
光周期和维生素对巴夫藻生长和物质生产能力的影响
2023-12-21 16:22王雨涵董苗萍靳翠丽周晓见
广东海洋大学学报 2023年6期
刘 凡,王雨涵,董苗萍,靳翠丽,刘 青,周晓见
(扬州大学环境科学与工程学院,江苏 扬州 225127)
巴夫藻(Pavlovasp.)细胞微小,繁殖速度快,营养物质丰富,是公认的重要营养产品[1-2]。巴夫藻中富含岩藻黄素和二十碳五烯酸(Eicosapentaenoic Acid,EPA)。岩藻黄素是最丰富的海洋类胡萝卜素,由于优越的抗氧化性能,在制药、营养和化妆品行业具有广泛应用价值[1-2]。EPA 是一种长链n-3 多不饱和脂肪酸,在预防心血管疾病、炎症、自身免疫性疾病等方面发挥作用,已成为全球范围内稀缺的营养物质[3-4]。有研究发现Pavlovasp.OPMS 30543 中岩藻黄素的含量为8 mg·g-1左右,远高于作为岩藻黄素主要商业来源的大型褐藻[5]。而EPA 摄取的主要来源是鱼油,鱼类中EPA 是通过食用含有EPA 的微藻在体内累积的,而巴夫藻因其能大量合成EPA 而被认为具有替代鱼油的潜力[6]。另外,由于Pavlova属没有细胞壁,可以直接作为天然食品食用,而无需将细胞内的EPA 提取出来,该优势也有助于培养巴夫藻细胞作为整体食物的商业化生产进程[7]。
规模化微藻培养的生长和胞内物质生产受培养体系中的光照和营养条件影响[8]。截至目前,光照条件对巴夫藻影响的研究主要集中在光强度和光质方面,对于光周期的研究比较欠缺[9-11]。而近年来针对其他微藻的研究发现,光周期不仅可能影响微藻的物质生产、色素组成,还会影响脂肪酸组成和生物柴油质量[12-13]。维生素作为一种微藻依赖的微量有机生长因子,影响微藻的生长速率,细胞大小和叶绿素含量等[14-15]。本研究以巴夫藻(Pavlovasp.)为研究对象,设置不同的光周期和维生素浓度,以双因素实验研究光周期和维生素对巴夫藻生长和物质产量的影响,并分析脂肪酸组成以及生物柴油质量的差异,为提高巴夫藻培养效益提供实验依据。
1 材料与方法
1.1 藻种来源
巴夫藻由中国海洋大学教育部海水养殖重点实验室提供。培养基采用盐度为30 的海水f/2 培养基[9]。
1.2 培养条件和实验设计
实验设置3 种维生素浓度1V、4V、8V(1V 为f/2配方中的维生素浓度;4V 和8V 分别是该浓度的4倍和8 倍);3 种光周期,以24 h 为1 个周期,“光∶暗”分别为8 h∶16 h、16 h∶8 h、24 h∶0 h;共9 种处理,简记为维生素浓度+光周期,如1V 24 h∶0 h,每个处理3 个重复。250 mL 三角瓶中装有150 mL 培养液,取对数生长期藻种接种,接种后初始660 nm光密度(记作D660,以下相同)在0.03 左右。照度为(65±2)μmol·m-2·s-1,温度(24±1)℃,培养14 d,每天早晚各摇动2次。
1.3 测定方法
1.3.1 生长情况监测和生物量测定 每2 d 取200 μL 藻液于96 孔板中,用酶标仪318-MC 测其D660[16]。培养结束时,用血球计数板测定最终细胞密度[17]。采用干质量法测定最终的生物量。取120 mL 混合均匀的藻液,以3 700 r·min-1离心10 min,去上清后加入10 mL 纯水,震荡混匀后再次离心去上清,将沉淀转移至已烘干称好质量的玻璃瓶中,用冷冻干燥机FreeZone 2.5 plus,-80 ℃真空冻干24 h后称质量,得到藻粉干质量[9]。
1.3.2 叶绿素a 含量测定 培养结束时,取5 mL 藻液,离心(4 000 r·min-1、10 min)去上清后加5 mL 丙酮4 ℃避光8 h 提取后,次日超声(700 W,超声5 s,间隔3 s)10 min、离心,取上清分别于470 nm、644 nm和661 nm处测定光密度,根据以下方程计算色素含量(mg·L-1)[18]。
1.3.3 岩藻黄素含量测定 取冻干后的藻粉,按1∶40(1 g 藻粉溶于40 mL 的无水乙醇)的料液比加入无水乙醇,然后在60 ℃水浴锅中避光浸提2 h,3 700 r·min-1离心10 min,转移上清液,重复上述操作至藻粉为灰白色,用分光光度计测定提取的上清液在445 nm 处的光密度,根据公式计算岩藻黄素的含量[4]。
