fourier transform infrared spectroscopy,FTIR)技术以其需要样品量少、无需复杂的样品准备过程[6]以及直接、无损等优点[7],成为揭示饲料样品内在分子结构特征的有力工具[8]。通过使用该技术,Ortuo等[9]揭示了不同品种柳树叶和茎的分子结构信息。此外,玉米秸秆[8]、紫花苜蓿[6]等相关的研究也相继报道。目前,青藏高原地区老芒麦的营养价值参数尚不明确。基于此,本研究拟通过FTIR技术揭示老芒麦不同部位的分子结构信息,并结合常规化学分析和康奈尔净碳水化合物-蛋白质体系(CNCPS)技术比较不同部位老芒麦的营养价值,探求它们之间的相关关系,以期为老芒麦的育种和开发利用提供数据支撑。
1 材料与方法
1.1 样品采集和制备
试验所需的老芒麦采自西南民族大学青藏高原生态保护与畜牧业高科技研究示范基地(四川省阿坝藏族羌族自治州红原县,海拔3 504 m)牧草育种试验田。在老芒麦乳熟期,于2个试验小区随机确定1 m2的样方,留茬5 cm左右刈割,按照四分法进行缩样后,人工分离其不同部位(茎、叶、穗),并保留全株老芒麦样本做为对照。所有样品在烘箱中于65 ℃烘干至恒重,制备为风干样品。部分样品过1 mm筛粉碎,用于常规化学成分分析,同时以四分法取出部分样品过0.25 mm筛用于采集FTIR分析。
1.2 常规营养成分及CNCPS组分分析
干物质(dry matter,DM)、粗灰分(Ash)和粗脂肪(ether extract,EE)含量采用AOAC(2005)[10]的方法进行测定;中性洗涤纤维(neutral detergent fiber,NDF)、酸性洗涤纤维(acid detergent fiber,ADF)和酸性洗涤木质素(acid detergent lignin,ADL)含量按照Van Soest等[11]的方法进行测定,使用全自动纤维分析仪(Fibretherm FT12,德国)完成;测定NDF和ADF成分之后的残样用于测定中性洗涤不溶蛋白质(neutral detergent insoluble crude protein,NDICP)、酸性洗涤不溶蛋白质(acid detergent insoluble crude protein,ADICP)、非蛋白氮(non protein nitrogen,NPN)和可溶性蛋白质(soluble protein,SCP)的含量,测定按照Licitra等[12]的方法进行,使用杜马斯定氮仪(Dumatherm,德国)完成;淀粉(starch)含量采用爱尔兰Megazyme淀粉总量检测试剂盒(产品编号:K-TSTA;Megazyme,爱尔兰)进行测定[13]。最后根据测定值及CNCPS公式[14]得出各组分的含量;此外,因饲草中氨(PA1)的含量几乎为0[15],因此,本试验未测定CNCPS蛋白质亚组分中PA1的含量。
