
发育期兔眼外肌的组织学特点
2024-01-25 11:44楼江涛彭一一王春晓许梅萍余新平
中华眼视光学与视觉科学杂志 2024年1期
楼江涛 彭一一 王春晓 许梅萍 余新平
作者单位:1温州医科大学附属眼视光医院 省部共建眼视光学和视觉科学国家重点实验室,温州 325027;2中山大学中山眼科中心 眼科学国家重点实验室 广东省眼科视觉科学重点实验室,广州 510060 (彭一一,现工作于华中科技大学同济医学院附属武汉儿童医院 武汉市妇幼保健院;王春晓,现工作于洛阳市第三人民医院)
斜视发病机制复杂,近年研究表明眼外肌的异常可能参与了斜视的发生和发展,如斜视患者存在眼外肌附着点异常,Pulley结构的异常等改变。斜视矫正手术时获取的眼外肌病理研究发现眼外肌存在肌肉萎缩、肌纤维减少而间质增多,胶原纤维组织增生[1]的情况;电子显微镜下还发现眼外肌的线粒体结构和功能异常[2]。也有研究发现斜视患者的活化卫星细胞数量存在异常[3]。
然而,既往研究混杂了不同年龄段患者的眼外肌,动物实验证实出生后早期的眼外肌存在一个涉及肌肉形态的发育[4]、神经的发育完善[5]和肌球蛋白重链的构成重整[6]等复杂发育过程。但这些研究未明确探究幼年至青春期的眼外肌特点;且眼外肌的分层不够详细(眼外肌的眶层可进一步分成内、外2 层[7]);同时这些研究主要以大/小鼠为研究对象,由于其眼外肌较小,限制了其在斜视研究领域的进一步应用,如眼外肌手术相关的研究。相较之下,兔眼外肌能弥补以上不足。
斜视手术治疗的主要实施对象为幼年和青春期阶段的人群,目前对该时期眼外肌的发育特点,尤其是卫星细胞数量等情况尚不明确。因此本研究拟以幼年和青春期的新西兰大白兔为研究对象,探究不同发育阶段的兔眼外肌的组织学是否存在不同。
1 材料与方法
1.1 实验动物及分组
分别选取2周龄、4周龄、3月龄新西兰大白兔各4只作为2周龄、4周龄组和3月龄组,雌雄不限。兔子购自杭州余杭科联兔业专业合作社,于16~20 ℃、50%湿度环境下,使用兔专用饲料饲养,自由进水。实验方案经温州医科大学动物伦理委员会批准(wydw2021-0309)。
1.2 眼外肌的获取及切片制备
对兔子予以过量麻醉(赛拉嗪,20 mg/kg)安乐死,颈动脉放血后,取完整长度的右眼上直肌,0.9%氯化钠溶液冲洗,组织固定液中固定24 h。
眼外肌常规脱水,石蜡包埋,沿垂直肌纤维走行方向连续切片(2 μm),取6 张中腹部的切片,1张用于HE染色,剩余5张用于免疫组织化学染色。
1.3 试剂
MyHC1(fast-2X亚型)、MyHC14(slow-tonic亚型)购自武汉三鹰生物技术有限公司;MyHC3(emb亚型)购自英国Abcam公司;MyHC8(neo亚型)购自美国Bio-Techne公司;MyoD购自武汉三鹰生物技术有限公司。