w:岩藻黄素质量分数(mg·g-1);D445:岩藻黄素提取液在445 nm 处的光密度值;N:稀释倍数;V:粗提取液体积(mL)表示岩藻黄素质量浓度为1 g·L-1时,光径为1 cm 时的光吸收值;m:藻粉质量(g)。
1.3.4 蛋白质和多糖含量测定 培养结束时,取5 mL 藻液,3 700 r·min-1离心10 min,去上清,加入5 mL去离子水,用超声波细胞破碎机JY92-IID冰浴超声破碎(200 W,超声3 s,间隔2 s)8 min,离心留上清液,用于测定蛋白质和多糖。蛋白质含量用考马斯亮蓝法测定[19],多糖含量用苯酚-硫酸法测定[20]。
1.3.5 脂肪提取与脂肪酸组成测定 采用氯仿/甲醇抽提法测定脂肪含量:取20 mg左右冻干后的藻粉,加入0.8 mL 去离子水,1 mL 三氯甲烷,2 mL 甲醇,混合均匀后,冰浴超声破碎8 min,再加入1 mL三氯甲烷和1 mL去离子水,3 700 r·min-1离心10 min,将氯仿层转移至烘干称好质量的玻璃瓶中,常温氮气吹干后测其质量[21-22]。
称取2 mg 脂肪,加入1 mL 正己烷,超声10 min,加入0.2 mL 的KOH 甲醇溶液(11 g KOH 溶于100 mL 甲醇),超声处理10 min,加入0.125 g 无水硫酸钠,静置后取上层,用0.22 μm 的膜过滤,转移至进样瓶中用于GC-MS分析[22]。
GC-MS 分析:采用Thermo Scientific ITO900 GC-MS 系统,色谱柱为TR-5MS(30 m×0.25 mm×0.25µm),载气为He2(99.999%),流速0.8 mL·min-1,样品以不分流形式注入,进样口温度230 ℃,进样量1 μL。程序升温:120 ℃恒温1 min,然后以3 ℃·min-1升至240 ℃,保持10 min。离子源温度为220 ℃。溶剂延迟4 min,采用全扫描质谱模式,范围在50~800 amu,耗时0.68 s。然后对各峰型进行脂肪酸甲酯的鉴定[22]。
1.3.6 生物柴油性能分析 由巴夫藻的脂肪酸组成,根据公式计算生物柴油的皂化值(SV)、碘值(IV)、十六烷值(CN)、不饱和度(DU)、长链不饱和因子(LCSF)、冷滤点(CFPP)[23]。
1.4 数据分析
本次实验每处理设置3 个重复,对获得各数据使用IBM SPSS 23软件分析,选用一般线性模型、单因素分析和全模型对光周期、维生素浓度和二者的交互效应进行分析。利用Duncan 法对方差分析结果显著的因素进行多重比较。各分析指标以P<0.05 作为判断标准,并以不同字母表示比较组内差异的显著性[24]。
2 结果
2.1 光周期和维生素对巴夫藻生长的影响
在培养的14 d 内,巴夫藻在3 种光周期和3 个维生素浓度组成的9 种处理下都能生长(图1(a))。2 d后,1V 8 h∶16 h、4V 8 h∶16 h和8V 8 h∶16 h 三个处理的生长速度低于其他处理,其D660一直低于其他组,但这三个组之间差别不大(P>0.05)。其余各组中,24 h∶0 h 的3 个组培养8 d 后,D660比16 h∶8 h组略低。从培养结束收获的细胞密度看,细胞密度受光周期显著影响(P<0.05),而维生素浓度及“光周期×维生素浓度”交互作用对细胞密度影响不大(P>0.05)(图1(b)、表1)。在3 个维生素浓度下,8 h∶16 h 光周期处理的细胞密度都显著低于其他2个光周期,而16 h∶8 h 和24 h∶0 h 两个光周期的细胞密度差别不大,其中1V 16 h∶8 h处理的细胞密度为52.19×106mL-1,显著高于1V 8 h∶16 h 的45.98×106mL-1和1V 24 h∶0 h的49.58×106mL-1。因此,巴夫藻生长主要受光周期影响,短周期8 h∶16 h 不利于巴夫藻生长,长周期16 h∶8 h 和24 h∶0 h 尤其是16 h∶8 h有利于巴夫藻生长。
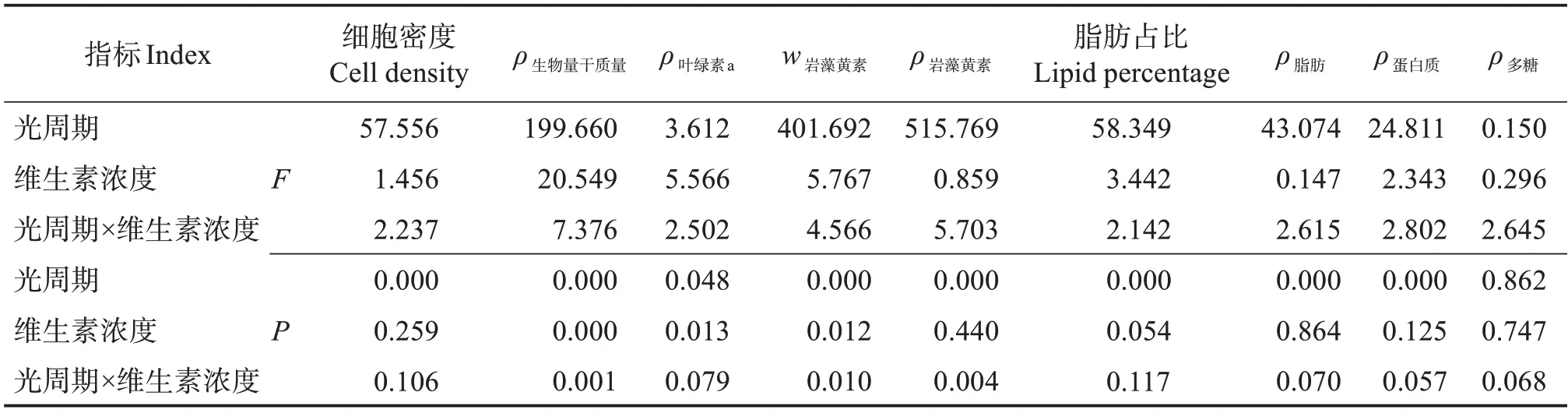
表1 光周期和维生素浓度对巴夫藻的影响双因素方差分析Table 1 Two-factor ANOVA analysis of effects of photoperiod and vitamin concentration on Pavlova sp.
