
子午岭次生油松林立木竞争格局研究
2023-01-11 03:00邹书珍尹才佳
植物研究 2023年1期
邹书珍 尹才佳 杨 茜 马 龙 康 迪*
(1. 西华师范大学生命科学学院,西南野生动植物资源保护教育部重点实验室,南充 637002;2. 陕西省循环农业工程技术研究中心,杨凌 712100)
森林是陆地生态系统的重要组成部分,与陆地碳汇、气候调节和生物多样性维持均有重要的关系[1-2]。越来越多的证据表明,世界各地的森林生态服务功能正在迅速变化并面临着更大的风险[3-5]。近年来大尺度、大数据研究逐渐兴起,研究者常常认为气候是导致森林变化的主导因素[6-9],而关注森林自身结构动态和森林变化内因的报道有减少趋势。研究表明,树木对有限资源的竞争直接影响到树木的生存或死亡,并能够导致包括种群的动态和空间格局在内的群落结构的改变[10-11]。如,Berger 等[12]观测了加拿大西部森林超过50 a 后报道,群落内部竞争产生的驱动力比外部气候因素更能推动森林的动态变化。因此,森林立木竞争是森林演替的重要内在驱动力,需要持续研究和关注[13-15]。此外,森林空间格局是种内、种间关系和环境因素长期综合作用的结果[14-16],空间格局与竞争格局长期以来也被认为是2个相关且互为因果的过程,研究竞争及格局有助于森林动态的预测和风险的应对[17-18]。
竞争和格局形成的关键因素是资源的可用性,因此在资源匮乏地区研究森林竞争更具代表性的。如在我国黄土高原地区,由于长期人为及自然干扰,植被覆盖率由曾经的53.0%下降到20世纪中叶的6.1%[19],植被的长期丧失导致黄土高原面临着我国最严重的水资源匮乏问题。同时,黄土高原植被的保护与重建是我国现如今旱区生态修复的重点和难点所在。目前国内外学者多关注黄土高原森林植物多样性、水分动态、养分动态和土壤理化性质[20-22]。然而,对于黄土高原地区次生森林格局的形成和维持研究较少,尤其是在干旱背景下的竞争格局,这就导致了黄土高原地区森林发展演替的内在机制仍不清楚,各种修复措施理论依据不足。
研究干旱地区森林竞争格局必须以科学选择代表性的物种为前提。油松(Pinus tabuliformis)是黄土高原现存次生森林的主要树种,因具有良好的抗干旱、抗贫瘠和抗风性,也是我国干旱半干旱地区重要的水土保持造林树种。本研究以子午岭典型自然扩散的油松次生林群落为研究对象,从林区的中心到边缘,依据胸径梯度选取4个处于不同演替阶段的样地进行对比研究,分析立木竞争格局和空间格局特征和二者可能存在的内在联系。本研究的目的如下:(1)定量描述黄土高原地区次生油松林不同发育阶段的竞争强度;(2)阐明竞争的物种间差异和径级间差异;(3)明确森林不同发展阶段竞争格局和空间格局的变化。这些结果有助于更好地理解干旱地区森林“干扰—再生”过程的内在驱动机制,且有望为世界其他干旱地区的森林恢复策略提供科学依据。
1 材料与方法
1.1 研究区域概况
子午岭位于黄土高原腹地(33°50′—36°50′N,107°30′—109°40′E,见图1),面积2.3×107hm2,是黄土高原现存最大的次生林区。子午岭地区为典型的温带半干旱、温带半湿润气候过渡带,海拔1 300~1 700 m,最大相对高差400 m。区域内年均温7.4 ℃,冬季极端低温-27 ℃,夏季极端高温可达36 ℃。年平均相对湿度为65%,年平均降水量约587.6 mm,最大降水量多发于7—9 月。土壤类型包括黄土和森林灰褐土。洛河和泾河流经该地区。
