
胍基乙酸对许氏平鲉生长性能、消化吸收、代谢及抗氧化能力的影响
2023-12-21 16:22郝甜甜于朝磊孙永智李宝山王晓艳王成强黄炳山
广东海洋大学学报 2023年6期
郝甜甜,于朝磊,孙永智,李宝山,王晓艳,王成强,李 璐,黄炳山
(1.山东省海洋资源与环境研究院/山东省海水渔用饲料工程技术研究中心/水生动物营养与饲料研发创新示范平台/山东省海洋生态修复重点实验室/烟台市海珍品质量安全控制与精深加工重点实验室,山东 烟台 264006;2.烟台经海海洋渔业有限公司,山东 烟台 264003)
许氏平鲉(Sebastes schlegelii),隶属于鲉形目(Scorpaeniformes) 鲉科 (Scorpaenidae) 平鲉属(Sebastes),具有肉质鲜美、营养丰富、抗病力强等优势,随着国家政策的支持和消费市场的日益成熟,已成为我国北方沿海地区深水网箱养殖对象,为具有开发前景的海水鱼类养殖鱼种之一[1]。近年来,本课题组在许氏平鲉配合饲料氮脂比[2]、不饱和脂肪酸[3]、维生素[4]等方面开展研究,而许氏平鲉在深水网箱养殖中受风浪、潮汐和水温、季节等环境因素影响,导致饲料中的蛋白、脂肪等能量物质更多应用于环境应激而不是用于生长,严重影响许氏平鲉深远海网箱养殖的经济效益。因此,开发功能性饲料添加剂,通过营养调控实现许氏平鲉将营养物质更多转化为生长而不是应激供能是目前面临的课题。
胍基乙酸(guanidinoacetic acid,GAA)又称胍乙酸、乙酸胍,N-咪基甘氨酸,是一种甘氨酸衍生物,也是肌酸合成的唯一前体物质,与磷酸肌酸一起通过三磷酸腺苷(ATP)再生参与细胞能量代谢[5]。GAA 目前已被欧盟纳入饲料添加剂目录。已有研究表明,饲料中添加胍基乙酸具有提高动物生长性能[6]、机体氨基酸代谢、肌酸合成、抗应激和抗氧化等诸多功能[7-8]。Lim 等[9]报道,GAA 结合磷酸基团形成的GAA-p 能使卷吻沙蚕(Glycera chirori)在海底火山沉积物这种极端环境中瞬时供能,趋利避害;王子苑等[10]发现在肉牛日粮中补充GAA 能提高磷酸肌酸和ATP 等高等能量物质的存贮量或利用效率;Degroot 等[11]在肉仔鸡实验中发现,在精氨酸不足的饲料中添加GAA,GAA 对内源精氨酸具有保留效应,可使精氨酸更多的用于蛋白质合成,从而提高动物机体的生长性能。但目前GAA 在饲料中的应用研究多见于畜禽,水产方面仅见于草鱼(Ctenopharyngodon idella)[12]、建鲤(Cyprinus carpiovarjian)[13]等淡水鱼,而GAA 在海水鱼上的研究鲜有报道。因此,本实验研究GAA对许氏平鲉生长性能、消化吸收、代谢及抗氧化能力的影响,评估GAA在实用饲料中的适宜添加水平,旨在为GAA在许氏平鲉网箱养殖配合饲料中的应用提供理论依据和技术支撑。
1 材料与方法