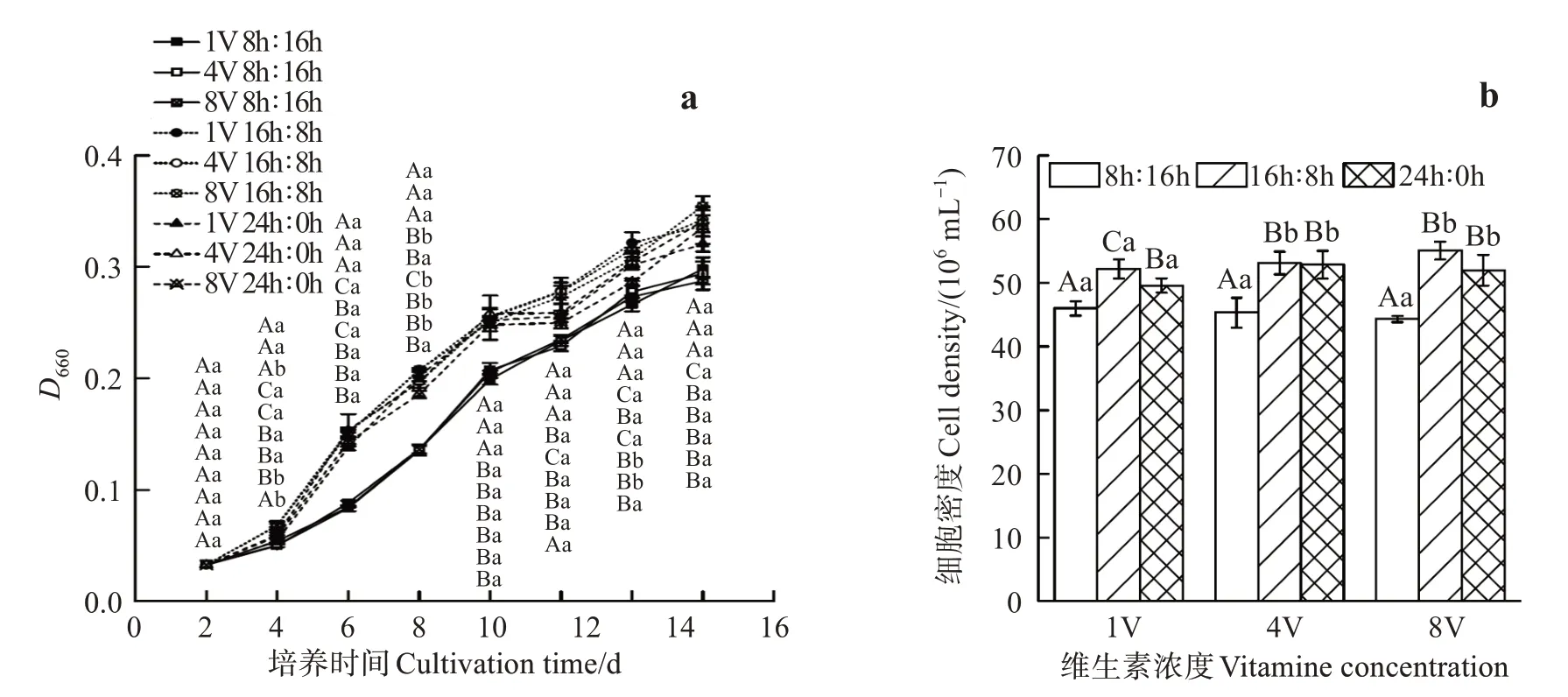
图1 光周期和维生素对巴夫藻生长和收获时细胞密度的影响Fig.1 Effects of photoperiod and vitamin concentration on growth and cell density of Pavlova sp.
