
镉(Cd)胁迫下蓖麻蛋白质组学筛查及RcBSK7抗性功能研究
2023-01-11 02:57赵慧博赵志强包春光李茹鑫黄凤兰
植物研究 2023年1期
赵慧博 赵志强 包春光 温 琦 李茹鑫 黄凤兰
(1. 内蒙古民族大学生命科学与食品学院,通辽 028000;2. 内蒙古民族大学农学院,通辽 028000;3. 通辽市农畜产品质量安全中心,通辽 028000;4. 蓖麻育种国家民委重点实验室,通辽 028000;5. 内蒙古自治区高校蓖麻产业工程技术研究中心,通辽 028000;6.内蒙古自治区蓖麻育种重点实验室,通辽 028000)
全国土壤污染状况调查表明,重金属无机污染物超标点位数占全部超标点位的82.8%,其中重金属镉(Cd)点位超标率约7.0%,成为最高无机污染物,这引起人们对土壤中重金属Cd 污染更广泛的关注[1]。植物长期生长在Cd 胁迫下,会加速细胞内活性氧(Reactive oxygen species,ROS)的生成从而破坏氧化系统的稳态[2-3];通过抑制叶绿素合成、破坏光反应和电子传递链等非气孔限制因素以及影响气孔导度、抑制碳固定等途径影响植物的光合作用[4-5];此外,Cd2+可以与植物细胞核相互作用,并通过影响DNA 碱基修饰等过程对DNA 产生损伤,这可能也是造成其蛋白质合成受阻的直接原因。
土壤重金属具有隐蔽性、长期性和不可逆性的特点,重金属污染的治理是一个世界性的难题,目前,重金属的防治主要有物理修复方法、化学修复方法和生物修复方法。其中生物修复具有投资少、效率高、不给环境造成二次污染的特点,该修复方法是一项既环保又便宜的“绿色”技术,具有巨大的潜力[6]。
蓖麻(Ricinus communis)为大戟科(Euphorbiaceae)蓖麻属(Ricinus)一年或多年生草本植物。由于蓖麻枝叶茂盛、茎秆粗壮、根系发达,能防风固沙,因此被广泛种植在高盐碱等环境瘠薄地区[7],同时蓖麻也因其抗逆性强、适应性广等特点而被称为理想的绿色环保植物[8]。研究发现[9],蓖麻对重金属Cd 具有较强的吸附性,同时非食用植物蓖麻具有对铅(Pd)、锌(Zn)等多种重金属的超富集转运系数和修复能力,因此被称为新型重金属富集植物并广泛应用于重金属污染土壤修复中。
近年来,随着植物基因组测序技术的推动发展,转录组学、差异蛋白质组学和比较代谢组学等多种高通量组学技术传播迅速并广泛应用于植物响应重金属胁迫的分子机制研究中,这对多角度阐明植物对重金属的耐受和解毒机制具有重要的意义。鉴于此,本研究通过对Cd 胁迫下蓖麻根系进行差异蛋白质组学分析,揭示不同浓度Cd 胁迫对蓖麻生长发育的影响以及蓖麻对重金属Cd 耐受性的分子机制;同时筛选出差异显著基因在拟南芥(Arabidopsis thaliana)中完成异源表达以验证其基因功能,为蓖麻的耐Cd 基因鉴定以及土壤中重金属Cd污染修复奠定良好的基础。
1 材料与方法
1.1 植物材料处理
植物材料:“2129”优良蓖麻品系和哥伦比亚野生型拟南芥(Columbia,Col-0),由内蒙古自治区高校蓖麻产业工程技术研究中心提供;拟南芥缺失突变体atbsk7(SALK_077982C)购自AraShare 网站(http://www.arashare.cn/)。
筛选过程:将蓖麻种子置于蛭石内,分别浇入700 mL 水溶液和Cd 溶液(0~4 000 mg·L-1),28 ℃培养72 h后,观察记录蓖麻种子的发芽状态,对Cd浓度进行初步筛选;根据初筛结果,将萌发的蓖麻种子移栽至白沙内,分别加以Cd溶液(0~1 000 mg·L-1),持续生长21 d 后,观察记录蓖麻植株的发育状态。本试验中所用沙土均为标准1 mm 无粉尘白沙,研究过程中所产生的Cd 污染物均按照环保部门要求进行处理、回收。
材料获取:将蓖麻种子种植于白沙内,无Cd 条件下正常培养生长14 d 后,随机分配幼苗并分别以0、300、700、1 000 mg·L-1的Cd 溶液(CdCl2·5H2O)处理,持续生长21 d 后,分别取植株根、茎、叶部经液氮超低温冷冻后于冰箱-80 ℃保存,用于重金属浓度测定和差异蛋白质组学分析,分别记作CK、ZA、ZB、ZC。每个处理取3个生物学重复,每个重复由3株样株混合获得。