研究地点选在黄土高原子午岭西北部(图1,为了更好地区别油松与其他落叶森林,采用冬季遥感影像制图),是次生油松林的主要分布区。该地区经历过数次“戍边屯田”等历史事件,森林曾遭到巨大破坏。大约150 年前人口迁出,耕地荒弃,植被开始恢复。据当地史料记载,在森林恢复初期,油松原产于该地区的一个很小的区域。经过100多年的自然扩散,局部连续的油松次生林已接近400 hm2。该地区次生油松林为典型温带针叶林群落,由单层乔木、稀疏灌木和草本3层构成,林隙、林窗中少量存在山杨(Populus davidiana)、白桦(Betula platyphylla)、蒙古栎(Quercus mongolica)和侧柏(Platycladus orientalis)等乔木;灌木层植株数量稀少,主要物种有鞘柄菝葜(Smilax stans)、胡枝子(Lespedeza bicolor)、陕西荚蒾(Viburnum schensianum)、木姜子(Litsea pungens)、灰栒子(Cotoneaster acutifolius)和峨眉蔷薇(Rosa omeiensis)等木本植物;草本层优势种为寸草(Carex duriuscula)、早熟禾(Poa annua)、野棉花(Anemone vitifolia)和老鹳草(Geranium wilfordii)等。

图1 研究所在流域及样地选择的地理位置Fig.1 Geographical location of sample watershed and plots
1.2 样地设置
经过全面的走访调查,依据空间代替时间法则,从油松林区的核心到边缘(沿着扩散演替的路径)选择了4 个处于不同演替阶段的研究样地(见表1)。本研究中演替阶段是指森林阶段内的子阶段,没有明显的分段特征,4 个阶段的区分首要依据林木平均胸径的大小确定,然后根据当地林业管理部门人员的描述验证。在每个样地中选择坡度、坡向差异小、处于油松适宜生长的海拔范围内,且生长发育良好、景观破碎度低、干扰较低的区域建立了4 块大小为0.25 hm2的大样方。在样方内,记录所有胸径大于2.0 cm 或高度大于2.0 m 的乔木的物种名称、胸径、平面二维坐标等数据。

表1 样地基本信息Table 1 Basic information of plots
1.3 数据处理
基于径级与龄级反应规律的一致性,采用径级代替龄级研究油松种群结构[1,11,16]。样地内的树木被分为9 个径级,其中胸径小于5 cm 为第一径级,随后每5 cm 一个梯度逐级递增,胸径40 cm 及以上为第9 径级。为方便下文表述,其中第1~3 径级(胸径小于15 cm)定义为小径级,第4~6 径级(胸径小于30 cm 大于等于15 cm)定义为中径级,其他(胸径大于等于30 cm)定义为大径级。
在竞争指数的计算中,每株乔木均被当做对象木进行一次独立的计算。竞争木是位于以对象木为圆心,其所在样地平均冠径为直径的圆形范围内的所有毗邻乔木。竞争指数计算采用Hegyi 法[23],并在调查范围内进行每木计算,计算公式如下:

式中:CIa为对象木a的竞争数;N为对象木a周围的竞争木的数量;Sa和Sn分别是对象木a和第n个竞争木的胸径;Dan表示对象木a和第n个竞争木之间的直线距离。
油松的空间竞争格局采用Kriging 插值法完成。油松空间分布格局及油松与其他树种的空间相关性分别采用一元O-ring 函数和二元O-ring 完成,详细方法见已发表文献[24-25]。