1.1 实验设计与饲料制作
以鱼粉、大豆浓缩蛋白、发酵豆粕和发酵棉粕为主要蛋白源,鱼油和豆油为主要脂肪源,设计粗蛋白质量分数为44.00%、粗脂肪质量分数为9.00%的基础饲料配方。在基础饲料中分别添加质量分数0.00%、0.01%、0.02%、0.04%、0.08%的胍基乙酸(GAA,纯度≥99%,购自湖南泰仁药业有限公司),配成5种等氮等脂的配合饲料,分别命名为D0、D1、D2、D3 和D4 组,其中D0 组为对照组。制作饲料时,所有饲料原料粉碎过孔径0.25 mm筛,按饲料配方表(表1)混合均匀,加入鱼油、豆油及适量水,再次混匀,经螺旋挤压机加工成粒径6 mm 的饲料颗粒,60 ℃烘干后放置-20 ℃冰箱备用。表2 为实验饲料营养组成。
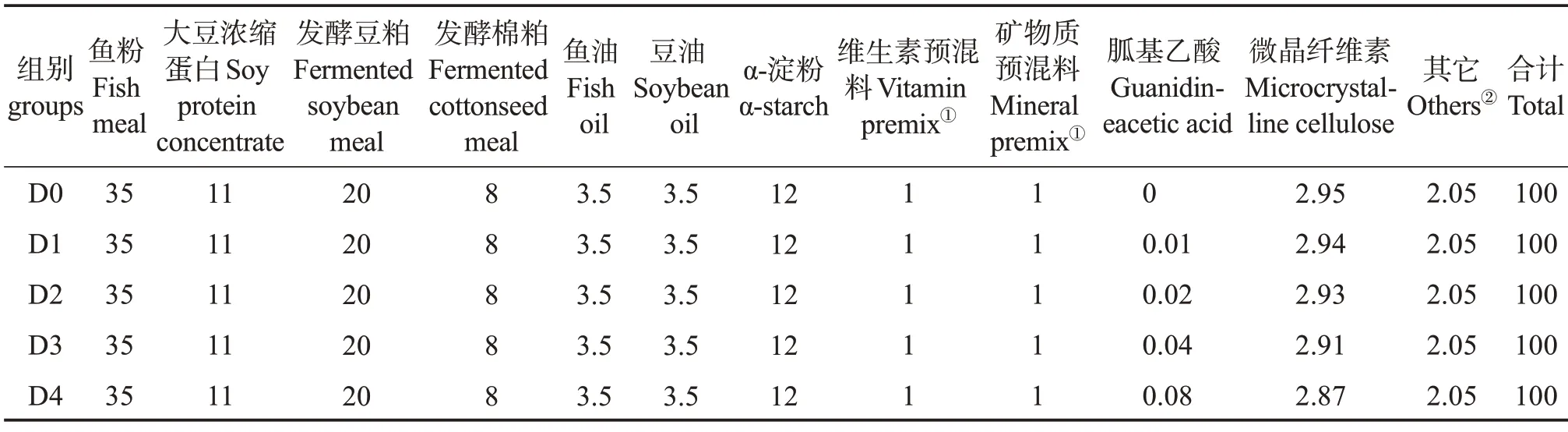
表1 基础饲料组分质量分数Table 1 Mass fraction of basic diets components %
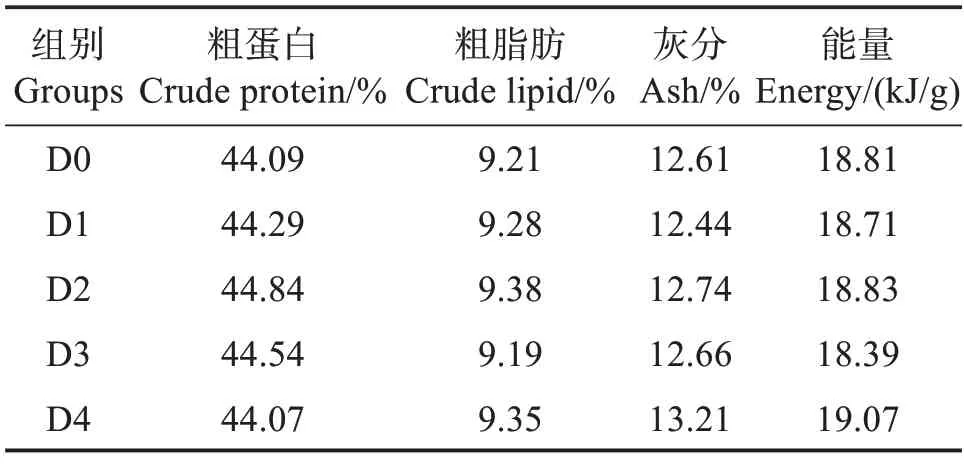
表2 实验饲料常规营养水平(干基)Table 2 Nutrient levels of experimental diets(dry mass basis)
1.2 实验用鱼和实验条件
实验鱼购自烟台经海海洋渔业有限公司,养殖地点为山东省长岛县离岸网箱养殖区。正式开始实验之前将实验鱼放入置于网箱(3 m×2 m×1 m)中暂养1 周,暂养期间投喂对照组饲料。暂养结束后,禁食24 h,挑选体格健壮,规格均一的许氏平鲉[初始体质量(125.00±1.00)g]随机分成5 组,每组3个网箱(1 m×1 m×1 m),每个网箱20 尾鱼。养殖时间为2023 年9 月28 日至11 月15 日,共49 d,每天投喂2 次(8:00、17:00),日投喂量为鱼体质量的1.5%~2.0%,观察每日摄食情况,按实际摄食情况调整投喂量。养殖期间如有死鱼记录数量并称质量。整个实验期间,自然海水温度(14~22 ℃)。
1.3 样品采集
样品采集过程严格按照山东省海洋资源与环境研究院实验动物伦理委员会要求执行,符合动物保护、动物福利和伦理原则。养殖实验结束后,实验鱼禁食24 h,采用质量浓度为30 mg/L 丁香酚(纯度≥98%)麻醉,以网箱为单位称质量,记录每网箱实验鱼的数量和体质量,计算成活率、增重率、特定生长率和饲料系数。每桶随机选择10尾鱼,分别测体质量、体长,计算肥满度,并抽血后解剖内脏分离肝脏;血样4 ℃静置4 h,以4 000 r/min 离心10 min,取血清,-70 ℃待测。
1.4 测定指标与方法
1.4.1 生长指标
式中,m0为实验开始时鱼体质量(g),mt为实验结束时鱼体质量(g),t为养殖周期(d),L为实验鱼体长(cm),mf为摄食量(g),wp为饲料蛋白质量分数,Ni、Nf分别为实验开始和结束时实验鱼存活数。