2.2 光周期和维生素对巴夫藻生产能力的影响
由表1 可知,光周期对多糖以外的所有物质产量都有显著影响(P<0.05),而维生素浓度及“光周期×维生素浓度”交互作用对生物量干质量和岩藻黄素产量有显著影响(P<0.05)。
图2(a)表示巴夫藻培养14 d 后收获的生物量干质量情况。双因素方差分析结果表明,光周期、维生素浓度及“光周期×维生素浓度”交互作用都对干质量有显著影响(表1)。在3 种维生素浓度下,24 h∶0 h的干质量在3个光周期处理中都是最小的,其次是8 h∶16 h光周期,16 h∶8 h光周期的干质量是3个光周期中最高的,在3个维生素浓度下分别达到501.50、500.90、497.57 mg·L-1,因此16 h∶8 h 是生物量干质量生产的最佳光周期。在8 h∶16 h 光周期下,维生素浓度升高时干质量降低,从1V 维生素浓度时的450.60 mg·L-1,至8V 维生素浓度时干质量降到最低值419.37 mg·L-1。在16 h∶8 h 光周期下,干质量不随维生素浓度发生变化,当光周期为24 h∶0 h 时,也是8V 维生素浓度的干质量最低,只有361.80 mg·L-1。因此,1V 是最佳的维生素浓度,而1V 16 h∶8 h 的干质量又比1V 24 h∶0 h 提高了38.6%,所以生物量干质量生产的最佳条件是1V 16 h∶8 h。
如图2(b)所示,叶绿素a 含量既受维生素浓度影响,又受光周期影响。“光周期×维生素浓度”交互作用对叶绿素a 含量影响不显著(表1)。维生素浓度为1V 时,随光照时间延长,叶绿素a含量增高,最高1.18 mg·L-1出现在1V 24 h∶0 h。而在维生素浓度为4V 和8V 时,16 h∶8 h 光周期下的叶绿素a含量普遍高于8 h∶16 h 光周期下的含量,也高于24 h∶0 h 光周期下的叶绿素a 含量。光周期对叶绿素a含量的影响在维生素为8V时表现突出,16 h∶8 h光周期下叶绿素a 含量高,最高0.10 mg·L-1出现在8V 16 h∶8 h。
光周期、维生素浓度及“光周期×维生素浓度”交互作用对岩藻黄素生产影响显著(表1)。如图2(c)所示,在3种维生素浓度下,24 h∶0 h的岩藻黄素含量在3 个光周期中都是最小的,为2.7~3.0 mg·g-1,其次是16 h∶8 h 光周期,含量为3.8~4.2 mg·g-1,8 h∶16 h 光周期的岩藻黄素是3 个光周期中最高的,为5.5~6.2 mg·g-1,因此8 h∶16 h 是岩藻黄素积累的最佳光周期。岩藻黄素产量取决于生物量干质量和岩藻黄素含量,由于8 h∶16 h 光周期下岩藻黄素含量最高,24 h∶0 h 光周期下岩藻黄素最低,岩藻黄素产量同样也是这样的规律(图2(d))。在24 h∶0 h 光周期下岩藻黄素产量受维生素浓度影响不显著(P>0.05),但在8 h∶16 h 和16 h∶8 h 两个光周期下,维生素浓度为8V 时的岩藻黄素产量都比维生素浓度为1V 和4V 时高,因此8V 是岩藻黄素生产的最佳维生素浓度。8V 8 h∶16 h 的岩藻黄素产量达到2.60 mg·L-1,是所有处理中最高值,是1V 24 h∶0 h 的2.08倍。因此,岩藻黄素生产的最佳条件是8V 8 h∶16 h。