1.4 软件使用
竞争指数的计算使用R 3.5.0 软件完成;竞争格局分析和绘图采用ArcGIS 10.0 完成;空间格局分析和绘图分别采用Programita 2008 和Origin 8.5完成;地图绘制采用ArcGIS 10.0 和3ds Max 2013完成;其他柱形图、散点图、拟合图采用Origin 8.5完成。
2 结果与分析
2.1 群落树种组成及主要树种竞争指数
在4 个演替阶段的群落中,油松种群密度分别为1 476、996、800、648 株·hm-2;白桦种群密度为48、32、44、16 株·hm-2;山杨种群密度分别为64、28、24、12 株·hm-2。区域内3 种优势乔木的竞争指数结果表明,油松种群竞争较强(见图2)。在所有演替阶段,油松均表现为不对称的竞争模式,即箱图的箱体偏下,中位数小于平均值。这表示少量的高竞争个体提高了种群整体的竞争强度。

图2 不同演替阶段优势乔木的竞争指数PT.油松;BP.白桦;PD.山杨;箱型图上下端短横为最大、最小值,箱体上、中、下3条横线分别代表75%分位数、中位数、25%分位数,方形小白点表示均值;图A~D分别表示Plot 1~4Figure 2 Competition index of dominant trees in different succession stages PT.P.tabuliformis;BP.B.platyphylla;PD.P.davidiana;The top and end short lines of the box plots showed the maximum and minimum values;The upper,middle and bottom three lines of the box showed 75%,median and 25% values respectively;The little squares white dot showed the mean values;Subfigures A-D showed Plot 1-4
2.2 油松种群数量结构及竞争的不对称性
如图3 所示,在演替早期(Plot1 和Plot2)油松径级结构均表现为增长型。然而随着演替的进行,小径级个体减少而大径级个体增加,油松种群径级结构趋向于均一化,这代表着其种群数量趋于稳定(见图3:Plot3 和Plot4)。我们进一步研究了油松胸径和竞争指数的关系(见图4)。结果表明,油松个体竞争指数与其胸径呈显著负相关(其中Plot1 和Plot4P<0.05,Plot2 和Plot3P<0.01),这表明小径级个体竞争压力高于大径级个体,即竞争基于树木大小存在不对称性。
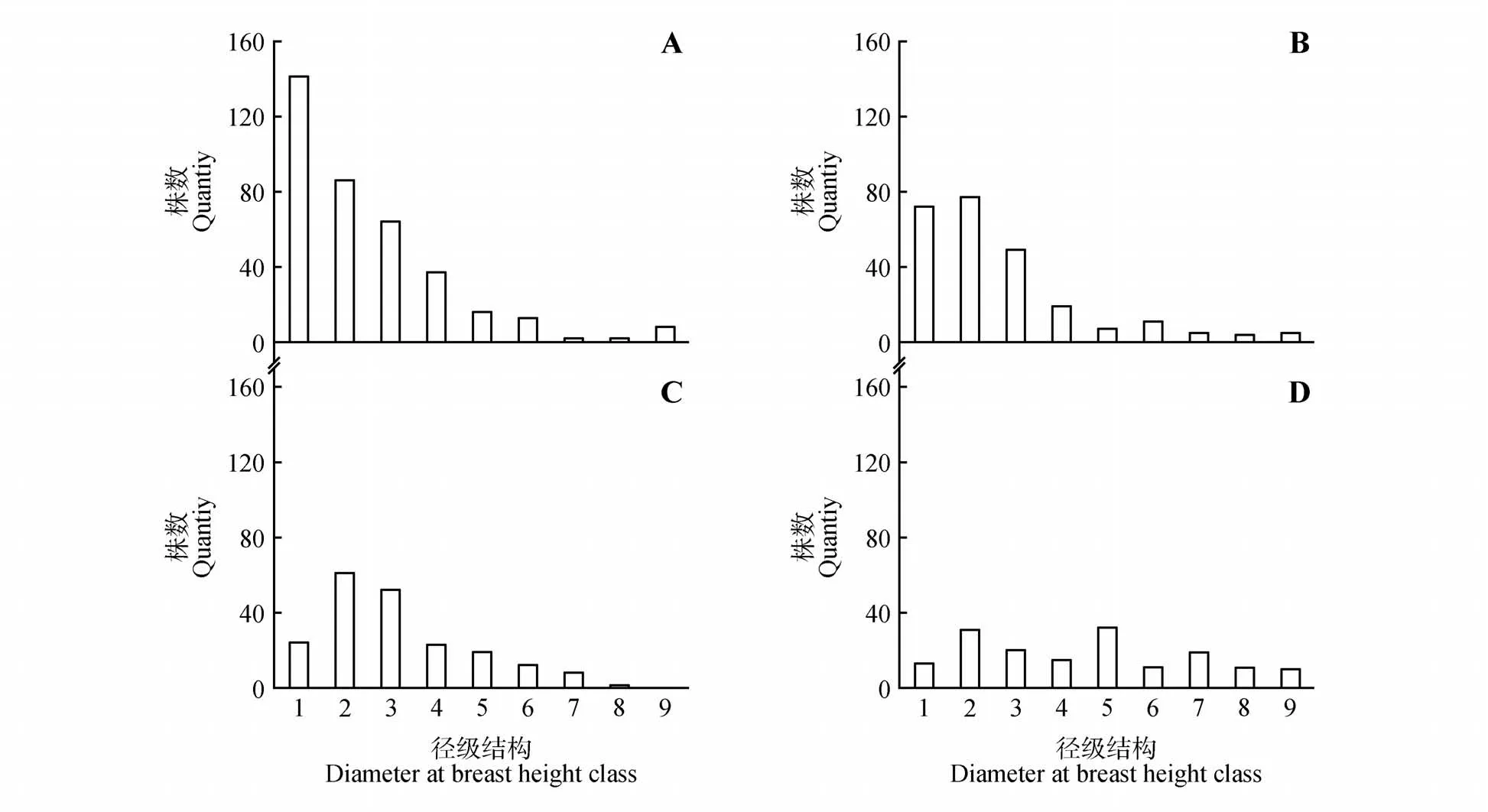
图3 不同演替阶段建群种油松种群径级结构图A~D分别表示Plot 1~4;下同Fig.3 Diameter at breast height structure of P.tabulaeformis in different succession stages Subfigures A-D showed Plot 1-4;The same as below