1.4 苏木精-伊红染色(Hematoxylin-eosin staining,HE 染色)
将切片脱蜡至水;苏木素染色(苏木素染色5 min,盐酸水溶液分化2 s,氨水水溶液返蓝15~30 s,水洗);伊红染色(95%乙醇脱水,伊红染液染色5~8 s);切片脱水,中性树胶封片。切片扫描。
1.5 免疫组织化学染色
切片脱蜡至水;置于柠檬酸抗原修复液(pH=6.0)中,于微波炉内进行抗原修复;加入3%H2O2阻断内源性过氧化物酶;加入封闭血清;加入一抗(一抗浓度:MyHC1—1:3 000,MyHC3—1:300,MyHC8—1:200,MyHC14—1:2 000,MyoD—1:200),4℃孵育过夜;加入二抗;DAB显色;复染细胞核;脱水封片。切片扫描。DAB显色后细胞呈现棕色表示阳性。
1.6 研究指标
1.6.1 肌纤维平均横截面积 每张切片随机选择至少2个包含眶、球层的高倍视野,用图像处理软件Image J分析肌纤维的横截面积。
1.6.2 肌球蛋白重链(Myosin heavy chain, MyHC)表达情况 MyHC根据出现表达的先后顺序大体上可分成未成熟亚型(emb亚型、neo亚型)和成熟亚型(如fast、slow亚型,eom-specific亚型等)。研究认为MyHC的构成决定了肌肉的收缩性质和抗疲劳特性[8]。本研究为探究MyHC表达随年龄的改变,选取了emb和neo 2种未成熟亚型以及快纤维亚型(fast-2X)和慢纤维亚型(slow-tonic)4种代表性亚型进行研究。每张切片随机选择至少2 个高倍视野,计算表达各MyHC的肌纤维占视野内总肌纤维的比例。
1.6.3 活化卫星细胞的比例 每张切片随机选择至少2个高倍视野,计算MyoD+细胞核数量与视野内肌纤维数量的比值作为计数活化卫星细胞数量的指标[9]。
在分析MyHC的构成及活化卫星细胞比例时,本研究参考Lucas的实验方法[7],将眶层进一步分为外眶层和内眶层。
1.7 统计学方法
实验研究。应用SPSS 26.0或GraphPad Prism 8.4.3进行数据统计分析。对于正态分布的数据(经Shapiro-Wilk检验,P≥0.05),以±s表示;非正态分布则以M(QR)表示。符合正态分布且方差齐的数据,采用单因素方差分析,非正态分布数据则采用Kruskal-Wallis非参数检验,用Bonferroni法进行事后检验。以P<0.05为差异有统计学意义。
2 结果
2.1 组织形态改变
2 周龄时,内、外眶层之间没有明显的界限,肌束形态不够明显。随着年龄的增加,内、外眶层与球层之间的界限逐渐清晰。其中,外眶层的延续性一直较好,而内眶层和球层则逐渐形成由肌外膜包裹的肌束(图1)。肌纤维的平均横截面积随年龄的增加而增加,2周龄组、4周龄组、3月龄组分别为118.50(100.55)、151.54(190.32)、207.56(282.96) μm2(P<0.001)。