1.4.2 营养成分分析 饲料和全鱼的营养常规成分测定按照AOAC 方法测定。水分采用105 ℃恒温干燥法(GB/T6435—2014),粗蛋白采用凯氏定氮法(GB/T6432—2018),粗脂肪采用索氏抽提法(GB/T6433—2006),粗灰分采用550 ℃失重法(GB/T6438—2007)。饲料能量采用燃烧法测定(IKA,C6000,Germany)。
1.4.3 肠道消化酶活力测定 肠道胰蛋白酶、淀粉酶、脂肪酶活力均采用南京建成生物工程研究所试剂盒测定,具体操作严格按照各试剂盒说明书执行。酶液蛋白质浓度采用考马斯亮蓝法测定。
1.4.4 血清生化指标测定 血清中谷草转氨酶(aspartate transaminase,AST)、谷丙转氨酶(alanine aminotransferase,ALT)活力、甘油三酯(triglyceride,TG)、总胆固醇(total cholesterol,TCHO)、高密度脂蛋白胆固醇(high density lipoprotein cholesterol,HDL-C)、低密度脂蛋白胆固醇(low density lipoprotein cholesterol,LDL-C)、血糖(glucose,GLU)含量均采用南京建成生物工程研究所试剂盒测定。
1.4.5 肝脏糖代谢酶活力和抗氧化酶活力指标测定 将肝脏样品与无菌生理盐水按照质量(g)体积(mL)比1∶9 冰浴下匀浆,以4 ℃、4 000 r/min 离心10 min,收集上清液获得肝脏匀浆液,用于肝脏糖代谢酶活及抗氧化酶活指标的测定。肝脏丙酮酸激酶(pyruvate kinase,PK)、磷酸果糖激酶(phosphofructokinase,PFK)、肌酸激酶(creatine Kinase,CK)酶活和肌酸(creatine,Cr)活力采用Elisa 试剂盒(南京建成生物工程研究所)测定;肝脏总抗氧化能力(total antioxidant capacity,T-AOC)、超氧化物歧化酶(superoxide dismutase,SOD)活力、丙二醛(malondialdehyde,MDA)含量、谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-PX)活力采用试剂盒检测(南京建成生物工程研究所)。肝脏匀浆上清液蛋白质浓度采用考马斯亮蓝法测定。
1.5 数据分析
采用SPSS 17.0 对所得数据进行单因素方差分析(One-Way ANOVA),差异显著,则采用Duncan 氏进行多重检验,显著水平为α=0.05,统计数据以平均值±标准差形式表示。
2 结果
2.1 胍基乙酸对许氏平鲉生长性能的影响
