
抗菌肽对异育银鲫生长、免疫和抗氧化性能的影响
2024-01-11 11:06郭益红赵宇凡朱文婷赵永志宣雄智李文嘉
饲料工业 2023年24期
■ 郭益红 赵宇凡 范 勇 朱文婷 赵永志 宣雄智 李文嘉
(1.苏州农业职业技术学院,江苏苏州 215008;2.张家港市大新镇农技推广服务中心,江苏张家港 215636;3.广东海纳川生物科技有限公司,广东佛山 528515;4.河南省农业科学院畜牧研究所,河南郑州 450002)
异育银鲫具有食性广、生长速度快、抗病力强等众多优点,且肉质鲜美,营养价值高,经济效益好,在我国淡水养殖中占据着十分重要的位置[1]。尤其是新品种“中科5 号”,饲料转化率高、耐低氧、成活率高等特点更加明显。但是随着高密度、集约化养殖模式的推广导致异育银鲫病害问题严重,在各种细菌性疾病方面尤为突出,养殖户为减少经济损失,擅自盲目过度使用抗生素致使鱼体药物残留超标,大量未被鱼体吸收的抗生素(约80%)随着鱼体排泄物将进入水体或沉降聚集于底泥,导致抗生素耐药性在环境中传播扩散[2]。鱼药和饲料添加剂等投入品的不规范使用已成为异育银鲫质量安全风险的主要来源之一,开发安全有效、无污染的抗生素替代产品受到业内学者和主产区渔业行政主管部门的高度关注。
抗菌肽是由20~60 个氨基酸残基组成的碱性多肽,广泛存在于大自然中,具有无残留、广谱抗菌、不易产生耐药性等特点,天然抗菌肽或人工合成抗菌肽具有替代抗生素的可行性[3]。在水产养殖领域,抗菌肽的研究主要集中于肉食性鱼类,如石斑鱼[4-5]、红姑鱼[6]等,在植食性鱼类异育银鲫中的应用效果和适宜用量尚不明确。试验以异育银鲫为研究对象,从生长性能、消化吸收能力、免疫和抗氧化性能等多个方面评估抗菌肽对异育银鲫生长、生理的影响,为抗菌肽在异育银鲫养殖生产中的合理使用提供理论依据。
1 材料方法
1.1 试验材料
试验用异育银鲫购自湖州菱湖水产苗种场,抗菌肽(柞蚕代谢物抗菌肽,活性单位20 000 U/g,分子量约为4 kD,商品名“富利泰”)购自广东海纳川生物科技股份有限公司,饲料原料购自淮安正昌饲料有限公司。
1.2 饲料配方设计