图4 不同演替阶段油松胸径与竞争指数的关系Fig.4 The relationships between diameter at breast height and competition index of trees of of P. tabulaeformis in different succession stages
2.3 油松空间竞争格局
在研究了黄土高原子午岭次生油松林竞争格局后发现,竞争指数在水平空间上也表现为不对称性,即高竞争个体以斑块状聚集存在于整个群落中(图5)。此外,在演替早期高竞争斑块的数量和范围均较大,随着演替的进行高竞争斑块的数量逐渐减少,但并没有完全消失。

图5 不同演替阶段油松竞争空间格局分析(Kriging插值法)Fig.5 Kriging interpolation analysis of the competition index of P.tabulaeformis in different succession stages
2.4 油松空间分布格局
利用单变量O-ring 函数分析了油松次生林立木空间分布格局(图6)。结果表明,油松在0~5 m尺度存在聚集分布现象,然而,随着演替的进行这种聚集的程度逐渐降低。在较大尺度,油松表现为随机分布模式。利用双变量O-ring 函数分析了群落内油松与其他乔木之间的空间相关性(见图7)。结果表明,在Plot1、Plot3、Plot4 中,油松与其他乔木在0~5 m 尺度存在空间负相关。在Plot2和Plot3 中,油松与其他树种在5~15 m 存在一定空间负相关。在各演替阶段的其他尺度,均无明显空间相关性。

图6 油松林立木空间格局分析虚线95%置信区间;cell:0.5 m×0.5 m;下同Fig.6 Spatial pattern analysis by single variable O-ring functions of P.tabulaeformis Area between the two dotted lines:95%confidence interval;cell:0.5 m×0.5 m;The same as below

图7 油松与群落内其他树种的空间相关性Fig.7 Spatial relations between P.tabulaeformis and other tree species
3 讨论
在本研究中,群落内乔木的数量随着森林演替过程而减少,特别是白桦、山杨等先锋树种减少较为明显,这种现象在生态学中称为相似性限制,通常是由竞争引起的[14,26-27]。研究结果表明,油松种群的竞争要高于白桦与山杨(见图2)。这种竞争的物种间差异常与其自身生物学特性有关,例如油松种子较重通常落在母树附近,故新生个体多聚集分布于母树附近,密度高导致了较强的种内竞争[27-28]。相比之下,白桦能产生大量、轻质、有翅的种子,具有较强的扩散能力,可选择生境资源较为宽裕处定居。此外,白桦不耐阴,因此其幼苗更容易在乔木密度较低且光照充足的环境定居,而扩散至高密度区域的繁殖体会被淘汰[18,29]。
种间竞争会导致先锋物种数量的减少,而种内竞争会导致后继物种的自疏,这是导致建群种油松种群数量减少的原因[14,30]。为了进一步解释油松种群数量减少的机制,我们研究了其种群数量结构,结果表明油松种群的数量减少主要是因为小径级个体的减少所致(见图3)。因此,黄土高原油松种内竞争的自疏作用机理是限制幼小个体的定居或建成。与本研究结论相似,以往研究曾报道山毛榉(Fagus longipetiolata)通过限制养分和光照资源,来限制其幼苗的生长。在演替过程中,随着上层林冠郁闭度提高以及干旱地区生境资源的相对匮乏,成树具有较强地上和地下竞争能力,而幼树则受到不利的影响。这是一个典型的非对称资源占有模式,导致了种群数量从增长型转化为稳定型。