由表3 可知,各组间许氏平鲉成活率无显著差异(P>0.05)。饲料中添加胍基乙酸显著提高了鱼体的终末质量、增重率、特定生长率和蛋白质效率(P<0.05),其中增重率在D3组达到最高值,比对照组(D0 组)显著提高33.48%。饲料系数则呈相反趋势,在D1、D2、D3 和D4 组显著低于D0 组(P<0.05),并在D3 组达到最低值。饲料中添加胍基乙酸对鱼体肥满度无显著影响(P>0.05)。
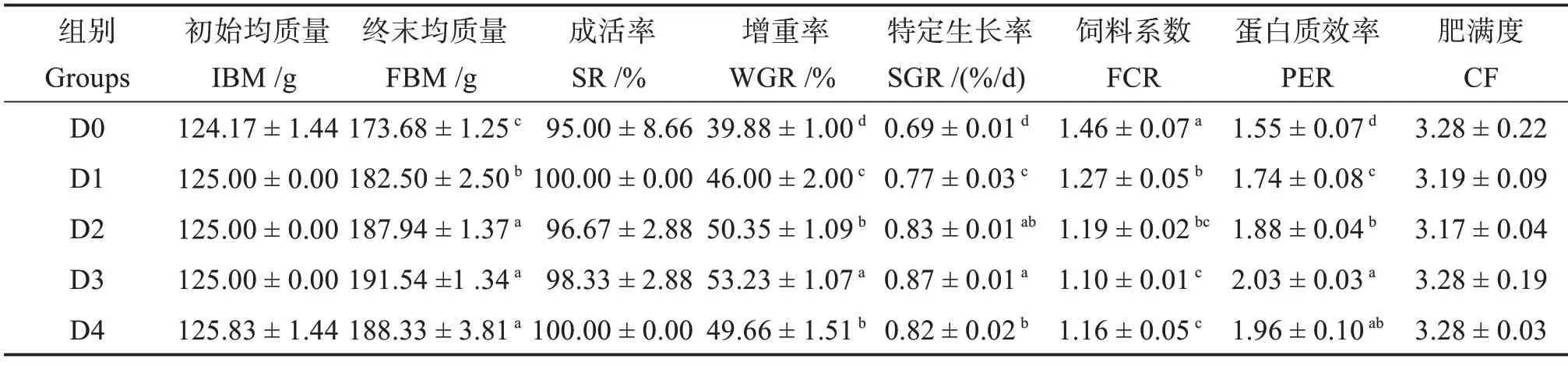
表3 胍基乙酸对许氏平鲉生长性能和形体指标的影响Table 3 Effects of guanidine acetic acid on growth performance and the morphometric parameters of Sebastes schlegelii.
2.2 胍基乙酸对许氏平鲉全鱼体成分的影响
由表4可知,随饲料中胍基乙酸含量升高,全鱼粗蛋白和粗脂肪含量虽有上升趋势,但各组之间差异不显著(P>0.05)。各组之间全鱼水分和粗灰分含量无显著差异(P>0.05)。
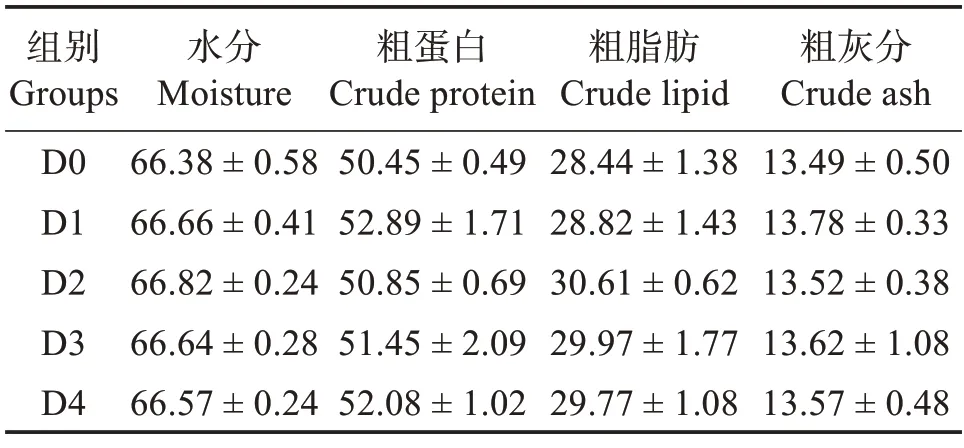
表4 胍基乙酸对许氏平鲉全鱼体成分的影响Table 4 Effect of guanidine acetic acid on proximate composition of the whole body of Sebastes schlegelii
2.3 胍基乙酸对许氏平鲉肠道消化酶的影响
由表5 可知,饲料中添加胍基乙酸显著影响了许氏平鲉肠道胰蛋白酶活力、脂肪酶活力和淀粉酶活力(P<0.05)。胰蛋白酶活力呈先上升后下降趋势,在D3 组达到最高值,显著高于其他各组(P<0.05);脂肪酶活力则在D3 和D4 组显著高于其他3 组(P<0.05);淀粉酶活力变化趋势与胰蛋白酶一致,在D1、D2、D3 和D4 组显著高于对照组(P<0.05)。
2.4 胍基乙酸对许氏平鲉血清生化指标的影响
由表6 可知,饲料中添加胍基乙酸对血清中ALT 和AST 活力无显著影响(P>0.05)。胍基乙酸显著提高了血清中TG 含量,其中在D3 组达到最高值,显著高于D0—D2 组(P<0.05),但与D4 组无显著差异(P>0.05);TCHO 含量呈稍上升趋势,但各组之间无显著差异(P>0.05);高密度脂蛋白胆固醇在D1—D4 组显著高于对照组(P<0.05);低密度脂蛋白胆固醇在各组之间无显著差异(P>0.05)。各组之间血糖浓度无显著差异(P>0.05)。
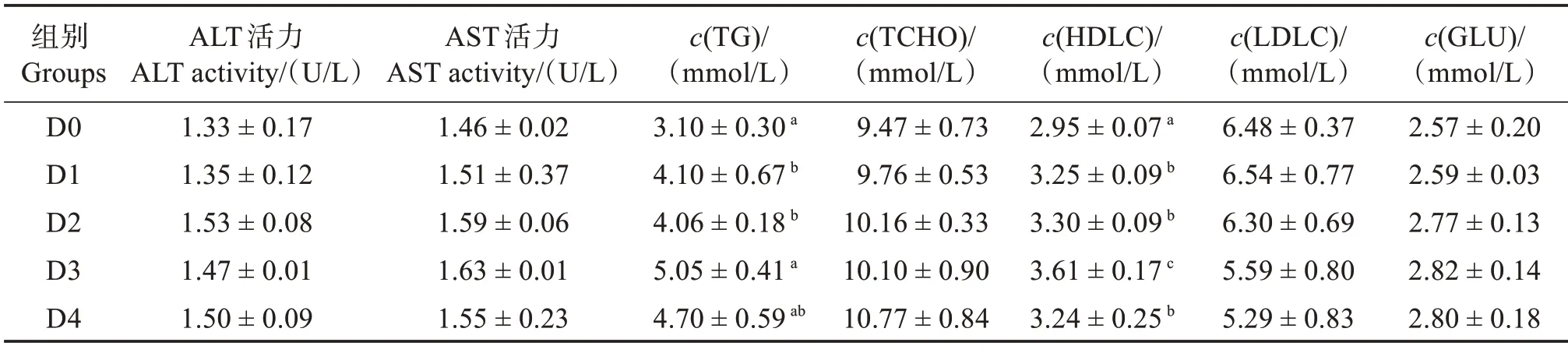
表6 胍基乙酸对许氏平鲉血清生化指标的影响Table 6 Effect of guanidine acetic acid on serum biochemical indices of Sebastes schlegelii
2.5 胍基乙酸对许氏平鲉肝脏糖代谢酶活的影响