1.3 FTIR数据的采集及分析
使用FTIR仪(Nicolet iS50,Thermo Fisher Scientific Co.,美国)对样品进行光谱扫描,扫描波段800~4 000 cm-1,分辨率4 cm-1,扫描次数64次,每个样品重复采集6个图谱,读取并记录样品红外光谱图。利用OMNIC 8.2软件对扫描样品后获得的FTIR谱系进行处理分析。
1.4 数据处理与分析
采用SAS 9.4软件中的PROC GLM模块对营养成分、CNCPS组分及分子结构参数进行单因素方差分析,采用Duncan氏法进行多重比较,试验结果用平均值和均值标准误表示,P<0.05表示差异显著,0.05≤P<0.10表示差异趋于显著。根据Pearson相关分析法,利用SAS 9.4软件中的PROC CORR模块进行相关性分析。
2 结果与分析
2.1 老芒麦不同部位的营养成分及CNCPS组分
2.1.1 老芒麦不同部位的蛋白质成分和CNCPS蛋白质组分
老芒麦中茎、叶和穗的比例分别为59.8%、20.6%和19.6%(风干基础)。由表1可知,老芒麦不同部位之间的粗蛋白质(CP)、NDICP、ADICP、NPN和SCP含量均存在显著差异(P<0.05)。其中,叶的CP、NDICP和ADICP含量在老芒麦各部分中均为最高,显著高于全株、茎和穗(P<0.05);茎的CP、NDICP和ADICP含量在各部位中均为最低,且CP和NDICP含量显著低于全株、叶和穗(P<0.05),ADICP含量显著低于全株和叶(P<0.05);穗的SCP和NPN含量显著高于其他部位(P<0.05)。
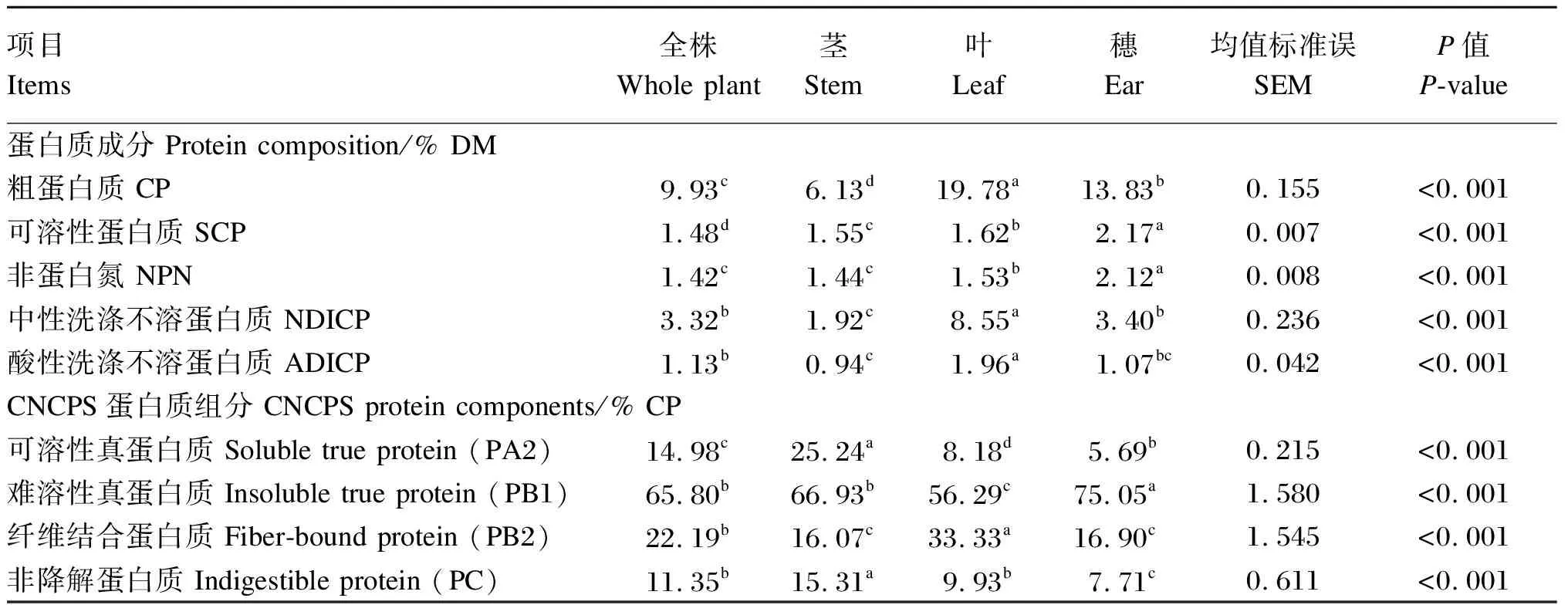
表1 老芒麦不同部位的蛋白质成分和CNCPS蛋白质组分
从CNCPS蛋白质组分来看,老芒麦不同部位之间的可溶性真蛋白质(PA2)、难溶性真蛋白质(PB1)、纤维结合蛋白质(PB2)和非降解蛋白质(PC)含量均存在显著差异(P<0.05)。其中,茎的PA2和PC含量显著高于全株、叶和穗(P<0.05),叶的PB1含量显著低于全株及其他部位(P<0.05),茎的PB2含量显著低于全株和叶(P<0.05)。