至此,我们证实了演替后期油松种群数量减少是因为小径级个体的减少,并且假设这是由于个体大小差异导致的不对称竞争所致。为了进一步验证这个假设,本研究分析了每木胸径与竞争指数的关系(见图4)。结果表明,在本研究关注的任何演替阶段,油松个体胸径与其竞争指数均呈现负相关,这表明黄土高原油松次生演替过程中始终存在着基于个体大小的非对称竞争。与本结论类似,近年有研究建议森林竞争研究中可考虑冠层的重叠交错结构模型,这是因为树木对光的竞争在个体大小上是不对称的[31]。在演替的后期,干旱森林乔木更新受到抑制,仅能依赖种群个体死亡所产生林隙和林窗进行补充,竞争也将趋于缓和[11,32-33]。
本研究结果还表明,竞争除了存在个体大小上的不对称性之外,还存在空间格局上的不对称,即高竞争压力以斑块状分布于整个群落中(见图5)。并且演替早期,强竞争斑块在数量和面积上均大于演替后期。Metsaranta 等[34]对加拿大班克松(Pinus banksiana)空间分布的研究,观察到立木聚集性随着时间的推移而逐渐消失。但在本研究中,高竞争斑块虽然在数量、范围和强度上均有所减小,但并没有完全消失。这可能是区域环境的异质性决定的,即非对称竞争能够驱动种群个体趋向于更加均一化的空间分布格局,但并不能让种群绝对的均匀[11,33]。
为了进一步解释竞争产生的原因,我们分析了油松空间分布格局(见图6)。结果表明,油松在不同演替阶段均表现出小尺度的聚集分布格局。然而随着森林的发展,聚集程度逐渐降低。此外,这些油松聚集分布的尺度为0~5 m,这与研究区域油松成体冠幅相当,这表明黄土高原油松邻体竞争的主要竞争因子为光因子,竞争方式主要源于冠层的重叠[16]。在更大尺度上,油松表现为随机分布。我们还发现在演替早期,油松与其他树种在小尺度呈现负相关,在大尺度无显著相关性(见图7)。
在主要乔木的空间相关分析中,我们发现其中2 个演替阶段(图7:Plot 2-3)油松与其他树种在5~15 m 尺度存在负相关,这个尺度已经远远超出了油松与其他物种树冠能够重叠的范围。这种现象可能是由于环境的异质性导致的,如:环境中土壤、水分和营养的差异能够为不同的物种提供其适宜生长的环境[35-37];坡度、光照和土壤氮的非均质性可能是物种分布格局建成和森林更新的重要驱动力[38-39]。还有研究表明,微地形、土壤和水分有关的环境异质性能够控制森林结构[27,40]。然而,这些发现和推测在空间尺度上已经超出了人们通常认知的“邻体竞争”范围,导致黄土高原次生油松林这种特殊格局的真正原因还需要未来更加深入地研究。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
南方农业·下旬(2022年4期)2022-05-24
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
山西林业(2021年2期)2021-07-21
中国水土保持科学(2019年6期)2019-04-26
现代园艺(2017年22期)2018-01-19
林业勘察设计(2017年4期)2017-07-06
西北植物学报(2015年9期)2015-07-05