以鱼粉、豆粕、菜粕等为主要饲料原料,以抗菌肽作为饲料添加剂,所有原料经粉碎后过60 目筛,微量组分采用逐级扩大混匀法加入,与水混匀后用制粒机(HKj200 章丘市华祥颗粒机械有限公司)制成直径为1.5 mm 的颗粒饲料,室温风干,用包装袋密封后放入-20 ℃冰箱备用,基础饲料配方及营养水平如表1所示。试验设计共分5 组,分别为不含抗菌肽的对照组(CK 组)、添加50 mg/kg(D1 组)、75 mg/kg(D2 组)、100 mg/kg(D3 组)和125 mg/kg(D4 组)抗菌肽的试验组。
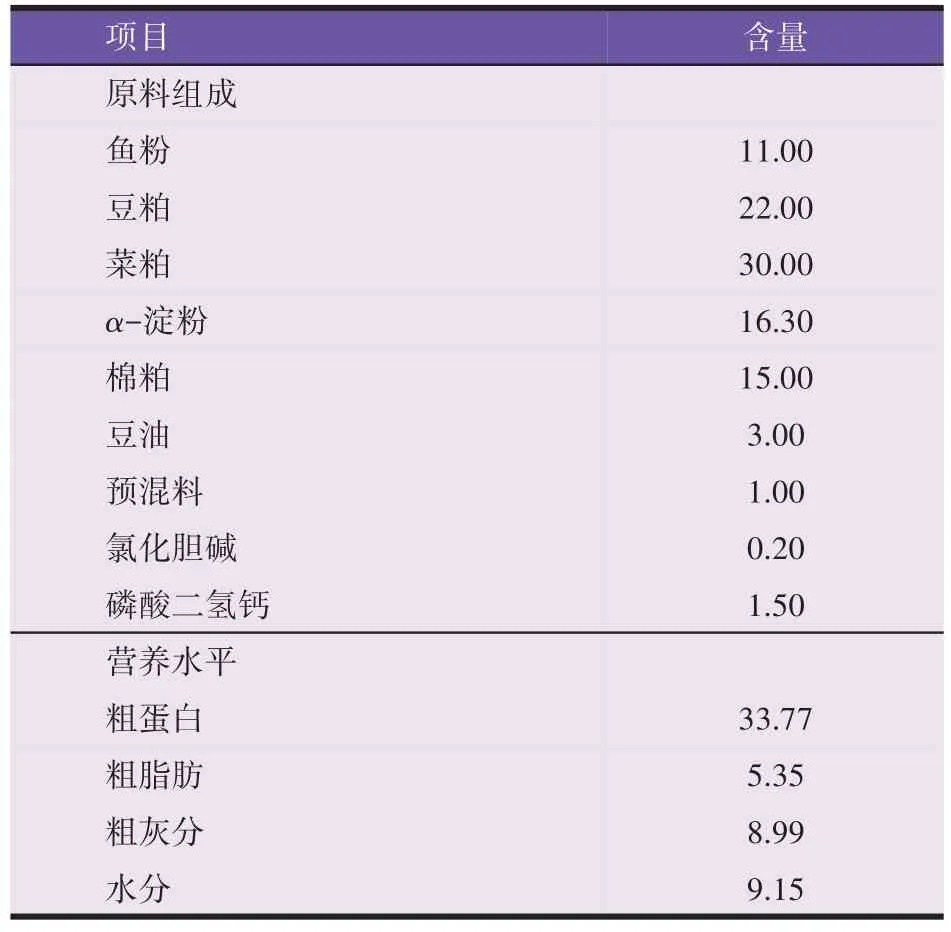
表1 基础饲料配方及营养水平(干物质基础,%)
1.3 饲养管理
试验于2021年5月—7月在兴化宏伟特种水产养殖场完成,试验共56 d。选取健康且规格一致的异育银鲫幼鱼375 尾[平均初始体重(9.40±0.2) g]用对照组饲料在池塘网箱内暂养驯化14 d。试验开始前,幼鱼禁食24 h,用40 mg/L 的丁香酚麻醉后称重,并放入长方形养殖水箱(400 L),每个试验组3 个平行,每个平行1 个水箱,25 尾。适量连续充气,使水体溶解氧≥4 mg/L,试验用水为经充分曝气48 h 的自来水,试验期间每天监测,水温为21~25 ℃,pH 7.1~7.5,氨氮≤0.2 mg/L,亚硝酸盐≤0.2 mg/L,每天采用连续性饱食投喂2 次(08:30、16:30),以试验鱼不再抢食为准,投喂结束后1.5 h 用虹吸法清除水箱底部残饵与粪便,监测水质,保持养殖水深和水质稳定,记录投饲量,饲养周期为56 d。
1.4 样品采集
饲养试验结束后,试验鱼禁食24 h,用40 mg/L丁香酚麻醉后将试验鱼全部捞出,计数并逐尾称重。每个养殖水箱随机挑取6 尾,其中3 尾称重后捣碎并混合、烘干(105 ℃),然后放入-20 ℃冰箱中用于分析全鱼成分。其余的鱼用注射器从尾静脉采血,低温离心(2 000 r/min,10 min,4 ℃)分离取上层血清。收集到的血清按平行混合后分装于0.5 ml 离心管,置于超低温冰箱(-80 ℃)保存备用。将上述抽完血的试验鱼分离肝脏、前肠,分别称内脏团和肝脏重量,用于分析形态学指标,去鱼皮分离背部侧线以上肌肉,所取各组织样品装袋后经液氮速冻保存于-80 ℃中用于试验后期测定各种相关酶活。
1.5 指标测定