对比策略:300、700、1 000 mg·L-1的Cd溶液处理蓖麻植株根部分别与水处理对照植株根部比对,分别记作:ZA_VS_CK,ZB_VS_CK,ZC_VS_CK。
1.2 Cd含量测定
重金属Cd 含量测定由青岛科创质量检测有限公司完成,具体方法如下:称取1.0 g样品经液氮研磨至粉末后置于聚四氟乙烯消解罐内,加入5 mL硝酸,待反应结束后,放入微波消解仪(MILESTONE ETHOS1,意大利)内消解;待温度降至50 ℃以下时,超纯水润洗3~4 次后经电感耦合等离子质谱仪(Thermo iCAPQ,美国)测定,每个处理取3个生物学重复,每个重复由3株样本混样获得。
1.3 蛋白质组学
利用酚提取法提取蓖麻材料根部的总蛋白质,经胰酶酶解后,根据iTRAQ 试剂盒(AB SCIEX,美国)操作说明标记肽段,被标记肽段被加载至Agilent 300Extend C18 色 谱 柱(4.6 mm×250 mm,5 μm)中经液相色谱流动相A(2%乙腈;0.1%甲酸)溶解后进入nanoACQUITY UPLC M-Class system(Waters,美国)超高效液相系统进行分离。经超高效液相系统分离后的肽段被注入NSI 离子源中进行电离后,进入Q Exactive HF质谱进行分析。
经质谱鉴定得到的差异蛋白质通过UniProt蓖麻蛋白质数据库进行检索,利用Proteome Discoverer 2.2 软件进行数据库检索、谱肽、蛋白质定量。将每个蛋白质在2 个比较对样品中的相对定量值进行t-test 检验,以P≤0.05 作为显著性指标,并使用R 语言进行聚类分析,同时通过对所有差异蛋白质向Gene Ontology 数据库(http://www.geneontology.org/)的各个term 映射,找出与所有蛋白质背景相比,在差异蛋白质中显著富集的GO条目。
1.4 RT-qPCR验证
在对差异蛋白质组学分析的基础上,选取9个差异蛋白质进行荧光定量PCR(RT-qPCR)分析,研究其在转录水平上的表达模式,其中以Actin(LOC8273907)作为内参对基因进行归一化。使用GoTaq®qPCR and RT-qPCR Systems(普洛麦格,北京)进行RT-qPCR,每个处理3 个生物学重复。Ct 值取平均值,采用2-ΔΔCt法计算获得RT-qPCR 结果(引物序列见表1)。

表1 RT-qPCR引物序列Table 1 RT-qPCR primer sequence
1.5 异源表达
选择在蛋白质组学以及RT-qPCR 中均显著上调的基因,对其进行参与Cd胁迫响应的功能验证。通过在NCBI进行blastp 搜索得知它编码了一个丝氨酸/苏氨酸蛋白激酶,本研究中命名为RcBSK7。
利用CaMV35s 启动子将基因RcBSK7的CDS序列克隆至pCAMBIA1305.2 载体,并将重组表达载体pCAMBIA1305.2-RcBSK7转化至GV3101 农杆菌(Agrobacterium)感受态。通过浸花法将重组农杆菌菌液转入拟南芥(野生型Col-0 系及突变体atbsk7),通过50 mg·L-1潮霉素抗性筛选分别获得过表达抗性植株RcBSK7-OE 和回补表达抗性植株RcBSK7-CE(引物序列见表2)。

表2 目的基因PCR引物序列Table 2 Primer sequences of double primers PCR
1.6 拟南芥抗性植株分子水平鉴定
使用M5 超光速mix 试剂盒(聚合酶,北京)对拟南芥抗性植株进行DNA 水平鉴定,最终确定过表达和回补表达拟南芥阳性植株。
1.7 过表达拟南芥抗性植株的生物学水平鉴定