在脂肪生产方面,双因素方差分析结果表明,光周期对脂肪占比和脂肪产量有显著影响(P<0.05),维生素浓度及“光周期×维生素浓度”交互作用对脂肪占比和脂肪产量影响不大(P>0.05)(表1)。如图2(e)所示,在3 种维生素浓度下,24 h∶0 h 的脂肪占比均优于其他光周期,最高值为8V 24 h∶0 h,脂肪占比达到30.18%;16 h∶8 h 光周期的脂肪占比也比较高,而8 h∶16 h 光周期的脂肪占比最低。而脂肪产量的高低取决于生物量干质量和脂肪占比的乘积。如图2(f)所示,在3 种维生素浓度下,8 h∶16 h光周期的脂肪产量在3 个光周期中都是最低的,16 h∶8 h 和24 h∶0 h 两个光周期下的脂肪产量差别不显著,但在维生素浓度为8V 时,16 h∶8 h 的脂肪产量显著高于24 h∶0 h,达到123.87 mg·L-1,是所有处理中的最高值。
双因素方差分析结果表明,光周期显著影响蛋白质产量(P<0.05),维生素浓度及“光周期×维生素浓度”交互作用对蛋白质产量影响不大(P>0.05)(表1)。如图2(g)所示,在维生素浓度1V 和4V时,16 h∶8 h 的蛋白质产量均显著高于其他两个光周期(P<0.05)。1V 16 h∶8 h 的蛋白质产量达55.54 mg·L-1,是所有处理中的最高值,比1V 24 h∶0 h提高了30.8%。
巴夫藻多糖产量在17~27 mg·L-1之间,双因素方差分析结果表明,光周期、维生素浓度及“光周期×维生素浓度”交互作用对多糖产量影响都不显著(P>0.05)(表1,图2(h))。
2.3 光周期和维生素对巴夫藻脂肪酸组成和生物柴油质量的影响
巴夫藻主要含6种脂肪酸(表2),其中饱和脂肪酸有C14:0、C16:0和C18:0,不饱和脂肪酸有C16:1(n-7)、C18:1(n-9)和C20:5(n-3)(EPA)。含量最高的脂肪酸为C16:0,在总脂肪酸中的质量占比为36%~41%之间;其次是C16:1(n-7),占比在32%~35%之间;排在第三位的是占比在11%~13%之间的C18:1(n-9);EPA 作为唯一的多不饱和脂肪酸,占比在5%~15%之间。C14:0 和C18:0 两种饱和脂肪酸的占比之和不超过6%。双因素方差分析结果表明,光周期对巴夫藻6 种脂肪酸占比都有显著影响,维生素浓度对脂肪酸组成影响不大,“光周期×维生素浓度”交互作用只对脂肪酸C14:0占比有显著影响(表3)。

表2 光周期和维生素浓度对巴夫藻脂肪酸组成的影响Table 2 Effects of photoperiod and vitamin concentration on fatty acid composition of Pavlova sp.%
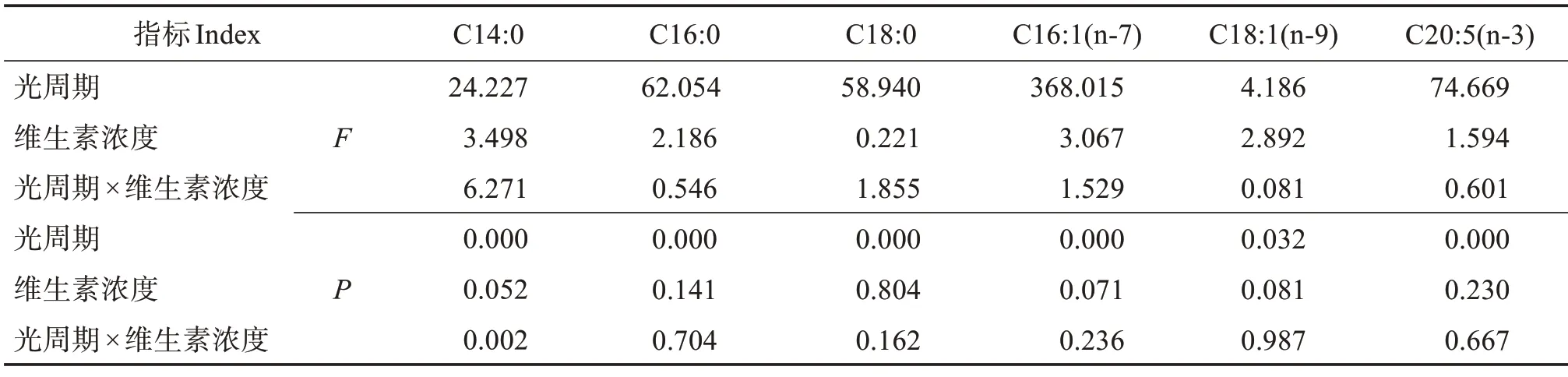
表3 光周期和维生素浓度对巴夫藻脂肪酸组成的影响双因素方差分析Table 3 Two-factor ANOVA analysis of photoperiod and vitamin concentration on fatty acid composition of Pavlova sp.