饲料和全鱼样品的粗蛋白质、粗脂肪、粗灰分、水分含量,背肌的粗蛋白、粗脂肪、水分含量,肝脏的粗脂肪和水分含量均按国标方法测定。全鱼和饲料蛋白质成分通过凯氏定氮仪(SKD-1000,上海沛欧分析仪器有限公司)测定,粗脂肪用索氏抽提法;灰分使用马弗炉(80-10TP,上海慧泰仪器制造有限公司)在550 ℃下烘6 h 测定;水分使用烘箱(GZX-9030MBE,上海博迅实业有限公司医疗设备厂)105 ℃过夜测定;肝脏总抗氧化力(T-AOC)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和丙二醛(MDA),前肠胰蛋白酶、脂肪酶、淀粉酶,血清溶菌酶(LZM)、总蛋白(TP)的测定试剂盒购自南京建成生物工程研究所有限公司。
计算试验鱼的增重率、饲料转化率、摄食率、成活率指标。
式中:Wt——试验结束时鱼体重(g);
W0——试验初始鱼体重(g);
t——试验时间(d);
Wd——死亡鱼体重(g);
I——饲料总投喂量(g)。
1.6 数据处理与分析
采用SPSS 22 和Excel 2016 软件对试验数据进行统计分析,采用单因素方差分析法(one-way ANOVA,LSD),用Duncan’s 多重比较法检验差异显著性,结果均以“平均值±标准误”来表示,P<0.05表示差异显著。
2 结果与分析
2.1 抗菌肽添加量对异育银鲫生长和饲料利用的影响
由表2 可知,抗菌肽添加量对异育银鲫增重率、摄食率和饲料利用率均无显著影响(P>0.05)。
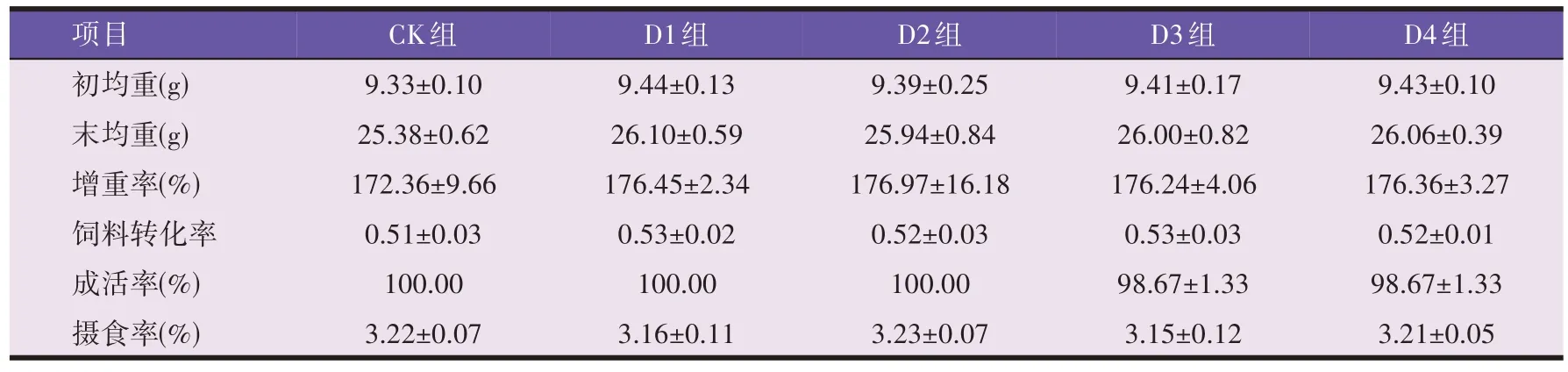
表2 异育银鲫的生长、饲料利用率及存活状况(n=3)
2.2 抗菌肽添加量对异育银鲫体组成的影响
由表3 可知,抗菌肽添加量对异育银鲫全鱼各成分含量无显著影响(P>0.05),随着抗菌肽添加量的增加,背肌粗脂肪含量呈下降趋势,与CK 组相比,D4组异育银鲫背肌和肝脏粗脂肪含量均显著下降(P<0.05)。
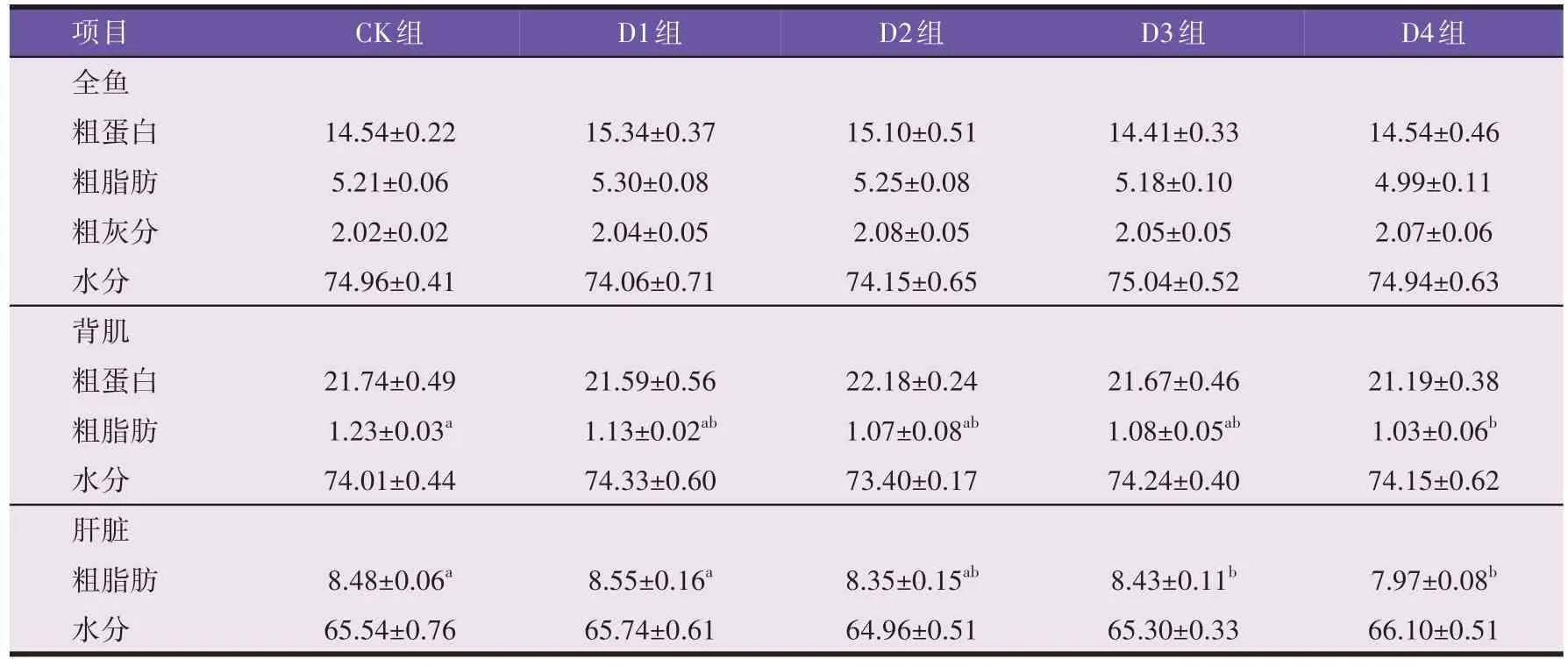
表3 异育银鲫的全鱼、背肌和肝脏成分(湿重,n=3,%)
2.3 抗菌肽添加量对异育银鲫消化吸收酶活性的影响
由表4 可知,抗菌肽添加量均可使异育银鲫前肠胰蛋白酶活性有所提高,与CK 组相比,D2 组异育银鲫该酶活性显著升高(P<0.05)。