以野生型和缺失突变体拟南芥植株作为对照,对过表达拟南芥植株进行Cd 耐受性分析。取适量种子经消毒清洗后点种于MS 固体培养基,在4 ℃春化72 h 后,移至16 h/8 h 光周期、23 ℃/20 ℃昼/夜循环的环境下培养14 d,分别移至Cd 浓 度 为0、50、75 μmol·L-1的MS 固体培养基内培养10 d,培养条件同上。使用10 X 智能S生 物 显 微 镜(DM6,德 国)及LAS X 软 件(v3.0.16120.2)对拟南芥植株根系及叶片生长状态进行观察统计,每组数据取5 个生物学重复的平均值。
(1)灌浆料的本构模型可参考混凝土的本构模型进行模拟,经过对比分析,试验结果与数值分析结果吻合较好,验证了模型参数选取的合理性。
2 结果与分析
2.1 重金属Cd溶液浓度筛选
为了研究Cd 胁迫对蓖麻生长发育的影响,筛选观察不同浓度Cd 胁迫对蓖麻种子发芽及蓖麻植株生长状态(见图1~2,表3)。根据筛选试验,选取0、300、700、1 000 mg·L-1Cd 处理后的蓖麻植株作为后续研究材料(见图3)。

图1 0~4 000 mg·L-1重金属Cd处理蓖麻种子发芽结果Fig.1 Germination results of castor bean seeds treated with 0-4 000 mg·L-1 heavy metal Cd concentration

图3 不同剂量Cd胁迫下蓖麻植株及根系生长状态A~D.0、300、700、1 000 mg·L-1的Cd处理完整植株Fig.3 Growth status of castor plant and root system under different doses of Cd stress A-D.Intact plants under 0,300,700,1 000 mg·L-1 Cd treatment

表3 0~1 000 mg·L-1重金属Cd处理蓖麻植株生长情况测定结果Table 3 0-1 000 mg·L-1 heavy metal Cd concentration treatment of castor plant growth status results
2.2 蓖麻植株Cd含量分析
由Cd 含量测定结果可知(见图4),随着Cd 胁迫浓度的增加,蓖麻根部、茎部以及叶部检测到的Cd浓度亦随之增加。其中,重金属Cd在蓖麻根部显著积累,Cd 含量分别为茎和叶部的25.6、33.5倍。由此推测,植物在Cd 胁迫下,作为土壤的第一接触器官的根系可能为Cd 富集的主要部位,并通过茎—芽转运至茎部以及叶部,此结果与John等[10]的研究一致。结合以上结果,选取蓖麻根部作为后续蛋白质组学研究材料。

图4 不同质量浓度重金属Cd处理蓖麻植株Cd含量测定结果*0.01<P≤0.05;**0<P≤0.01Fig.4 Cd content determination results of castor plants treated with different heavy metal Cd concentrations*0.01<P≤0.05;**0<P≤0.01

图2 0~1 000 mg·L-1重金属Cd处理蓖麻植株生长状态A~L分别为0~1 000 mg·L-1的Cd胁迫下蓖麻植株的生长状态Fig.2 The growth status of castor plant plants treated with 0-1 000 mg·L-1 heavy metal Cd concentration A-L are respectively 0-1 000 mg·L-1 of the growth status of castor plant under Cd stress
2.3 差异蛋白质的功能分析
基于同位素标记相对定量(iTRAQ)技术对不同Cd 胁迫处理的蓖麻植株根部进行蛋白质组学分析,共检测到217个、156种蛋白质点差异表达显著(Ratio≥1.2,P≤0.01),其中75 个差异蛋白质表达量显著下调,142个差异蛋白质表达量显著上调。