2.1.2 老芒麦不同部位的碳水化合物成分和CNCPS碳水化合物组分
由表2可知,老芒麦不同部位之间的NDF、ADF、ADL、淀粉、非纤维性碳水化合物(NFC)、半纤维素、纤维素以及碳水化合物含量均存在显著差异(P<0.05)。其中,茎的NDF、ADF、ADL、纤维素和碳水化合物含量最高,显著高于全株(ADL除外)、叶和穗(P<0.05);叶的NFC含量显著高于全株(P<0.05),NDF、淀粉和碳水化合物含量则显著低于其他部位(P<0.05);在淀粉和半纤维素含量方面,穗显著高于其他部位(P<0.05)。
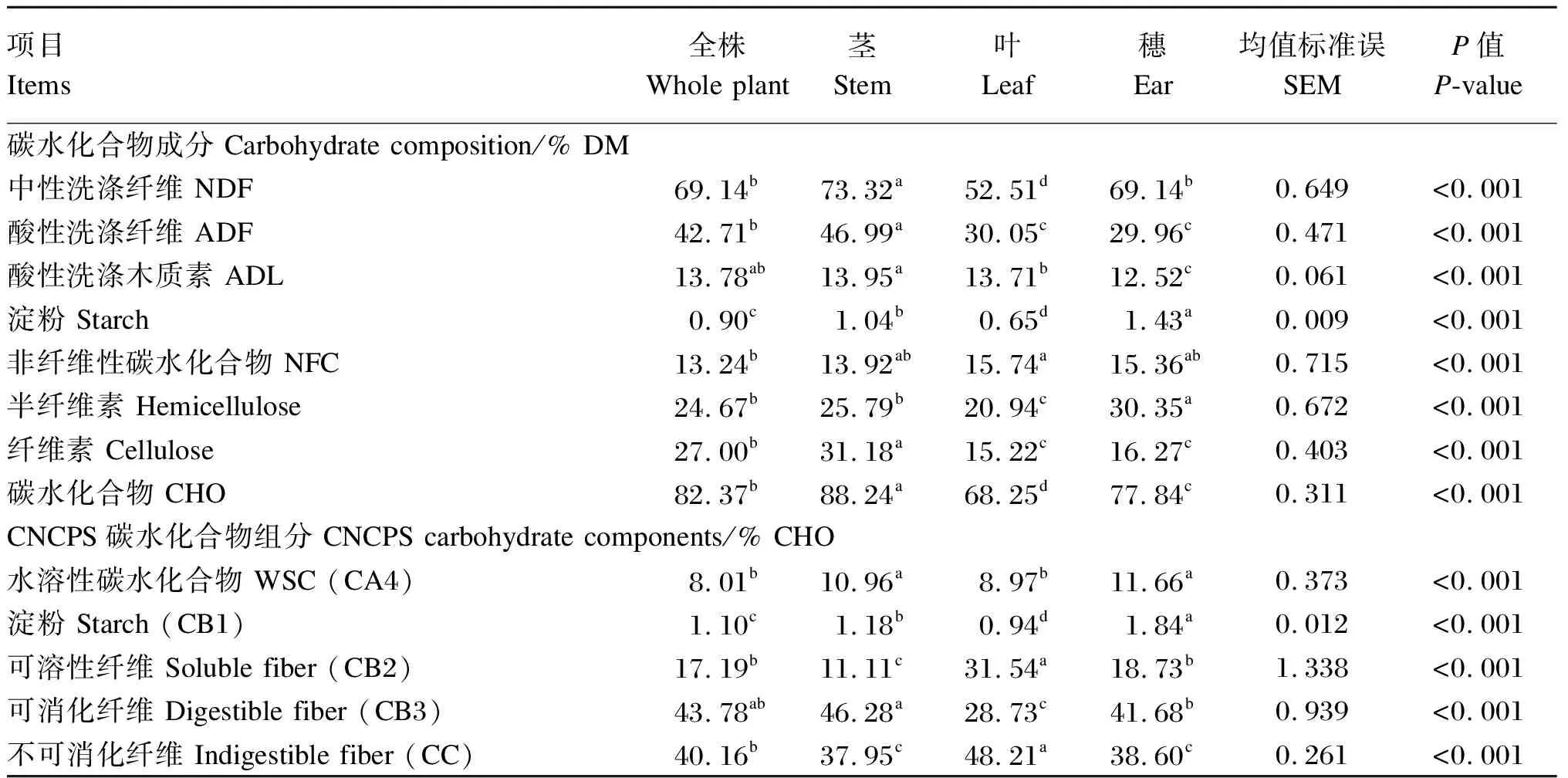
表2 老芒麦不同部位的碳水化合物成分和CNCPS碳水化合物组分
从CNCPS中的碳水化合物组分来看,老芒麦不同部位之间的水溶性碳水化合物(CA4)、淀粉(CB1)、可溶性纤维(CB2)、可消化纤维(CB3)和不可消化纤维(CC)含量均存在显著差异(P<0.05)。其中,穗的CA4含量显著高于全株和叶(P<0.05),CB1含量显著高于全株、茎和叶(P<0.05);叶的CB2含量显著高于其他部位(P<0.05),CB1和CB3含量则显著低于其他部位(P<0.05);茎的CB3含量在各部位中为最高,显著高于叶和穗(P<0.05);茎的CB2和CC含量在各部位中为最低,显著低于全株和叶(P<0.05)。
2.2 老芒麦不同部位的营养成分分子结构光谱参数
2.2.1 老芒麦不同部位的蛋白质分子结构光谱参数
由表3可知,老芒麦不同部位之间的蛋白质分子结构特征区域的部分峰参数存在显著差异(P<0.05)。从蛋白质酰胺带参数来看,叶的酰胺Ⅰ区峰高、酰胺Ⅱ区峰高和酰胺Ⅱ区峰面积均显著高于其他部位(P<0.05);穗的酰胺Ⅰ区峰面积显著高于其他部位(P<0.05);叶的酰胺Ⅰ区峰面积、酰胺Ⅰ区峰高/酰胺Ⅱ区峰高值及酰胺Ⅰ区峰面积/酰胺Ⅱ区峰面积值在各部位中最低。从蛋白质二级结构参数来看,叶的α-螺旋峰高及α-螺旋峰高/β-折叠峰高值均为最高,且α-螺旋峰高显著高于其他部位(P<0.05);全株的α-螺旋峰高、β-折叠峰高以及α-螺旋峰高/β-折叠峰高值均为最低,且α-螺旋峰高和α-螺旋峰高/β-折叠峰高值显著低于其他部位(P<0.05)。
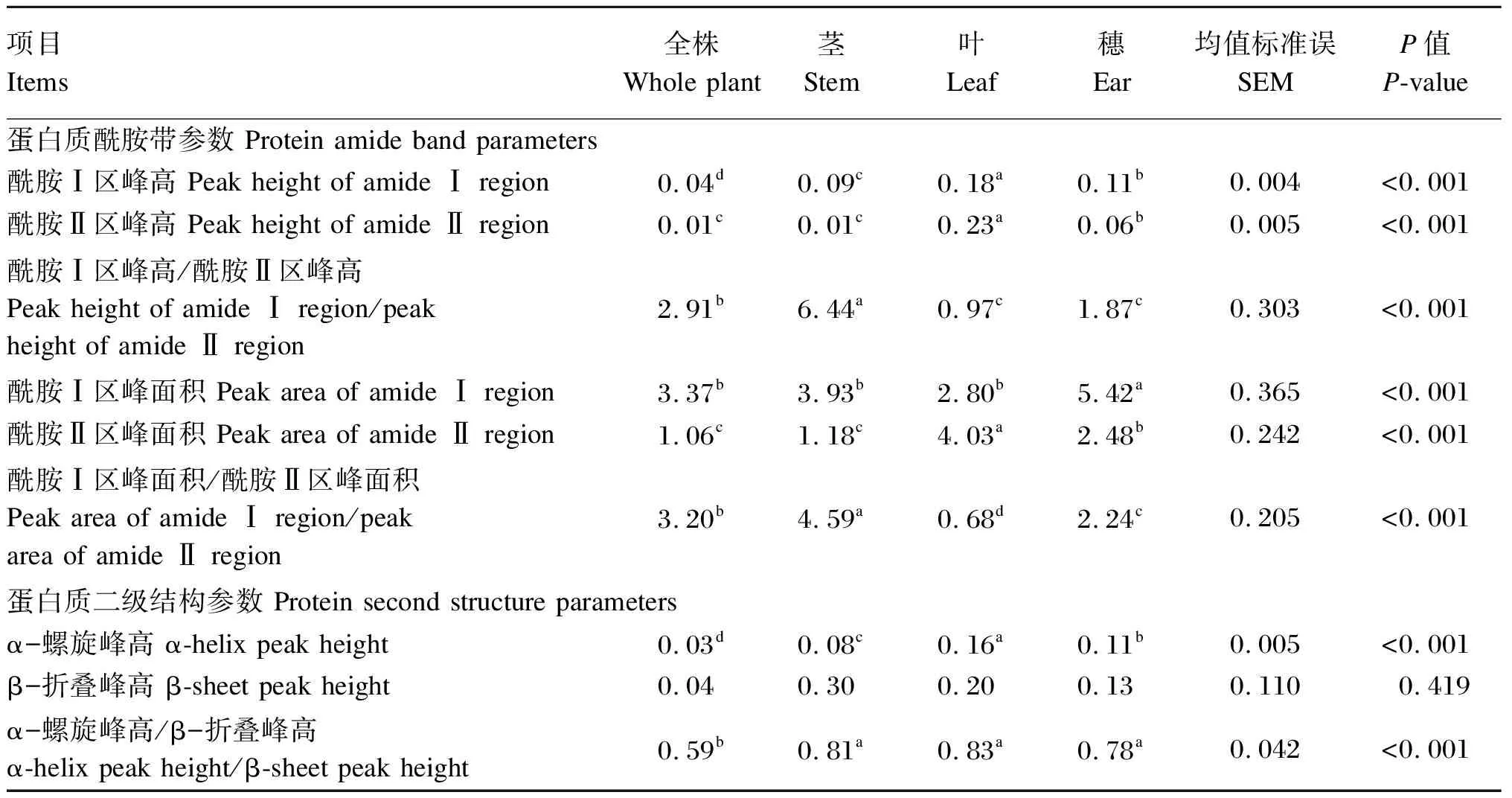
表3 老芒麦不同部位的蛋白质分子结构光谱参数
2.2.2 老芒麦不同部位的碳水化合物分子结构光谱参数