如表2所示,含量最高的饱和脂肪酸C16:0的含量受光周期显著影响,在3种维生素浓度下,24 h∶0 h的C16:0平均占比为40.66%,显著高于其他两个光周期。C16:1(n-7)的含量也受光周期的显著影响,在3种维生素浓度下,24 h∶0 h和16 h∶8 h光周期下的占比显著高于8 h∶16 h。在8V维生素浓度下,16 h∶8 h的C18:1(n-9)占比显著高于8 h∶16 h,但在3 个光周期下变动范围不足2%。在3 种维生素浓度下,EPA含量都符合8 h∶16 h>16 h∶8 h>24 h∶0 h的规律,且差异显著,即在8 h∶16 h光周期下最适合EPA积累。1V 8 h∶16 h 的EPA 占比为15.46%,是1V 24 h∶0 h的EPA 占比的2.28 倍。C18:0 含量符合24 h∶0 h>16 h∶8 h>8 h∶16 h 的规律,且差异显著。光周期对C14:0 占比有显著影响(P<0.05),维生素浓度对其占比无显著影响(P>0.05),“光周期×维生素浓度”交互作用对其占比有显著影响(P<0.05)。
双因素方差分析结果表明,光周期显著影响巴夫藻脂质生产的生物柴油性能各指标,维生素浓度及“光周期×维生素浓度”交互作用对生物柴油性能影响不大(表4)。
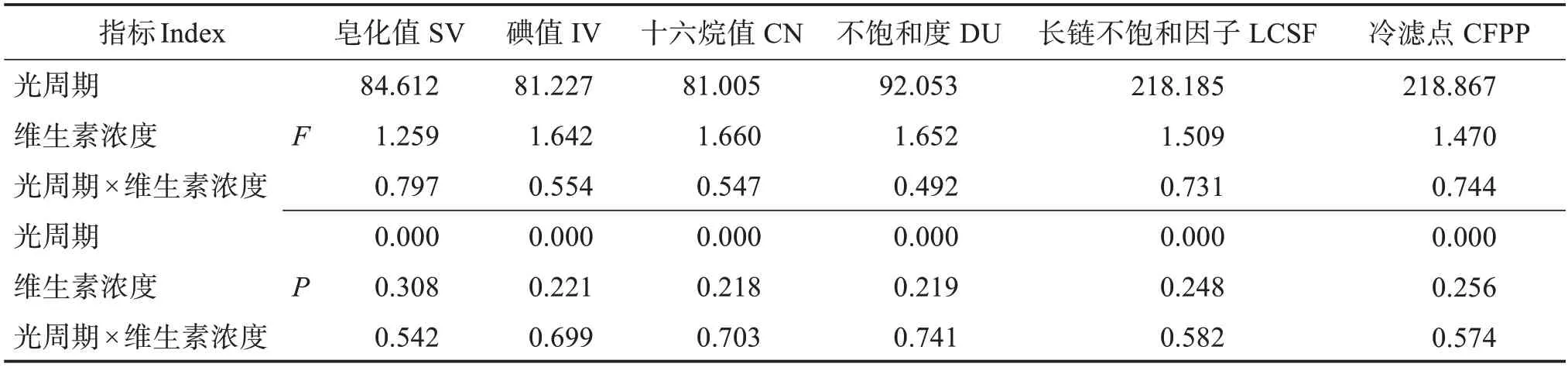
表4 光周期和维生素浓度对巴夫藻脂质的生物柴油性能的影响双因素方差分析Table 4 Two-factor ANOVA analysis of effects of photoperiod and vitamin concentration on the biodiesel performance of lipid derived from Pavlova sp.
巴夫藻脂质的生物柴油各指标见表5。生物柴油特性如十六烷值(CN)、运动黏度、氧化稳定性等,取决于原料微藻生物柴油生产过程中使用的脂肪酸。不同光周期和维生素浓度条件下培养的巴夫藻生物柴油的CN 值有明显区别,CN 值最低为44.55,出现在8 h∶16 h 光周期下,该光周期下的CN值显著低于其他两个光周期。而CN 值最高值为53.98,出现在24 h∶0 h 光周期,该光周期下的CN 值普遍高于其他两个光周期。不饱和度(DU)和碘值(IV)通常用来评价生物柴油的氧化稳定性,二者的趋势往往是一致的,都是其值越低,生物柴油的氧化稳定性越高[23]。DU 和IV 的值都符合24 h∶0 h <16 h∶8 h <8 h∶16 h的规律,最低值出现在24 h∶0 h光周期,其次是16 h∶8 h光周期,而DU和IV最高值出现在8 h∶16 h。冷滤点(CFPP)通常用于预测生物柴油在低温下的流动性能[23]。在9 种组合培养条件下的巴夫藻CFPP 都低于5 ℃,其中冷滤点最低值为-0.41 ℃,出现在8 h∶16 h 光周期下,该光周期下的CFPP 值显著低于其他两个光周期。16 h∶8 h光周期下CFPP 略高,最高值出现在24 h∶0 h 光周期。4V 24 h∶0 h 条件下生产的巴夫藻生物柴油CN值最高,DU 和IV 最低即氧化稳定性最好,但冷滤点略高即低温流动性相对差一些。
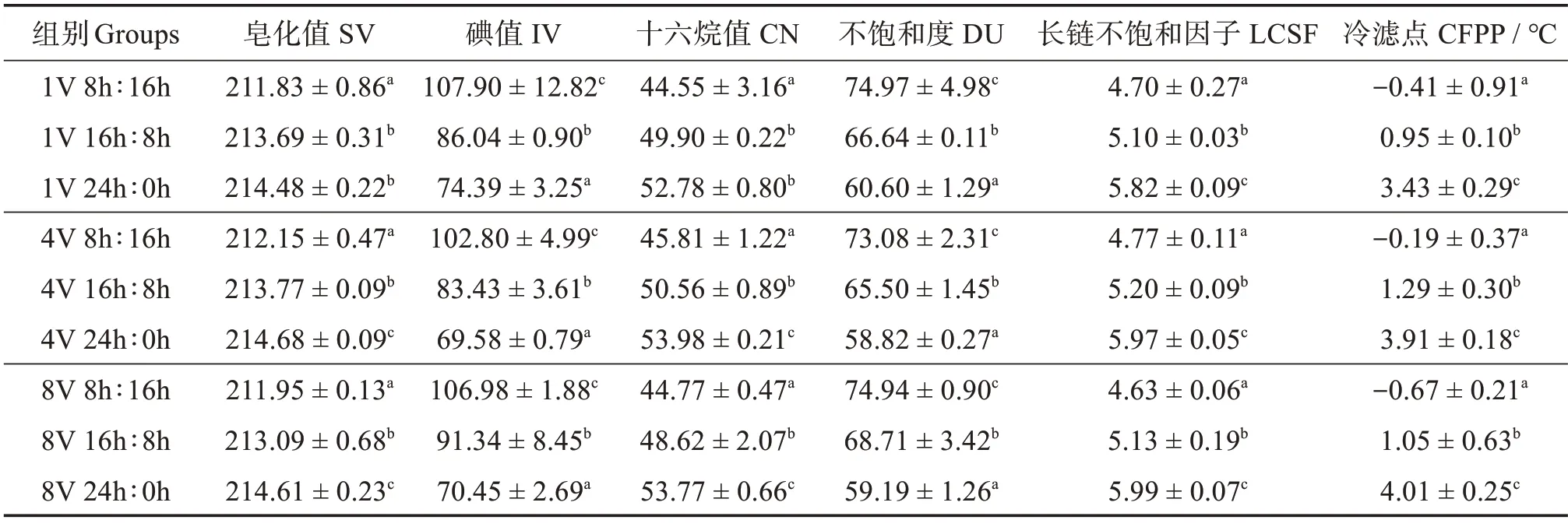
表5 光周期和维生素浓度对巴夫藻脂质的生物柴油性能的影响Table 5 Effects of photoperiod and vitamin concentration on the biodiesel performance of lipid derived from Pavlova sp.
3 讨论
3.1 光周期和维生素对巴夫藻生长和生物量积累的影响