饲料中添加胍基乙酸对许氏平鲉肝脏糖代谢酶活产生了显著影响(P<0.05)(表7)。PK 活力和CK 活力均在D1、D2、D3 和D4 组显著低于对照组(P<0.05);PFK 则随胍基乙酸水平变化呈先下降后上升趋势,在D3 组达到最低值,显著低于对照组(P<0.05),其他组则与对照组无显著差异(P>0.05);Cr 含量则随胍基乙酸水平变化呈上升趋 势,在D2—D4 组显著高于D0 和D1 组(P<0.05)。
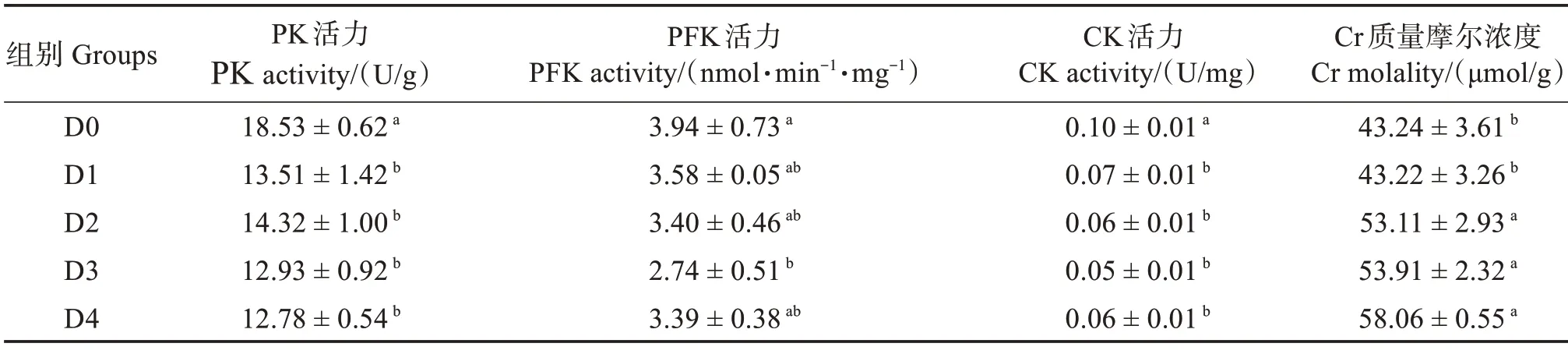
表7 胍基乙酸对许氏平鲉肝脏糖代谢酶活的影响Table 7 Effect of guanidine acetic acid on carbohydrate metabolic enzymes in the liver of Sebastes schlegelii
2.6 胍基乙酸对许氏平鲉肝脏抗氧化能力的影响
由表8 可知,饲料中添加胍基乙酸显著提高了许氏平鲉肝脏的抗氧化能力,其中T-AOC 在D1—D4 组显著高于对照组(P<0.05),SOD 活力在D2、D3 和D4 组显著高于对照组(P<0.05);GSH-PX 活力则随饲料中胍基乙酸水平的生长呈先上升后下降趋势,在D2 和D3 组显著高于D0、D1 和D4 组(P<0.05);MDA 含量在D1—D4 组显著低于对照组(P<0.05)。
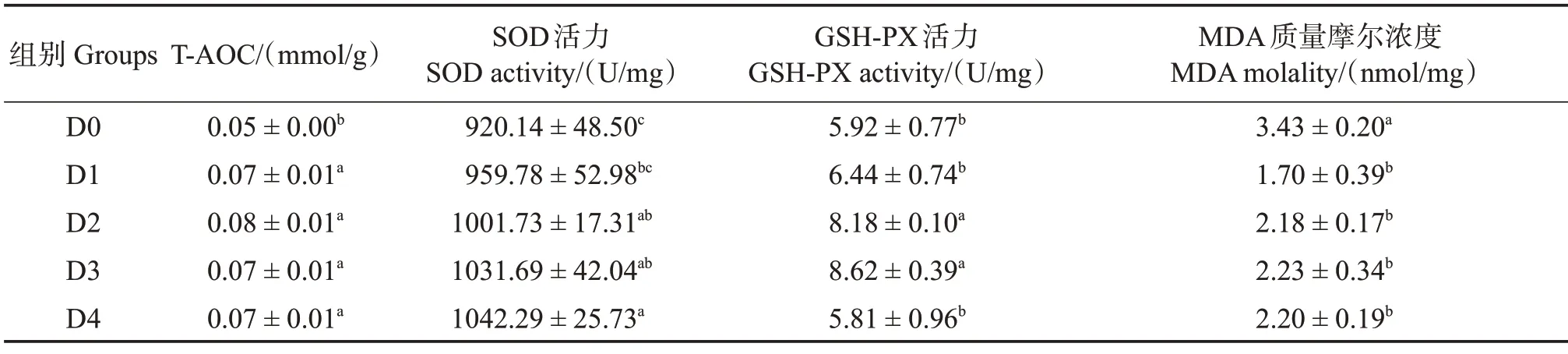
表8 胍基乙酸对许氏平鲉肝脏抗氧化能力的影响Table 8 Effect of guanidine acetic acid on antioxidant capacities in the liver of Sebastes schlegelii
3 讨论
3.1 胍基乙酸对许氏平鲉生长性能和体成分的影响
生长性能是衡量深远海网箱养殖鱼类经济效益的重要指标,提高生长性能对于提高深远海网箱养殖竞争力尤为关键。班博等[14]研究表明,饲料中添加胍基乙酸,可促进动物机体能量代谢,减少碳水化合物、脂肪和蛋白质分解功能,提升动物生产性能,提高饲料利用率。研究发现,在饲料添加质量分数0.12%~0.18% 的GAA,尼罗罗非鱼(Oreochromis niloticus)增重率显著提高[15];饲料中添加300 mg/kg GAA 显著提高草鱼增重率[12];饲料中添加0.4 g/kg GAA 显著提高牛蛙(Lithobates catesbeianus)的生长性能[16]。本实验结果也表明,饲料中添加GAA(质量分数0.01%~0.08%)显著提高许氏平鲉增重率和特定生长率,降低了饲料系数。