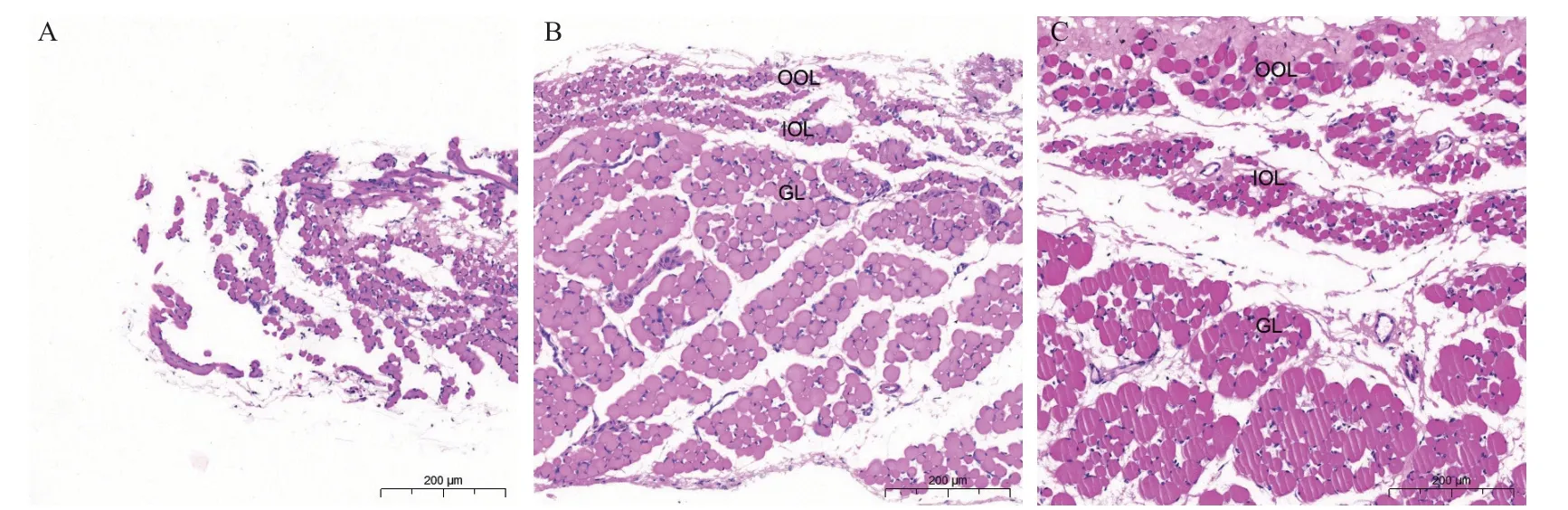
图1.兔眼外肌的组织形态改变Figure 1.Morphological changes of extraocular muscles in rabbits
2.2 MyHC表达的变化
MyHC1 主要在内眶层和球层表达,在外眶层表达较少(图2A-C),但其在内眶层(P=0.250)和球层(P=0.340)的表达组间差异无统计学意义,在外眶层的表达组间差异有统计学意义(P<0.001)。采用Bonferroni法校正显著性水平的事后检验发现,2周龄组的表达水平较4 周龄组(P=0.011)和3 月龄组(P<0.001)的表达水平低。4周龄组和3月龄组的表达水平差异无统计学意义(P>0.05)。见表1。
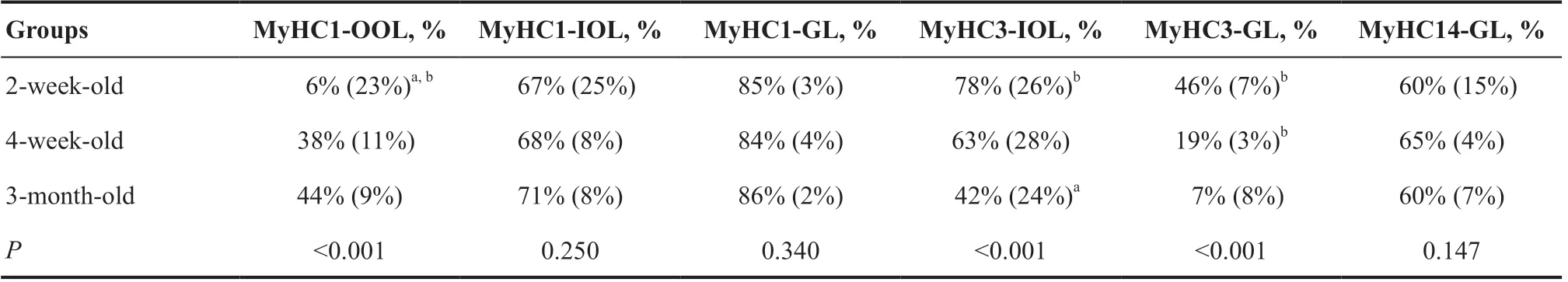
表1.发育期兔眼外肌的MyHC的表达情况Table 1.The expression of MyHC in developing rabbits' extraocular muscles

图2.兔眼外肌中MyHC1、MyHC14表达的变化Figure 2.Changes of the expression of MyHC1, MyHC14 of extraocular muscles in rabbits
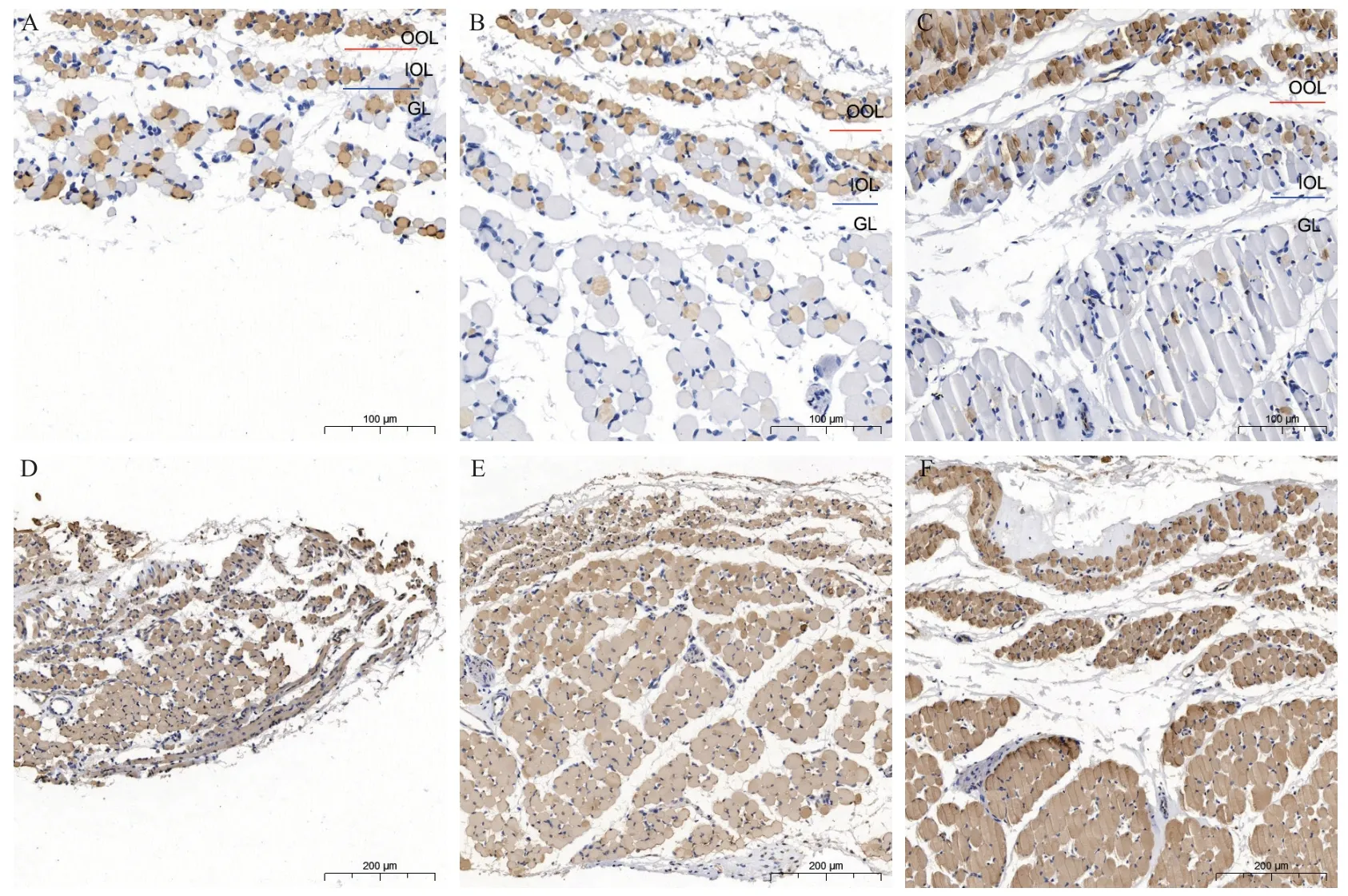
图3.兔眼外肌中MyHC3、MyHC8表达的变化Figure 3.Changes of the expression of MyHC3, MyHC8 of extraocular muscles in rabbits
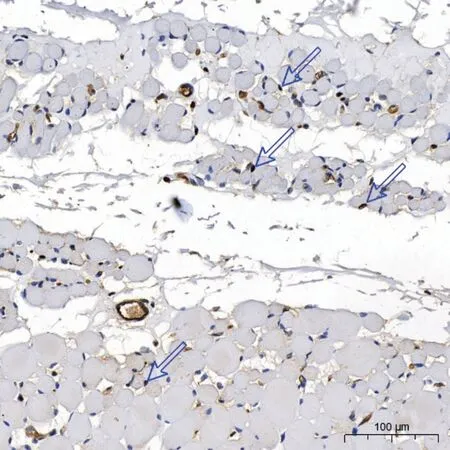
图4.3月龄组兔眼外肌中典型的活化卫星细胞Figure 4.Typical activated satellite cells in extraocular muscle in 3-month-old group of rabbits
MyHC14 主要在球层纤维中表达,且相较MyHC1、MyHC3、MyHC8为弱表达,而眶层纤维几乎没有表达。其在3组眼外肌的球层的表达差异没有统计学意义(P=0.147),见图2D-F和表1。
MyHC3 在外眶层的所有纤维中均有表达,在内眶层和球层的部分纤维中表达(图3A-C)。