由表4可知,老芒麦不同部位之间的碳水化合物分子结构参数存在显著差异(P<0.05)。其中,全株的总碳水化合物亚峰Ⅰ峰高和总碳水化合物亚峰Ⅱ峰高显著高于其他部位(P<0.05);茎的结构性碳水化合物峰面积、结构性碳水化合物亚峰Ⅰ峰高、纤维素复合物峰面积、纤维素复合物峰高以及总碳水化合物亚峰Ⅲ峰高(全株除外)显著高于其他部位(P<0.05);叶的结构性碳水化合物亚峰Ⅱ峰高和结构性碳水化合物亚峰Ⅲ峰高显著高于其他部位(P<0.05);穗的结构性碳水化合物峰面积显著低于其他部位(P<0.05)。
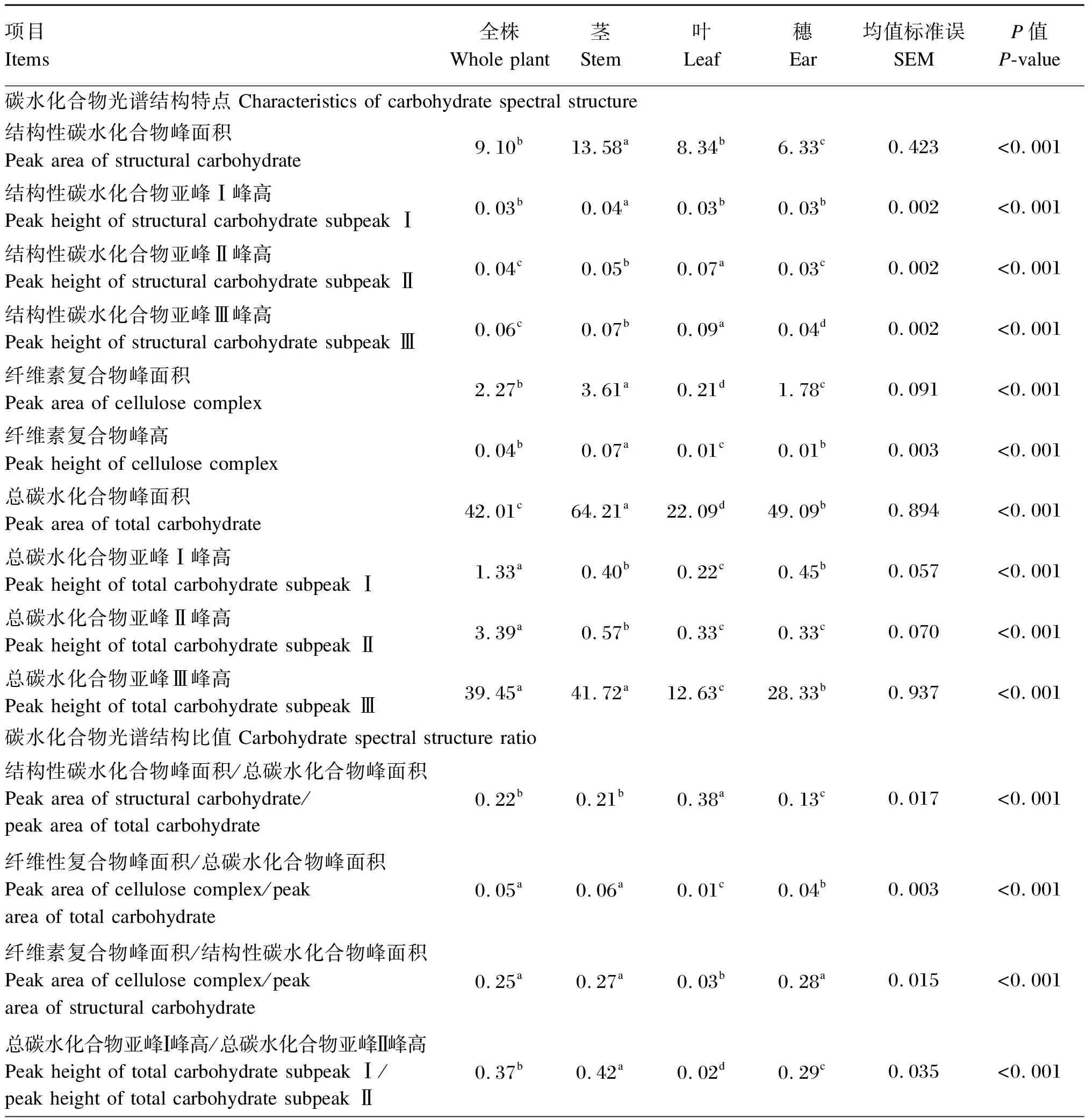
表4 老芒麦不同部位的碳水化合物分子结构光谱参数
老芒麦全株的总碳水化合物亚峰Ⅱ峰高/总碳水化合物亚峰Ⅲ峰高值最低,显著低于其他部位(P<0.05);茎的总碳水化合物亚峰Ⅰ峰高/总碳水化合物亚峰Ⅱ峰高和总碳水化合物亚峰Ⅰ峰高/总碳水化合物亚峰Ⅲ峰高值最高,显著高于其他部位(P<0.05);叶的纤维性复合物峰面积/总碳水化合物峰面积和纤维素复合物峰面积/结构性碳水化合物峰面积值显著低于全株、茎和穗(P<0.05);穗的结构性碳水化合物峰面积/总碳水化合物峰面积值最低,显著低于全株、茎和叶(P<0.05)。
2.3 老芒麦不同部位的营养价值与分子结构之间的相关关系
2.3.1 老芒麦不同部位的蛋白质分子结构与其蛋白质组分之间的相关关系
由表5可知,老芒麦不同部位的蛋白质分子结构光谱参数与蛋白质组分之间存在一定的相关关系。其中,在蛋白质成分中,CP、ADICP、NDICP含量与酰胺Ⅰ区峰高、酰胺Ⅱ区峰高、α-螺旋峰高及酰胺Ⅱ区峰面积呈极显著正相关(r=0.71~0.95,P<0.01);SCP和NPN含量与酰胺Ⅰ区峰面积呈极显著正相关(r=0.72~0.73,P<0.01)。

表5 老芒麦不同部位的蛋白质分子结构与其蛋白质组分之间的相关关系
在CNCPS蛋白质组分中,PA2含量与酰胺Ⅱ区峰高、酰胺Ⅰ区峰高/酰胺Ⅱ区峰高值和酰胺Ⅰ区峰面积/酰胺Ⅱ区峰面积值呈极显著正相关(r=0.81~0.94,P<0.01),与酰胺Ⅰ区峰高和酰胺Ⅱ区峰面积呈显著负相关(r=-0.77~-0.58,P<0.05);PB1含量与α-螺旋峰高/β-折叠峰高值和酰胺Ⅰ区峰面积/酰胺Ⅱ区峰面积值呈显著正相关(r=0.60~0.76,P<0.05);PB2含量与酰胺Ⅱ区峰高、酰胺Ⅰ区峰面积和α-螺旋峰高呈显著负相关(r=-0.70~-0.53,P<0.05)。