基于国内外对GAA的研究,认为其促进动物生长机制主要与能量补充、节约精氨酸和促进肌纤维发育等3 个方面密切相关:(1)饲料中补充GAA 可提高磷酸肌酸和肌肉中的肌酸含量,为细胞能量代谢提供更多ATP,减少了机体脂肪和蛋白质耗能[17],从而提高鱼体生长性能;(2)饲料中添加外源GAA减少了机体内源精氨酸消耗,可促使更多精氨酸用于蛋白质合成、细胞信号转导和激素释放等生理功能,进而促进鱼体生长[18];(3)GAA 可通过抑制miR-133a-3p和miR-1a-3p的表达激活Akt/mTOR/S6 信号通路,改善肌肉组织能量,促进肌纤维的发育,进而改善动物机体生长性能[19]。肥满度可以作为衡量鱼体能量状态的常用形体学参数[20]。本研究发现,饲料中添加GAA对许氏平鲉肥满度无显著影响,推测原因可能与GAA对能量代谢的调控能力有关,使得鱼体脂肪的储备和消耗处于动态平衡状态。
李增民[21]研究表明,GAA 可以通过提高肝脏中氨基酸代谢酶(GGT)和核酸代谢酶(ADA)的活力来促进机体蛋白质的合成和沉积。但本实验结果显示,各组间鱼体水分、粗蛋白、粗脂肪和粗灰分含量差异不显著,表明饲料中添加GAA对鱼体营养成分变化无不良影响,与在建鲤[13]、尼罗罗非鱼[15]上研究结果一致。
3.2 胍基乙酸对许氏平鲉肠道消化酶的影响
鱼体肠道消化酶活力反应机体消化吸收和营养利用的能力,与鱼体生长发育息息相关[22]。本实验结果表明,饲料中添加质量分数0.01%~0.08%GAA 显著提高了许氏平鲉肠道胰蛋白酶和淀粉酶活力,0.04%和0.08%组显著提高了肠道脂肪酶活力,说明GAA能够通过提高肠道消化酶活力来促进鱼体生长,这与本研究中许氏平鲉的生长结果一致。GAA 对鱼体肠道消化酶的研究还未见报道。在畜禽上有研究证实,GAA 可以通过促进有益菌的增殖和增强消化道内α-淀粉酶、蛋白酶活力来改善动物机体消化道内环境及pH 值[23],说明GAA 可以协同改善肠道菌群及提高消化酶活力来达到促进鱼体生长的效果。
3.3 胍基乙酸对许氏平鲉血清生化指标的影响
Abdel-Tawwab 等[24]研究表明血清生化指标能反应机体健康状况。已有研究发现在畜禽饲料中添加1 200 mg/kg 的GAA 对动物机体是安全的[25],但缺少鱼体方面数据。AST 和ALT 酶活力不仅可以反映机体代谢程度,还与肝细胞炎症和变形密切相关,因此AST 和ALT 酶活力的高低可以作为判定肝功能是否受到损伤的指标[26]。本研究结果发现,饲料中添加GAA 对血清中AST 和ALT 酶活无显著影响,表明饲料中添加GAA对许氏平鲉肝脏未造成损伤。血脂主要的代谢指标由TG 和TCHO 组成,包括HDL-C 和LDL-C,HDL-C 通过胆固醇逆向转运把外周组织及血浆中的胆固醇运回肝脏代谢,LDL-C 则是向组织转运肝脏合成的内源性胆固醇[27-28]。徐超等[29]研究表明,正常状态下,鱼体脂质代谢处于动态平衡状态,当血脂水平在正常范围内上升时,表明鱼体机体内脂肪转运程度相应增强,与鱼体生长性能密切相关。张翼等[30]研究发现,在育肥肉牛饲料中添加GAA后,降低了血清中低密度脂蛋白胆固醇含量。本实验结果发现,饲料中添加质量分数0.01%~0.08%的GAA 显著提高了血清TG和HDL-C含量,LDL-C含量有下降趋势,但差异不显著,说明饲料中添加GAA能一定程度提高许氏平鲉血清中的脂质代谢。推测原因是GAA 通过提高鱼体血清脂质代谢速率用于提高鱼体的生长速度[31],与本实验生长数据相吻合。