MyHC3 在内眶层和球层的表达组间差异有统计学意义(均P<0.001)。在内眶层,2 周龄组(P<0.001)与4 周龄组(P=0.040)的表达水平较3月龄组高。2 周龄组和4 周龄组之间的表达水平差异无统计学意义(P=0.101)。在球层,同样为2周龄组与4 周龄组的表达水平均较3 月龄组高(均P<0.001),而2 周龄组与4 周龄组之间差异无统计学意义(P=0.054)。见表1。
MyHC8 在3 组兔眼外肌的所有肌纤维中均有表达,不随年龄的增长而改变(图3D-F)。
2.3 活化卫星细胞比例的变化
图4 显示眶层的活化卫星细胞数量多于球层。2周龄组外眶层、内眶层和球层的活化卫星细胞比例分别为2%(3%)、0%(1%)、0%(0%);4周龄组分别为4%(5%)、1%(9%)、0%(1%);3月龄组分别为20%(13%)13%(16%)、13%(5%)。内、外眶层和球层的活化卫星细胞比例组间差异均有统计学意义(P<0.001)。在校正显著性水平之前,均表现为2周龄组与4周龄组的数量低于3月龄组(均P<0.05)。只有4 周龄组和3 月龄组的内眶层的活化卫星细胞比例在用Bonferroni法校正显著性水平后,差异无统计学意义(P=0.065)。
3 讨论
斜视在人群中的患病率为3%~6%,是常见的眼科疾病。目前斜视的发病机制尚不明确,认为与融合机制、神经系统发育、屈光与调节及眼外肌等的异常有关。眼外肌作为特殊的骨骼肌,在形态上可以分为内、外眶层和球层[7];同时有MyHC构成复杂[8],终身存在活化卫星细胞[10]等特点。
MyHC是肌球蛋白的主要组成成分,其构成主要决定了肌纤维的收缩特性,对眼外肌完成精确的眼位调控具有重要意义[8]。卫星细胞是肌肉组织的干细胞,是眼外肌再生修复的主要细胞来源。与肢体骨骼肌不同,即使在正常生理状态下,眼外肌也存在活化卫星细胞[9]。二者能较好地体现眼外肌的功能特点和修复能力。既往对发育期个体眼外肌的变化特点尚不明确,限制了我们对斜视发病及治疗后变化的机制探索。
本研究发现处于发育期的2周龄至3月龄兔眼外肌存在组织学上的差异。随着年龄增长,眼外肌形态逐步发育完善、肌纤维增粗,内、外眶层和球层之间的界限愈发明显;未成熟亚型MyHC(MyHC3)表达减少,成熟亚型MyHC(MyHC1)表达增加;并伴有活化卫星细胞数量的持续增加。理清发育期个体的眼外肌组织学特点将为更深入地探究斜视的发病机制打下基础,也能为临床斜视手术干预时机的选择提供新的思路。
3.1 MyHC构成随发育变化的特点
本研究发现MyHC构成在发育期兔的眼外肌内、外眶层和球层存在变化:反映emb未成熟亚型的MyHC3在内眶层和球层的表达逐渐减少,反映fast-2X成熟亚型的MyHC1在外眶层的表达逐渐增加;而反映neo亚型的MyHC8 和slow-tonic成熟亚型的MyHC14则没有显著改变。这种变化可能是对眼外肌精细运动功能的适应性改变[8]。