将上述差异蛋白质进行功能分类(见图5),将156种差异蛋白质分为7个类别:其中参与胁迫/防御/解毒的蛋白质数量最多,约占总蛋白质数量的34%;其次是参与能量与碳代谢的蛋白质,约为总蛋白质的21%;而参与蛋白质折叠与降解、DNA 损伤修复、氨基酸代谢等蛋白质分别约为总蛋白质的14%、4%、3%等。
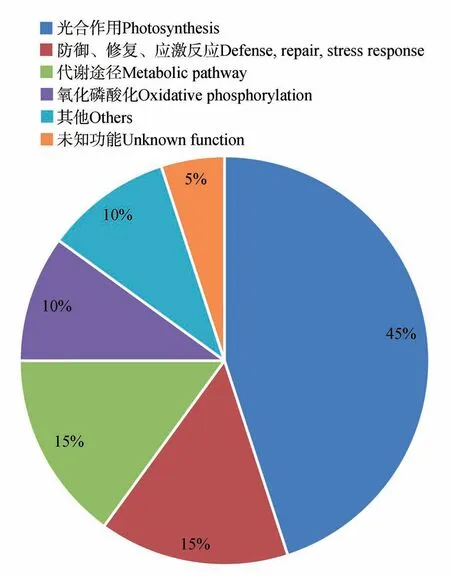
图5 差异蛋白质功能分类Fig.5 Differential protein function classification
2.4 差异蛋白质聚类分析
对各组差异蛋白质聚类分析结果如下(见图6):300 mg·L-1的Cd 胁迫下,与防御解毒和能量碳代谢相关的蛋白质上调显著;700 mg·L-1的Cd 胁迫下,除较多营养贮存相关的蛋白质表达量显著上调外,参与叶绿素合成、CO2固定以及某些参与非生物胁迫的膜转运相关蛋白质的表达量均呈现不同程度的上调;1 000 mg·L-1的Cd 胁迫下,少数参与细胞程序性死亡及急性创伤诱导相关蛋白质表达量特异性上调,这是在其他两组比对中T 未观察到的结果。



图6 差异蛋白质层次聚类分析A.ZA_VS_CK 差异蛋白质聚类分析;B.ZB_VS_CK差异蛋白质聚类分析;C.ZC_VS_CK差异蛋白质聚类分析;列表示分组;行表示差异蛋白质Fig.6 Differential protein cluster analysis A. ZA_VS_CK differential protein cluster analysis;B. ZB_VS_CK differential protein cluster analysis;C. ZC_VS_CK differential protein cluster analysis;Column represents grouping;Row represents the difference protein
2.5 差异蛋白质GO功能注释
对各组差异蛋白质进行GO功能注释。由图7可知,GO 功能注释共分生物过程、细胞成分、分子功能3 大类。其中,各组中参与细胞呼吸、能量代谢等生物过程的差异蛋白质比例较大;其次分别是参与细胞骨架、细胞内非膜细胞器等细胞组成成分,以及ATP 结合蛋白、K+通道蛋白等参与分子功能相关的差异蛋白质。以上结果与上述差异蛋白质层次聚类分析一致,即各差异蛋白质与生物学功能保持一定的相关性。

图7 差异蛋白质GO功能注释分析A.ZA_VS_CK差异蛋白质GO注释;B.ZB_VS_CK差异蛋白质GO注释;C.ZC_VS_CK差异蛋白质GO注释Fig.7 Differential protein GO function annotation analysis A. ZA_VS_CK differential protein GO function annotation analysis;B. ZB_VS_CK differential protein GO function annotation analysis;C.ZC_VS_CK differential protein GO function annotation analysis
2.6 差异蛋白质的RT-qPCR分析
在以上分析结果的基础上,选取差异显著且可能参与Cd 胁迫响应的蛋白质进行RT-qPCR 分析,研究其在转录水平的表达模式(见图8)。结果发现响应Cd 胁迫的各差异蛋白质在转录水平也呈现出相同的表达模式,即可以从各差异蛋白质编码基因出发,在转录水平研究蓖麻根系对Cd 胁迫的响应机制。其中,ZA 中B9RCB9和B9RGI8分别在转录水平显示出更显著的上调和下调趋势;而B9T1G7 在ZA、ZB 中均显示出较高的上调表达量,且结果与蛋白质组学分析结果高度一致,故编码以上蛋白质的基因引起高度的注意。

图8 蓖麻植株响应Cd胁迫的部分差异蛋白质RT-qPCR检测结果Fig.8 RT-qPCR detection results of some differential proteins in castor plants in response to Cd stress