表4 异育银鲫前肠消化酶活性(n=3)
2.4 抗菌肽添加量对异育银鲫抗氧化相关酶活性的影响
由表5 可知,与CK 组相比,D2、D3 组异育银鲫肝脏SOD 活性和T-AOC 显著升高(P<0.05),D4 组异育银鲫肝脏CAT 活性显著下降(P<0.05);不同添加量的抗菌肽均可降低肝脏MDA 含量,但差异并不显著(P>0.05)。

表5 异育银鲫肝脏抗氧化酶活性及丙二醛含量(n=3)
2.5 抗菌肽添加量对异育银鲫血清免疫指标的影响
由表6 可知,与CK 组相比,不同添加量的抗菌肽均可降低异育银鲫血清TP 含量,但差异不显著(P>0.05);与CK 组相比,D2、D3 组异育银鲫血清LZM 活性显著升高(P<0.05)。

表6 异育银鲫的血清指标(n=3)
3 讨论
3.1 抗菌肽添加量对异育银鲫生长和饲料利用的影响
多数研究表明,在饲料中添加适当浓度的抗菌肽可提高养殖动物的生长性能和饲料利用率[4-10]。本试验条件下,不同的抗菌肽添加量对异育银鲫增重率和饲料利用率均无显著影响,可能是由于饲料中抗菌肽添加量占比较低,所以对异育银鲫生长影响细微,同时也表明抗菌肽作为饲料添加剂对水产动物生长性能的影响可能与水产动物的种类、生长发育状况、抗菌肽的使用剂量、养殖环境等多种因素有关。
3.2 抗菌肽添加量对异育银鲫体组成的影响
鱼的肌肉成分组成受品种、饲料、养殖环境等多种因素的影响,其中饲料和生长环境是重要的影响因素。饲喂高蛋白饲料会使鱼肌肉脂肪含量升高,而过多的脂肪沉积将使得鱼肉产生“油腻感”,降低了鱼肉的口感[8]。张莉等[6]研究表明加州鲈全鱼粗脂肪含量会随抗菌肽添加量的升高而下降,另有研究表明在饲料中添加Surfactin 抗菌肽能够降低斜带石斑鱼全鱼和肝脏的脂肪含量[12-13]。本试验条件下,随着抗菌肽添加量的升高,异育银鲫背肌粗脂肪含量呈下降趋势,当添加量为125 mg/kg(D4组)时,异育银鲫背肌和肝脏粗脂肪含量均显著下降,可能是由于抗菌肽可以加强肌肉脂肪代谢,降低脂肪酸合成速度,减少养殖动物体脂沉积[11],也有研究认为抗菌肽降低肝脏粗脂肪的作用与其乳化作用有关[5],值得注意的是,肌肉粗脂肪含量的下降有助于提高鱼肉口感[12]。