以往研究比较幼年和成年等阶段大、小鼠,发现随着年龄的增加,未成熟亚型(MyHC3 和MyHC8)的表达减少[11],而成熟亚型(如fast、slow等亚型)的表达增加[12]。但这些研究均未考虑青春期这一特殊阶段。在年龄选择上,本研究着眼于幼年期和青春期的兔子,更符合临床上斜视手术的受众群体的年龄分布。同时,本研究根据前人经验[7],将眼外肌的眶层进一步分成内、外眶层,这种更精细的分层一方面细化了MyHC的发育特点,另一方面也为更精准地探究眼外肌运动的调控机制做了铺垫。
本研究中,MyHC8(neo亚型)表达稳定,并未随MyHC3(emb亚型)表达减少。这可能是同Zhou等[11]针对小鼠的研究类似,即neo亚型表达的减少要较emb亚型晚。既往基于大鼠的研究[13]发现,MyHC14(slow-tonic亚型)在出生前为高水平表达;出生2周后,表达水平下降,最终仅在少数纤维中分散表达。在本实验中,slow-tonic亚型也在球层纤维中分散表达,但不随发育改变,说明在兔眼外肌中,该亚型的发育性改变可能出现在出生后2周内或3个月之后。
MyHC的表达受复杂机制的调控,目前认为,可能和遗传因素[14]、神经支配[15]、机械刺激[12]、激素水平[16](如性激素、甲状腺激素)等因素相关。在未来研究中,同期探究其神经发育改变及激素水平变化将有利于增进对MyHC不同亚型表达的调控机制的理解。
3.2 活化卫星细胞数量随发育的变化特点
通常认为Pax7 是卫星细胞的特异性标记,活化的卫星细胞则特异性地表达MyoD[17]。目前少有研究探究眼外肌的活化卫星细胞的数量与年龄的关系,尤其与发育不同阶段的关系。本研究发现活化卫星细胞在发育期兔的眼外肌中随着年龄的增长而增多。
Formicola等[18]探究了成年和老年小鼠眼外肌的卫星细胞数量,发现老年小鼠的卫星细胞少于成年小鼠。本研究以幼年(2周龄)至性成熟期(3月龄)的新西兰大白兔眼外肌为研究对象,发现2周龄和4周龄兔眼外肌的活化卫星细胞数量没有差异,且数量极少(0~1%)。3月龄时,活化卫星细胞数量显著增加。3 月龄兔正处于性成熟期,对应于人类的青春期[19],该阶段是发育成熟的一个重要时期,提示该阶段个体的眼外肌组织修复能力可能更强。
此外,卫星细胞的数量受性激素水平[20]、甲状腺激素水平[21]、胰岛素样生长因子-2表达量[22]等的影响,仍有待进一步研究以上激素和生长因子是否参与了活化卫星细胞数量的改变。同时,3月龄之后,活化卫星细胞数量是仍旧保持高水平还是随着发育成熟而进一步发生改变,也需要进一步实验研究加以验证。
3.3 视觉输入在眼外肌发育过程中所起的作用