2.7 拟南芥抗性植株鉴定
为了分析基因RcBSK7在蓖麻响应Cd 胁迫研究中的作用,以WT 及突变体为对照,对RcBSK7-OE 进行Cd 耐受性分析(见图9~10)。在无Cd 处理下,Col-0、atbsk7和RcBSK7-OE 的根系、叶片均发育良好。在50 μmol·L-1Cd 处理下,Col-0 和atbsk7根系的相对伸长长度明显低于RcBSK7-OE,除此之外,Col-0 和atbsk7的叶片生长受到较明显抑制,其中atbsk7个别植株叶片出现较明显的黄化、枯萎等现象。在75 μmol·L-1Cd 处理下,比对观察发现Col-0、atbsk7、RcBSK7-OE 在受损伤程度加深的基础上,表现出同50 μmol·L-1Cd 胁迫相似的响应趋势,即RcBSK7-OE 对Cd 的承受能力更强、更稳定。

图9 Col-1、atbsk7、RcBSK7-OE对重金属Cd耐受性分析A1~A3.无Cd 胁迫Col-1、atbsk7、RcBSK7-OE;B1~B3. 50 μmol·L-1 Cd 胁迫Col-1、atbsk7、RcBSK7-OE;C1~C3. 75 μmol·L-1 Cd 胁迫Col-1、atbsk7、RcBSK7-OEFig.9 Analysis of tolerance of Col-1,atbsk 7,RcBSK7-OE to heavy metal Cd A1-A3.Col-1,atbsk7,RcBSK7-OE without Cd stress;B1-B3.Col-1,atbsk7,RcBSK7-OE under 50 μmo·lL-1 Cd stress;C1-C3.Col-under 75 μmo·lL-1 Cd stress Col-1,atbsk7,RcBSK7-OE

图10 不同质量浓度Cd 胁迫下Col-1、atbsk7、RcBSK7-OE根长及侧根数分析A.侧根数量统计结果;B.根长统计结果;*0.01<P≤0.05,**0<P≤0.01Fig.10 Analysis results of the root length and lateral root number of Col-1,atbsk7,RcBSK7-OE under different Cd stress concentrations A.Statistical result of the number of lateral roots;B.Statistical result of the root length;*0.01<P≤0.05,**0<P≤0.01
3 讨论
关于蓖麻植株响应Cd 胁迫的生理学研究已有 文 献 记 载[11-12],在Cd 胁 迫 下,蓖 麻 植 株 的H2O2、丙二醛含量随Cd 胁迫浓度的增加而显著增加,结果导致细胞内ROS 积累。即生理水平研究结果主要强调Cd 胁迫对蓖麻植株产生氧化损伤,同时蓖麻植株通过提高抗氧化能力以维持氧化系统的稳态,此结果同样在蛋白质水平上得到验证。
3.1 蓖麻植株响应Cd 胁迫的差异蛋白质组学研究与讨论
3 种剂量的Cd 胁迫下,蓖麻根系中主要的下调蛋白质包括微管蛋白α链、果胶酯酶等参与细胞骨架及细胞壁合成相关蛋白质;铁氧还蛋白-NADP+还原酶、柠檬酸1,2-加氧酶等参与光合作用和能量代谢的蛋白质以及与DNA损伤修复相关蛋白质,随Cd 胁迫浓度的增加,蛋白质表达量下调越显著。