2.3.2 老芒麦不同部位的碳水化合物分子结构与其碳水化合物组分之间的相关关系
由表6可知,老芒麦不同部位的碳水化合物分子结构光谱参数与碳水化合物组分之间存在一定的相关关系。其中,在碳水化合物成分中,NDF含量与纤维素复合物峰面积、总碳水化合物峰面积及纤维性复合物峰面积/总碳水化合物峰面积值呈极显著正相关(r=0.88~0.95,P<0.01);ADF含量与结构性碳水化合物峰面积、纤维素复合物峰面积及纤维性复合物峰面积/总碳水化合物峰面积值呈极显著正相关(r=0.69~0.82,P<0.01);ADL含量与结构性碳水化合物峰面积呈极显著正相关(r=0.84,P<0.01);淀粉和半纤维素含量与结构性碳水化合物峰面积/总碳水化合物峰面积值呈极显著负相关(r=-0.84~-0.83,P<0.01);纤维素和碳水化合物含量与纤维素复合物峰面积、总碳水化合物峰面积及纤维性复合物峰面积/总碳水化合物峰面积呈极显著正相关(r=0.71~0.96,P<0.01)。
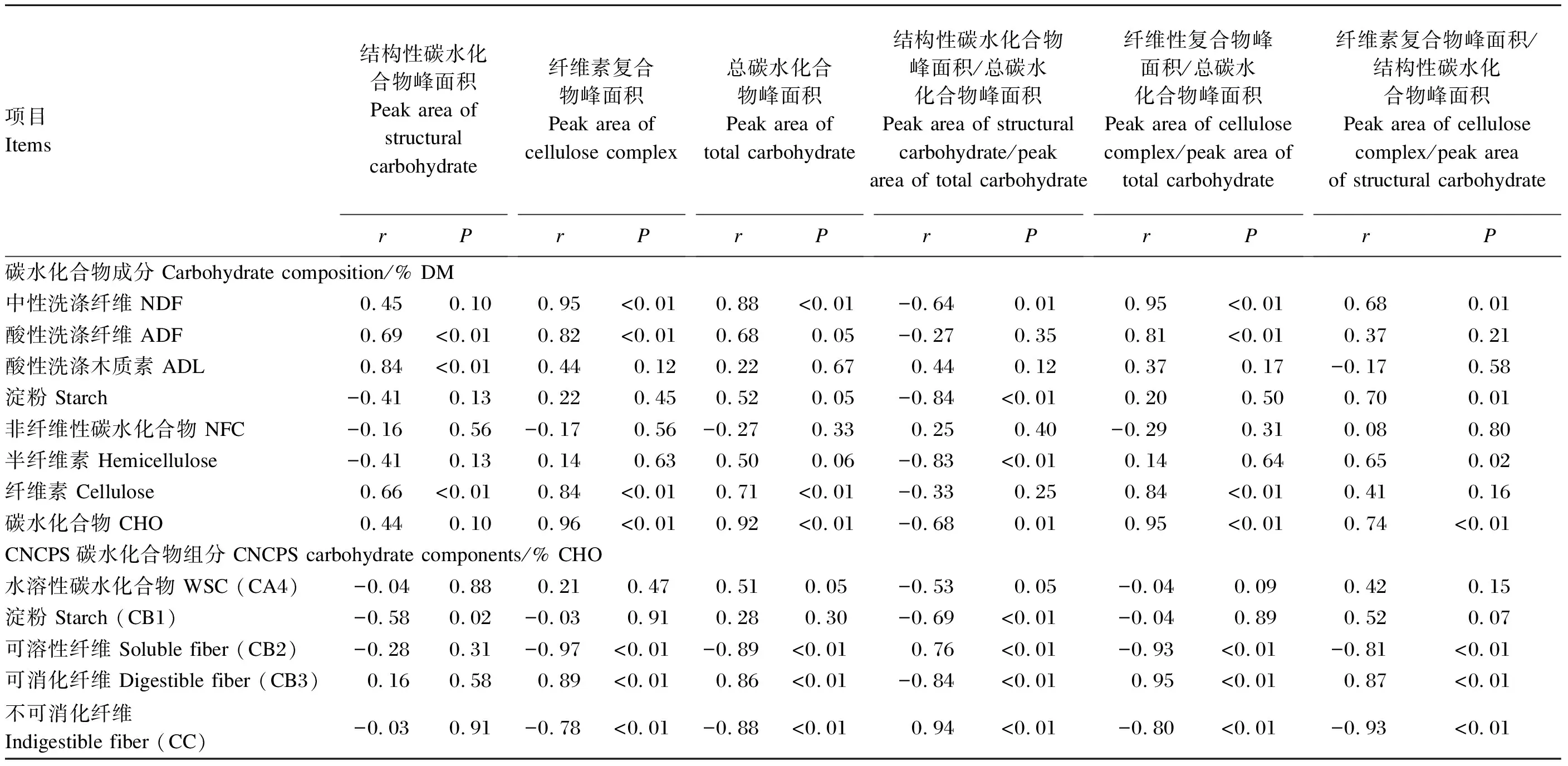
表6 老芒麦不同部位的碳水化合物分子结构与其碳水化合物组分之间的相关关系
在CNCPS碳水化合物组分中,CB1含量与结构性碳水化合物峰面积/总碳水化合物峰面积呈极显著负相关(r=-0.69,P<0.01);CB2、CC含量与结构性碳水化合物峰面积/总碳水化合物峰面积值呈极显著正相关(r=0.76~0.94,P<0.01),与纤维素复合物峰面积、总碳水化合物峰面积、纤维性复合物峰面积/总碳水化合物峰面积值及纤维素复合物峰面积/结构性碳水化合物峰面积值呈极显著负相关(r=-0.97~-0.78,P<0.01);CB3含量与纤维素复合物峰面积、总碳水化合物峰面积、纤维性复合物峰面积/总碳水化合物峰面积值及纤维素复合物峰面积/结构性碳水化合物峰面积值呈极显著正相关(r=0.86~0.95,P<0.01),与结构性碳水化合物峰面积/总碳水化合物峰面积呈极显著负相关(r=-0.84,P<0.01)。
3 讨 论
3.1 老芒麦不同部位的营养成分及CNCPS组分
3.1.1 老芒麦不同部位的蛋白质成分和CNCPS蛋白质组分