Marco 等[32]早期实验发现,大鼠体外注射GAA后,能够激活胰岛素信号通路,而胰岛素增加会导致血糖浓度降低,本实验结果发现,饲料中添加GAA 并没有显著影响许氏平鲉的血糖浓度。Polakof 等[33]报道,血糖浓度变化受多种因素制约,如养殖条件、物种、鱼体代谢能力、鱼类对胰岛素的敏感性及鱼体激活胰岛素的能力等,造成以上结果的差异,可能是因为物种不同导致其对胰岛素的敏感性及激活胰岛素的能力不同。
3.4 胍基乙酸对许氏平鲉肝脏糖代谢酶活的影响
肝脏是鱼类糖脂代谢的主要场所[34]。糖酵解是鱼体葡萄糖分解代谢的关键途径,而PK 和PFK 则是糖酵解途径中的两个关键限速酶[35]。Yang 等[36]报道,鱼类为适应环境胁迫会采取一系列与糖酵解相关的酶完成氧化磷酸化过渡到糖酵解转化,这种反应是一种进化保守,是鱼体适应环境胁迫的一种有效策略。如在鲤(Cyprinus carpio)[37]和半滑舌鳎(Cynoglossus semilaevis)[38]研究中发现,鱼体受到环境胁迫时会启动糖酵解途径以增加葡萄糖摄取用于维持能量需求。李洁蕾等[39]报道,在保育猪饲料中补喂600 mg/kg GAA 21 d 后显著增加了血浆中PK 和PFK 酶活力,但本实验研究结果显示,饲料中添加质量分数0.01%~0.08%GAA显著降低了许氏平鲉肝脏PK 含量,在0.04%和0.08%水平下显著降低了肝脏中PFK 含量,说明GAA 可以减少许氏平鲉肝脏中葡萄糖的分解代谢,使葡萄糖更多的以肝糖原形式储存起来,其原因或许为GAA作为肌酸的前体物质,可以直接增加机体生命活动所需的ATP,用以缓解深远海网箱养殖面临的风浪、潮汐、水温等环境应激。关于GAA 在许氏平鲉养殖中的抗应激作用还需进一步研究。
肌酸激酶(CK)与肌肉收缩、细胞能量转运和ATP 再生有直接关系,并在肌酸代谢中发挥着能量缓冲的作用[40]。本实验结果表明,饲料中添加GAA(质量分数0.01%~0.08%)降低了许氏平鲉肝脏中肌酸激酶活力,而在尼罗罗非鱼[15]、牛蛙[16]上等研究表明饲料中添加胍基乙酸可以显著提高CK 活性,与本实验结果不一致,其原因可能为许氏平鲉作为肉食性海水鱼类其饲料中动物性蛋白源较多,保证了鱼体摄入的Cr 含量较为充足,导致额外添加的GAA 在不同营养源下的作用途径也不一致。肌酸不足时,GAA 可补充体内肌酸的缺乏,参与机体肌酸代谢进而上调肌酸激酶的活性,用于机体能量补充[41];而肌酸充足时,额外添加的GAA 基本不直接参与肌酸激酶功能[42],其功能主要使高磷酸肌酸和ATP 等能量物质储存起来[10],可用于抵抗网箱养殖面临的风浪、潮汐、水温等环境因子应激。这可能是造成本实验中许氏平鲉肝脏肌酸激酶活力降低的原因之一。