分析3 种剂量Cd 胁迫下,蓖麻根系中显著上调的蛋白质可知,300 mg·L-1的Cd 胁迫下,环氧水解酶、琥珀酸脱氢酶、细胞色素b5 蛋白(Cyt-b5)等多种参与三羧酸循环和细胞氧化磷酸化等途径的蛋白质表达量显著上调。以上蛋白质一方面可以作为中间电子传递体参与细胞中的各种氧化还原反应而调节蓖麻体内ROS 平衡,另一方面参与细胞氧化磷酸化途径并合成供植物直接吸收的能量ATP,其活性的提高对蓖麻抵抗Cd 胁迫具有重要的作用。值得注意的是,O-甲基转移酶、差向异构酶等参与木质素合成的蛋白质表达量特异性上调。研究发现[13],木质素是植物次生细胞壁的重要组分,不仅能维持植物的正常生命活动,还可以通过增强植物细胞壁隔水性和机械强度进而提高植物的抗逆性,即蓖麻根系细胞壁的固持作用有助于降低和缓解Cd 毒害。结合以上结果可知:300 mg·L-1的Cd 胁迫下,蓖麻主要通过阻隔根系对重金属Cd 的吸收以及提高自身抗氧化能力而抵抗Cd胁迫损伤。
2-Cys 过氧化物酶、超氧化物歧化酶等作为植物内重要的抗氧化酶系统,其表达量在700 mg·L-1的Cd 胁迫下的增加对清除细胞内的ROS 具有重要的作用,这一结果也与已往研究结果一致,Zhang 等[14]通过研究Cd 对蓖麻抗氧化酶活性的影响发现,蓖麻根系中超氧化物歧化酶和谷胱甘肽过氧化物酶的活性较高,它们能诱导根部有效解毒以提高蓖麻对Cd 的耐受性。此外,参与植物光合作用的蛋白质在本组中特异性上调,包括碳固定过程的关键酶(S)-2-羟酸氧化酶、1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)及参与叶绿素合成的4-羟基-3-甲基丁-2-烯基二磷酸还原酶等,以上蛋白质的积累表达可提高蓖麻的光合速率以抵抗Cd 胁迫。有研究表明[15],Cd 胁迫下,蓖麻叶片中镁(Mg)含量未出现明显下降趋势,Mg 能够防止Cd 阻碍叶绿体合成叶绿素,即同样证实蓖麻通过维持光合作用,从而减缓Cd 毒害作用。值得注意的是,亚精胺合酶1 的蛋白质表达量特异性上调,而亚精胺合酶作为多胺生物合成代谢中的关键酶,所合成的多胺(PA)对促进植物生长发育、提高植物抗逆性具有重要的作用。此外,有研究证实亚精胺合酶因可以抑制重金属Cd 向地上部分转运而被应用于重金属Cd 的解毒效应中[16]。当Cd胁迫质量浓度增加至700 mg·L-1时,蓖麻植株主要通过提高抗氧化酶活性、抑制Cd2+运转等防御解毒过程而提高对重金属Cd的耐受性。
1 000 mg·L-1的Cd 胁迫下主要的上调蛋白质中,除参与木质素合成、抗氧化酶系统、光呼吸途径及离子稳态等相关蛋白质呈现一定数量的积累外,发现催化半乳糖代谢的UDP-葡萄糖4-表异构酶及参与细胞程序性死亡的CASP 样蛋白1D1、急性创伤诱导产生的蛋白WIN1、缺氧条件下被诱导的乙醇脱氢酶[17]的表达量特异性上调。其中UDP-葡萄糖4-表异构酶催化半乳糖代谢在生成ATP 的同时,其生成的代谢产物UDP-葡萄糖可进一步参与葡萄糖醛酸及戊糖磷酸等代谢途径而参与植物逆境胁迫。CASP 样蛋白1D1、创伤诱导蛋白WIN1 等氧化应激蛋白质可能是蓖麻受到更严重的损伤时,机体通过启动自身免疫防御系统、激活坏死细胞的死亡程序以响应Cd 胁迫。以上结果说明,1 000 mg·L-1的Cd 胁迫下,蓖麻植株受损严重,即使通过阻隔根系对重金属Cd 的吸收、抑制Cd2+运转、提高抗氧化能力和光合作用也无法彻底恢复Cd 胁迫损伤。在此情况下,蓖麻通过诱导激活细胞程序性死亡以响应Cd胁迫。
3.2 蛋白激酶响应Cd胁迫机制研究与讨论
目前,关于螯合肽(PCs)、金属硫蛋白(MTs)、ABC 运转蛋白、重金属ATP 酶(HMAs)转运蛋白等蛋白质通过螯合作用响应重金属Cd 胁迫的分子机制已被多数研究证实[18],而与丝/苏氨酸蛋白激酶响应重金属Cd 胁迫相关的分子机制还未明确。本研究中通过蛋白质组学和RT-qPCR 验证结果选取蛋白激酶作为研究对象对其进行功能验证,一方面因其在转录水平、蛋白质水平的高表达量,另一方面希望可以寻找并探索蓖麻参与重金属Cd解毒的特异性分子机制。