微藻光合过程强烈依赖培养体系的光相关变量[13,22]。光合代谢由光期和暗期组成,光期产生的化学能用于暗期碳固定途径[25,12-13]。研究者们普遍认为光合过程需要光暗反应匹配才能高效形成光合产物[25,13,22]。适当的光/暗周期可提高光合系统的电子转移速度,并且在规模化培养中有助于调节微藻培养物的温度、降低设备运行的能源消耗[12-13,22]。本研究中,生产众多优质物质的巴夫藻,无论在生长还是物质生产能力方面都表现出明显的光周期效应。16 h∶8 h 处理组在生长方面表现出明显优势,培养14 d 结束时得到了最高的细胞密度、干质量、脂肪产量和蛋白质产量,而连续光照在生长和物质产量方面并没有优势。实际上,海洋浮游植物对光周期的响应具有很大的物种特异性。已有研究表明,赫氏圆石藻(Emiliania huxleyi)、细孔圆颗石藻(Cyclococcolithus leptoporus)、布朗葡萄藻(Botryococcus braunii)和斜生四链藻(Tetradesmus obliquus)在连续光照下生长较快,而球等鞭金藻(Isochrysis galbana)、金色藻(Chrysochromulinasp.)和远洋颗石藻(Coccolithus pelagicus)在连续光照下生长较差或完全不生长[34-35];两株颗石藻(Coccolithus)在14∶10光周期下生长较快,结合新绿藻(Neochloris conjuncta),陆生新绿藻(N.terrestris),德克萨斯新绿藻(N.texensis)在12 h∶12 h 光周期下生长更好,还有的微藻的生长速度受光周期的影响不显著[27]。造成微藻对光周期响应有差异的原因,可能与藻种的敏感性有关,与藻种自然分布的区域有关,还可能与微藻的细胞组成有关[26]。四列藻(Tetraselmis tetrathele)在光期为6~20 h 之间时藻细胞密度、叶绿素a和蛋白质含量等指标均达到较高水平,但在光期少于4 h 和超过20 h 时,则生长受到明显抑制[28]。因此,从种间特异性上看,巴夫藻应属于中长光照种类,光周期16 h∶8 h较适合巴夫藻生长,并生产较大生物量干质量。维生素供给在微藻规模培养中起着至关重要作用,然而,关于这些微量营养素对微藻生长、多样性和生产力的影响的资料很少[14-15]。本研究结果发现,维生素浓度对于巴夫藻生物量干质量有影响,当维生素浓度为1V 时,能获得最高的生物量干质量,而且维生素与光周期有交互作用,在1V 和长光照的16 h∶8 h光周期条件下,能使生物量干质量达到最高。
3.2 光周期和维生素对巴夫藻岩藻黄素生产的影响
本研究使用的巴夫藻能产生大量的岩藻黄素,质量分数可达6.21 mg·g-1,产量能达到2.60 mg·L-1。这与Kanamoto 等[5]筛选得到的最优巴夫藻株Pavlovasp.OPMS 30543 在f/2 培养基中获得的岩藻黄素质量分数1.48 mg·g-1、产量0.73 mg·L-1相比,分别是其4.20 倍和3.56 倍。造成这两株巴夫藻产量差距的原因,可能与培养时长不同有关,分别是14 d 和7 d,有可能Pavlovasp.OPMS 30543 岩藻黄素的积累还不够充分;还与光周期不同有关,分别是8 h∶16 h 和12 h∶12 h,本研究已证明短光照有利于岩藻黄素积累;还与维生素浓度相关,分别是8倍和1 倍浓度,而高浓度维生素有利于岩藻黄素积累;藻株本身的差异也至关重要。因此,本研究的巴夫藻是生产岩藻黄素的优质藻株。因为岩藻黄素能够捕获光能并保护光合器官,光照条件被认为是影响微藻合成岩藻黄素的最重要因素[25]。然而,光照条件对于岩藻黄素积累的影响研究主要集中在光强和光质方面[29]。