GAA在肝脏经甲基化后转化为Cr,Cr是细胞内能量代谢的重要物质,也是能量储存的临时场所[43]。本实验中,饲料中添加GAA显著提高了许氏平鲉肝脏中Gr 含量,说明饲料中额外补充的GAA 以Cr 的形式转化和保留,也是许氏平鲉机体Cr 的潜在来源,与在眼斑拟石首鱼(Sciaenops ocellatus)[8]上研究一致。
3.5 胍基乙酸对许氏平鲉肝脏抗氧化能力的影响
许氏平鲉网箱养殖中常受风浪、潮汐、水温和季节等因素的影响,鱼体在应对环境应激时会造成大量活性氧和自由基的积累。因此,在许氏平鲉网箱养殖中增加鱼体T-AOC、SOD、GSH-PX 等抗氧化酶活力和降低脂质过氧化反应中MDA 的含量来缓解氧化应激,对提高许氏平鲉网箱养殖经济效益具有重要意义。Aziza等[15]研究表明,在尼罗罗非鱼饲料中添加质量分数0.12%~0.18%的GAA,可显著提高鱼体SOD 和GSH 酶活和降低MDA 含量;Zeng等[5]发现,饲料中添加0.4~0.6 g/kg GAA 可显著提高牛蛙血清中T-AOC活性,降低MDA含量。在本实验中,饲料中添加质量分数0.01%~0.08% GAA 显著提高了许氏平鲉肝脏中T-AOC活性,降低了MDA含量;SOD活性则在0.02%~0.08%组显著高于对照组;GSH-PX活性随GAA 水平的变化呈先上升后下降趋势,表明GAA可在一定程度上通过增加内源性抗氧化酶活力达到提高机体抗氧化能力的效果,与上述研究结果一致。其作用机理为:(1)与GAA 的代谢产物Cr有关,Cr已被证实可作为外源抗氧化剂来提高动物的抗氧化能力[44];(2)外源GAA 的摄入,鱼体节约的部分内源性精氨酸能够淬灭自由基,发挥抗氧化作用[45]。
4 结论
本实验条件下,以增重率为评价指标,许氏平鲉饲料中GAA 的适宜添加量为0.04%;饲料中添加质量分数0.04%~0.08%GAA 可提高许氏平鲉肠道消化酶活力,并可通过调节代谢产物Cr的含量发挥抗氧化和能量调节作用。
猜你喜欢
水产养殖(2021年5期)2021-05-21
水电与抽水蓄能(2021年2期)2021-05-14
农业工程学报(2021年4期)2021-05-09
农业工程学报(2020年6期)2020-05-19
保健与生活(2018年17期)2018-01-27
电子制作(2017年24期)2017-02-02
渔业致富指南(2016年12期)2016-11-11
湖南农业(2016年3期)2016-06-05
现代盐化工(2015年3期)2015-01-23
食品工业科技(2014年9期)2014-03-11