饲料中的营养成分可以在一定程度上反映饲料的营养价值。粗饲料CP含量高且NDF和ADF含量较低时,其营养价值通常较高[16]。Hao等[17]研究了构树不同部位(叶、茎和全株)的化学组成及瘤胃降解特性,结果表明,叶的CP、水溶性碳水化合物(WSC)、Ash、钙和磷含量最高,茎的NDF、ADF和ADL含量最高。Ortuo等[9]测定了不同品种柳树的叶和茎的化学成分,与茎相比,叶的纤维化合物的强度较低,CP含量较高。闫贵龙等[18]研究发现,与茎皮、苞叶等部位相比,玉米叶片的CP含量最高。以上结果与本试验得到的结果相一致。老芒麦叶的CP含量最高,符合植物叶片中蛋白质含量通常高于茎秆的规律。
新版本CNCPS体系(V 6.5)将蛋白质划分为PA1、PA2、PB1、PB2和PC[15]。其中,PA2是指在10 h内在瘤胃内迅速降解的蛋白质部分[15],PB1、PB2是粗饲料中主要的蛋白质来源,PB2含量高意味着更多的CP将通过瘤胃;而PC含量越高,说明其蛋白质在瘤胃内消化率越低,提供的蛋白质质量越差[14,18]。本试验结果显示,老芒麦叶的PB2含量高于其他部位,茎的PC含量最高,说明老芒麦叶片在瘤胃内的发酵速率可能更快,蛋白质品质较好,可以更好地被反刍动物利用,而纤维性物质含量较高的茎秆部分更难被动物消化降解。
3.1.2 老芒麦不同部位的碳水化合物成分和CNCPS碳水化合物组分
饲草中NDF、ADF含量是反映牧草营养价值高低的重要指标。NDF含量越高饲料的适口性越差;ADF含量越高,其在家畜体内的消化率越低[19]。ADL是难以被动物消化利用的成分,与消化率成负相关,同时还会降低其他营养物质的消化利用。茎作为整株植物的支撑,在植物生长过程中被木质化[17]。本试验结果显示,老芒麦茎的NDF、ADF和ADL含量高于其他部位;叶的NFC含量较高,而NDF、ADF、ADL、半纤维素和纤维素等纤维化合物含量较低。茎的高纤维含量可能会影响动物的采食量和养分的消化率[20]。
在CNCPS体系中,CA4代表糖类,CB1代表淀粉,CB2代表可溶性纤维部分,CB3代表可利用的NDF部分,CC代表着不可利用的碳水化合物部分,这些亚组分在瘤胃中以不同的速率降解[15]。本试验中,与叶相比,茎的CA4含量较高,但CB1+CB2含量较低,说明茎中糖类含量较高,而淀粉和可利用的细胞壁含量较低。
3.2 老芒麦不同部位的营养成分分子结构光谱参数
3.2.1 蛋白质分子结构光谱参数
研究表明,动物饲料中CP的溶解度、消化率等不仅受CP化学组成的影响,还受蛋白质内在分子结构的影响[15]。研究蛋白质的分子结构特性有助于深入评估其在动物体内的可消化性[21]。酰胺Ⅰ区和酰胺Ⅱ区是与蛋白质二级结构相关的2个主要条带[22],通常使用该指标进行蛋白质结构方面的预测[23]。酰胺Ⅰ区和Ⅱ区的峰高、峰面积之间的差异反映了官能团的数量和CP含量的差异[24]。本试验中,老芒麦不同部位的酰胺Ⅰ区峰高、酰胺Ⅰ区峰面积存在较大差异,叶的酰胺Ⅰ区峰高最高,穗的酰胺Ⅰ区峰面积最高,全株的酰胺Ⅰ区峰高、酰胺Ⅰ区峰面积较低,这与化学方法测得的CP含量趋势一致。