本研究发现,光周期对巴夫藻的色素组成包括叶绿素a和岩藻黄素含量均有显著影响。维生素浓度为1V 时,24 h∶0 h 时叶绿素a含量最高;但是当维生素浓度升高时,16 h∶8 h 光周期下叶绿素a 含量最高。岩藻黄素和叶绿素a 共同组成光捕获复合物,但二者的化学结构不同,吸收的光谱不同,前者吸收蓝光和绿光,后者吸收红光和蓝光[29]。岩藻黄素与叶绿素a 对光周期的反应也不同,始终是光照时间最短的8 h∶16 h 光周期下含量最高。维生素浓度对于巴夫藻的岩藻黄素积累有影响,提高维生素浓度至8V 时,能提高岩藻黄素含量。而且维生素与光周期有交互作用,在8V 和短光照的8 h∶16 h 光周期条件下,能使岩藻黄素含量达到最高。因此,8V 8 h∶16 h 是岩藻黄素积累的最佳条件。
3.3 光周期和维生素对巴夫藻脂肪酸组成和生物柴油质量的影响
光周期调控能提高巴夫藻的脂质产量并对脂肪酸组成和生物柴油质量产生影响。巴夫藻脂质含量在光周期24 h∶0 h下最高,能达到30.18%;脂质产量在8V 16 h∶8 h获得最大值,达到123.87 mg·L-1。巴夫藻中含量最丰富的两种脂肪酸C16:0 和C16:1(n-7),在24 h∶0 h光周期下含量都是最高值。脂肪酸组成的变动引起生物柴油质量的变化,高含量的饱和脂肪酸(SFA)导致CN的增加,充足的单不饱和脂肪酸(MUFA)显著改善低温下的流动性能[30-31]。本研究发现,巴夫藻的SFA+MUFA 占总脂肪酸质量分数为80%~90%,是优质的生物柴油燃料生产的原料[30-31]。生物柴油指标中,CN 值越高,生物柴油的点火性能越好,发动机性能越好[18,31]。欧洲标准EN 14214 和ASTM 美国标准D6751 的生物柴油参考标准分别为CN >51 和CN >47[18]。反映生物柴油氧化稳定性的DU 和IV,均基于单不饱和脂肪酸和多不饱和脂肪酸的比例,DU 和IV 越低则氧化稳定性越好[23,32]。本研究中,4V 24 h∶0 h 条件下生产的巴夫藻生物柴油,CN 值最高可达53.98,其CN 值大于51,优于欧洲标准EN 14214 和ASTM 美国标准D6751,说明该条件下生产的巴夫藻可以作为生物柴油的直接原料,同时其DU 和IV 最低,即氧化稳定性也是最优的。另外,大量研究表明Pavlovasp.还可以产生大量的C20:5(n-3)(EPA)[3,6,33]。本研究结果发现巴夫藻脂肪酸主要由16:0、C16:1(n-7)组成,并含有大量的C20:5(n-3),这与燕燕等[33]报道的绿色巴夫藻的脂肪酸组成相似。而且光周期会影响EPA 的积累,在8 h∶16 h 光周期下最适合积累EPA,在脂肪酸中占比最高能达到15.46%。与光周期相比,维生素浓度对于巴夫藻的脂肪生产影响不大,脂质含量、脂肪酸组成及EPA 含量均不受维生素浓度影响。
综上所述,巴夫藻作为岩藻黄素、EPA和优质生物柴油的生产者,在规模化培养中根据产品应用要求,通过合理调控光周期和维生素浓度,可进一步提高其物质生产效率和产品品质。
猜你喜欢
遗传(2023年9期)2023-09-25
农业科技通讯(2023年1期)2023-02-12
云南化工(2021年10期)2021-12-21
海峡姐妹(2019年8期)2019-09-03
天然产物研究与开发(2019年1期)2019-03-01
集美大学学报(自然科学版)(2018年1期)2018-03-21
西部皮革(2018年11期)2018-02-15
天然产物研究与开发(2018年1期)2018-02-02
中成药(2017年12期)2018-01-19
少儿科学周刊·少年版(2017年7期)2017-09-29