以往研究发现眼外肌的发育受视觉刺激的影响。一方面,眼睑张开时机是促进眼外肌发育的一个关键期[8,12,23-24]。不同类型的MyHC的表达量发生改变:成熟亚型MyHC(如fast ⅡA/ⅡX亚型)的表达只有在眼睑张开后才出现[12],未成熟亚型(如emb亚型)则在这个阶段开始表达减少[23];同时伴随着α-辅肌动蛋白3、伴肌动蛋白的出现和表达增加[23]。此外,nm MyHC ⅡB的表达位置由肌膜下向球层的肌纤维过渡[24]。另一方面,眼外肌的正常发育也需要正常的视觉刺激。暗室饲养会引起涉及能量代谢、蛋白质合成、脂质代谢、肌肉相关等基因的改变[25]。同时暗室饲养导致大鼠眼外肌的fast/slow亚型MyHC的表达下降[26];单眼视觉信息剥夺导致眼外肌肌纤维构成的改变[27],滑车神经变细[28]。此外,MyHC的表达也受前庭-眼反射的调控[27]。可见异常的视觉经验可能会直接扰乱眼外肌的正常发育,从而诱发斜视的发生。而斜视所致的异常视觉经验对眼外肌发育的影响也值得进一步探讨。
新西兰大白兔一般在2 周龄左右张开眼睑[29],本研究结果表明新西兰大白兔在睁眼后至成年前的快速发育阶段,眼外肌的各生物学特性也存在发育性改变,提示幼年至青春期可能是眼外肌发育成熟的一个重要时期。当然,本研究尚存在诸多不足。首先,本研究未纳入眼睑张开前及成年后兔眼外肌的特点,在未来实验中纳入发育前期(眼睑张开前,如刚出生或1周龄)和成年兔(6月龄~1岁龄)及其他年龄段兔为研究对象,将进一步明确各发育阶段眼外肌的特点。其次,本研究是建立在动物实验上,因此所得结论可能不能完全照搬至人类;但这为人类的相关研究打下了基础。
综上,发育期兔的眼外肌也存在发育性改变,无论是肌肉的组织形态还是MyHC的构成及活化卫星细胞数量均发生了显著变化。本研究结果为斜视发病机制的探究提供了一定思路。同时,在未来实验中,可以兔眼外肌为模型,探究不同发育阶段的眼外肌的手术效果,为临床斜视手术干预时机提供更多参考。
利益冲突申明本研究无任何利益冲突
作者贡献声明楼江涛:收集数据,参与选题、设计及资料的分析和解释;撰写论文;对编辑部的修改意见进行修改。彭一一:收集数据,参与选题、设计。王春晓:收集数据。许梅萍、余新平:选题、 设计,修改论文中关键性结果、结论,对编辑部的修改意见进行核修
猜你喜欢
动物营养学报(2022年11期)2022-12-08
特产研究(2021年6期)2021-12-08
科学生活(2019年7期)2020-01-01
河南农业科学(2019年9期)2019-09-24
养殖与饲料(2019年10期)2019-02-25
山东畜牧兽医(2018年3期)2018-04-26
农村百事通(2016年6期)2016-05-14
中国卫生标准管理(2015年8期)2016-01-15
听力学及言语疾病杂志(2015年5期)2015-12-24
中国当代医药(2015年29期)2015-03-01