饲料中蛋白质在瘤胃内的降解特性与蛋白质α-螺旋和β-折叠参数有关[25]。α-螺旋/β-折叠值越大,通常表明蛋白质的质量更好[26]。Doiron等[27]研究表明,饲料α-螺旋/β-折叠值的变化可反映其真可吸收CP的供应量。Yu等[21]研究表明,较低的CP利用率可能与β-折叠比例较高有关。本试验中,叶的α-螺旋峰高显著高于其他部位,说明叶片蛋白质利用率可能更高,这与叶的PB2含量最高的结果相一致。
3.2.2 碳水化合物分子结构光谱参数
饲料碳水化合物组成及其分子结构特性均与营养价值密切相关[28]。分子结构的差异在理论上会造成饲料原料营养价值的不同[29],进而影响饲料养分在瘤胃内的降解率[21]以及小肠内的吸收率[30]。结构性碳水化合物区域的3个特征峰高主要与半纤维素和纤维素化合物有关[31]。本试验研究表明,茎的结构性碳水化合物峰面积以及结构性碳水化合物中3个亚峰的峰高均较高,这与其纤维素和半纤维素的含量结果相吻合。
纤维素复合物峰面积和峰高通常与植物中的纤维素含量有关[32],分子结构上的差异会影响饲料在瘤胃中的降解特性[33]。在总碳水化合物区域内,总碳水化合物峰面积越大,其碳水化合物、淀粉的含量通常越高[34]。本研究中,茎的纤维素复合物峰面积显著高于其他部位,这与常规化学成分中NDF和ADF含量在茎中较高一致。该结论也与Ortuo等[9]研究结果一致。
3.3 老芒麦不同部位的营养价值与分子结构之间的相关关系
3.3.1 蛋白质分子结构与其蛋白质组分之间的相关关系
研究表明,蛋白质分子结构与其营养价值之间存在紧密的联系,可以借助蛋白质分子结构特性对饲料蛋白质营养价值进行估测[33]。本研究中,CP含量与酰胺Ⅰ区峰高、酰胺Ⅱ区峰高及酰胺Ⅱ区峰面积存在显著正相关。Xin等[26]和Ismael等[35]在关于生物燃料副产品、菜籽粕混合颗粒的研究中也得出相似结论。Doiron等[27]和Samadi等[36]在研究不同温度热处理对亚麻籽、油菜籽蛋白质分子结构的影响时发现,α-螺旋/β-折叠值与NDICP、ADICP含量呈显著正相关,而本试验中α-螺旋峰高/β-折叠峰高值与蛋白质化学成分之间并不存在相关性。Huang等[7]研究发现,双低菜籽粕的PA2含量与酰胺Ⅰ 区峰高/酰胺Ⅱ区峰高值呈正相关,而α-螺旋峰高/β-折叠峰高值与蛋白质亚组分之间没有相关性。上述结果与本试验研究结果一致。然而,Gomaa等[37]在研究埃塞俄比亚芥籽粕和油菜籽粕时发现,PA2含量与酰胺Ⅰ区峰高、酰胺Ⅰ区峰面积和α-螺旋峰高呈正相关。这些差异可能与样品的种类、加工方式、成分等因素有关。