3.3 抗菌肽添加量对异育银鲫消化酶活性的影响
消化酶活性可以反映鱼类的基本消化生理特征,常用于衡量鱼类对饲料营养成分的消化吸收情况,消化酶活性升高有利于营养物质的消化、吸收和利用[13-15]。有研究表明,在饲料中添加抗菌肽可以提高吉富罗非鱼[16]、斜带石斑鱼[5]肠道蛋白酶和脂肪酶的活性。本试验条件下,不同的抗菌肽添加量均可使异育银鲫前肠胰蛋白酶活性有所提高,且当添加量为75 mg/kg 时,该酶活性显著升高,这可能是由于抗菌肽能促进异育银鲫肠道发育,也有研究表明抗菌肽可能通过降低肠道条件致病菌的数量,增加了肠道益生菌群的营养物质和生长空间来改善肠道微环境[17-18],从而提升消化酶活性。
3.4 抗菌肽添加量对异育银鲫抗氧化相关酶活性的影响
总抗氧化力是用于衡量机体抗氧化能力的综合性指标,组成鱼体抗氧化系统的酶主要有超氧化物歧化酶、过氧化氢酶和谷胱甘肽过氧化物酶。超氧化物歧化酶能清除体内超氧阴离子自由基,过氧化氢酶可以将超氧化物歧化酶催化活性氧自由基所产生的H2O2转化为水和氧气[19]。丙二醛是脂质过氧化终产物的指标,间接反映细胞受氧化损伤的程度[9],有研究表明饲料中添加0.015%抗菌肽可以提高锦鲤血清溶菌酶和过氧化氢酶的活性[20],增强鲤鱼补体旁路途径活性[21],添加0.015%和0.02%的天蚕素抗菌肽可以提高湘云鲫血清过氧化氢酶活性,降低丙二醛的含量[7]。还有研究表明同一蛋白水平下增加抗菌肽可以降低赤点石斑鱼肝脏丙二醛含量,增加肝脏过氧化物酶和过氧化氢酶的活性[4]。本试验条件下,与CK组相比,在饲料中添加75 mg/kg 和100 mg/kg 的抗菌肽可以显著提高异育银鲫肝脏超氧化物歧化酶活性和总抗氧化能力,降低肝脏丙二醛含量,表明抗菌肽可以诱导异育银鲫抗氧化系统的激活,提高机体抗氧化能力,降低肠道内的自由基水平,从而减轻肠道的氧化损伤。抗菌肽提高鱼体抗氧化能力的机制可能是由于其具有抗菌作用,能抑制肠道病原菌的繁殖,减少内毒素积累诱发的氧衍生为自由基,引起脂质过氧化损伤[10]。