丝氨酸/苏氨酸蛋白激酶是植物体内参与植物生长发育调控和抗逆研究的一类重要的蛋白激酶,常见的丝氨酸/苏氨酸蛋白激酶包括类受体蛋白激酶(RLKs)、丝裂原活化蛋白激酶(MAPKs)、钙依赖性蛋白激酶(CDPKs)等。本研究通过蛋白质组学研究发现,Cd2+诱导蓖麻根系内1种丝氨酸/苏氨酸蛋白激酶的表达,且在分子水平和生物学水平上均证实该蛋白质的表达对拟南芥响应Cd胁迫具有重要作用。类受体蛋白激酶在植物细胞外信号的感知和传导中起关键作用,叶瑶瑶[19]在水稻(Oryza sativa)响应Cd 胁迫研究中发现,RLK在水稻中的过表达可以增强抗氧化保护酶活性、减少水稻体内的Cd 含量,因此推测RLK 可能通过调节参与Cd 转运的基因表达以及减轻Cd 诱导产生的氧化胁迫等途径而增强植物对重金属Cd 的耐 受 性。此 外Ouelhadj 等[20]研 究 中 发 现,Hv-LysMR1在受Cd 胁迫和衰老损伤的大麦(Hordeum vulgare)叶片中被显著诱导,但还无法证实Hv-LysMR1的诱导表达是由于感知衰老信号、还是由于感知重金属胁迫、或者是由于因Cd 胁迫损害而引起的叶片衰老所导致,这可能需要进一步证实。丝裂原活化蛋白激酶通过3 种蛋白激酶MAPKKK,MAPKK和MAPK组成了级联信号系统以参与胞外信号传导通路,Jonak 等[21]研究发现,将紫花苜蓿(Medicago sativa)幼苗暴露在过量的Cu2+或Cd2+下,激活了SIMK、MMK2、MMK3 和SAMK 4 种不同的丝原激活蛋白激酶,但通过对激酶进行动力学分析发现,Cu、Cd 诱导的应激似乎是通过不同的信号转导通路介导的,Cu 作为氧化还原活性金属离子,可能通过诱导产生ROS 进而激活SIMKK 触 发SIMK 和SAMK 活 化,而 介 导Cd 激 活SIMK、MMK2、MMK3 和SAMK 的上游成分仍有待确定。而Ye 等[22]在拟南芥的研究发现,Cd 胁迫下,一氧化氮(NO)与MAPKs存在相互作用,即NO可能是MAPKs 的上游信号。与此同时,熊兵[23]通过在酿酒细胞中研究发现,Cd 镉胁迫应答参与细胞壁完整性(Cell Wall Integrity,CWI)和高渗透压甘油(High Osmolarity Glycerol,HOG)2 个MAPK 信号传导途径。钙依赖性蛋白激酶作为一种Ca2+传感器在响应逆境胁迫中发挥重要的作用[24]。结合钙依赖蛋白激酶响应逆境胁迫机制发现,其主要通过调节K+跨膜运输进而提高离子稳态和ROS稳态,从而缓解逆境胁迫对植物的氧化损伤。此外,14-3-3 蛋白与钙依赖蛋白激酶之间的交叉磷酸化及协同作用为其响应逆境胁迫提供了更多可能[25]。以上各类丝氨酸/苏氨酸蛋白激酶的作用机制为后续对RcBSK 7响应Cd胁迫机制的探索提供了更多的研究方法。
4 结论
结合已有研究及本研究结果来看,蓖麻根系作为Cd 富集的主要部位,并通过茎-芽转运至茎部以及叶部。蛋白质组学分析强调,暴露于Cd 胁迫中会导致蓖麻植株氧化还原系统的改变,ATP合成失调,离子稳态破坏等反应。这可能是Cd 胁迫造成蓖麻植株生长抑制的主要原因。
总之,根据iTRAQ 蛋白质组学研究分析,了解并揭示了蓖麻在不同剂量Cd 胁迫下的响应机制。同时,筛选出显著上调的蛋白质并完成其编码基因(RcBSK7)在拟南芥中的过表达,鉴定结果验证了该基因可以显著增强拟南芥对Cd 胁迫的耐受性,后续将在蓖麻中完成RcBSK7的过表达,并对其参与植物响应Cd 胁迫进行验证并对其响应机制进行探究,以期将该基因用于培育高Cd 耐受性的蓖麻品种。本研究加深了对蓖麻响应Cd 胁迫机制的认识,为蓖麻响应Cd 胁迫的基因鉴定和土壤中重金属污染修复提供了有价值的理论依据。
猜你喜欢
科学(2020年2期)2020-08-24
国际放射医学核医学杂志(2020年4期)2020-07-27
上海农业学报(2017年3期)2017-04-10
创新作文(3-4年级)(2015年8期)2015-09-15
医学研究杂志(2015年3期)2015-06-10
中国粮油学报(2015年5期)2015-02-06
天然产物研究与开发(2014年6期)2014-04-27
植物营养与肥料学报(2014年1期)2014-03-11
中国医学科学院学报(2013年6期)2013-03-11
中国医学科学院学报(2012年3期)2012-03-25