3.3.2 碳水化合物分子结构与其碳水化合物组分之间的相关关系
碳水化合物的分子结构与碳水化合物组分构成及含量有关,相应的峰值或峰面积可以作为预测饲料原料营养价值的参数[38]。Xin等[26]在研究豆粕、麦麸和玉米酒糟等饲料时发现,纤维素复合物峰面积与NDF含量呈正相关,总碳水化合物峰面积与CC含量呈负相关。Xin等[8]在研究玉米秸秆时发现半纤维素含量与结构性碳水化合物峰面积存在显著负相关,CB2含量与纤维素复合物峰面积存在显著负相关。以上结果均与本试验结果一致。Ji等[31]在研究不同来源牧草时发现,CB3含量与纤维素复合物峰面积、结构性碳水化合物峰面积呈负相关;Chen等[39]在研究干酒糟及其可溶物(DDGS)副产品时发现,光谱参数与碳水化合物组分之间没有相关性,以上结果与本试验结果不一致,导致这种差异的原因可能是由于饲料种类不同等因素导致的。
4 结 论
① 高原老芒麦对草食家畜具有良好的营养价值,老芒麦不同部位的营养价值存在显著差异。其中,叶的CP、NDICP和NFC含量最高,穗的NPN、SCP、淀粉和半纤维素含量最高,茎的NDF、ADF、ADL、纤维素及碳水化合物含量最高。而CNCPS中的蛋白质和碳水化合物组分在不同部位也存在明显不同。从营养成分含量来看,老芒麦叶的营养价值较高,茎的营养价值相对较低。
② 老芒麦不同部位的分子结构参数存在差异且与其营养特性之间存在较强的相关关系,利用FTIR技术可以快速、无损地评定青藏高原老芒麦的营养价值。
猜你喜欢
分子催化(2022年1期)2022-11-02
农产品质量与安全(2022年4期)2022-08-24
医学概论(2022年4期)2022-04-24
毛纺科技(2022年3期)2022-04-07
小猕猴智力画刊(2021年11期)2021-11-28
疯狂英语·读写版(2020年11期)2020-12-21
中学生数理化(高中版.高考理化)(2019年6期)2019-06-22
中学化学(2016年10期)2017-01-07
中国中药杂志(2016年6期)2016-05-11
农村百事通(2004年12期)2004-08-24