另有研究表明在吉富罗非鱼饲料中添加50 mg/kg抗菌肽或在黄颡鱼饲料中添加100 mg/kg 抗菌肽对肝脏或血清超氧化物歧化酶和血清溶菌酶活性有显著的负面影响[22-23],本试验条件下,当抗菌肽添加量为125 mg/kg 时,异育银鲫肝脏超氧化物歧化酶活性有所下降,过氧化氢酶的活性则显著下降,有研究表明在饲料中添加10 mg/kg 与30 mg/kg 抗菌肽能使黄颡鱼血清过氧化氢酶活性提高约50%[23],说明与超氧化物歧化酶相比,肝脏过氧化氢酶活性可能更易受抗菌肽剂量的影响,同时也说明高剂量的抗菌肽可能对鱼抗氧化性能存在不利影响,具体机制有待进一步研究。
3.5 抗菌肽添加量对异育银鲫血清免疫指标的影响
抗菌肽在体内除了能直接杀死病原菌,更多的是作为免疫效应因子来激活宿主的免疫防御系统,从而提升免疫功能来消除感染,保护宿主免受病原微生物的侵害[24]。鱼类血清生化指标能够反映鱼体的生理代谢和免疫状况[15]。血清总蛋白主要包括白蛋白和球蛋白,是衡量体液免疫水平的重要指标[25]。本试验条件下,不同的抗菌肽添加量均降低了异育银鲫血清总蛋白含量,具体机制有待进一步研究。
溶菌酶常用来评估鱼类非特异性免疫性能。溶菌酶是水产动物体内的一种重要非特异性防御因子,可以催化水解细菌细胞壁而导致细菌溶解死亡[25]。有研究表明抗菌肽可以被养殖动物血淋巴吸收,从而诱导溶菌酶和酸性磷酸酶活性的增加,进而提高养殖动物的免疫力[26]。本试验条件下,与对照组相比,当抗菌肽的添加量为75 mg/kg和100 mg/kg时,异育银鲫血清溶菌酶活性均显著升高,表明抗菌肽能一定程度提高异育银鲫的非特异性免疫能力,这与Surfactin 抗菌肽在斜带石斑鱼[9]、吉富罗非鱼[27]的研究结果类似。
4 结论
本试验条件下,在异育银鲫饲料中添加抗菌肽可以降低鱼体肌肉脂肪含量并提升鱼体免疫和抗氧化性能,抗菌肽添加量为75 mg/kg 和100 mg/kg 时效果较佳。
猜你喜欢
现代畜牧科技(2021年9期)2021-10-13
科学养鱼(2020年5期)2020-12-20
食品工业科技(2020年24期)2020-12-09
当代水产(2019年8期)2019-10-12
绿色科技(2016年20期)2016-12-27
广东饲料(2016年5期)2016-12-01
国外医药(抗生素分册)(2016年5期)2016-07-12
湖北农业科学(2014年21期)2014-12-22
特产研究(2014年4期)2014-04-10
