
饲粮中添加不同饼粕源发酵饲料对断奶仔猪生长性能、营养物质表观消化率、血清生化指标和肠道菌群的影响
2024-03-05 12:13李佳橙张文喆南珊珊薛瑞鑫王海亮冯家骐聂存喜
动物营养学报 2024年2期
李佳橙 张文喆 南珊珊 薛瑞鑫 王海亮 匡 宇 冯家骐 聂存喜*
(1.石河子大学动物科技学院,石河子 832000;2.新疆农垦科学院,石河子 832000)
近年来,随着畜牧业的不断发展,饲料资源越发短缺。豆粕作为一种优质的蛋白质饲料被广泛的用于全世界畜牧业生产中[1],尽管我国大豆产量逐年上升,2020年产量已达1 960.18万t,但国产大豆无法满足饲料用豆粕的缺口,2020年进口大豆10 031.45万t,其中进口大豆的80%被用于加工成为畜禽饲料(数据来自国家统计局)。我国大豆产量不足,且近年来豆粕价格波动严重[2],制约着我国畜牧业的发展。为解决供需矛盾,降低养殖成本,迫切需要更多优质、廉价的蛋白质饲料替代豆粕,以解决养殖业蛋白质饲料来源问题。饼粕是油料榨油后剩余的固体部分,蛋白质含量一般在30%以上,是良好的豆粕替代品[3]。但饼粕类饲料中含有大量的抗营养因子,如游离棉酚、酚基葡糖苷、2-羟基牛蒡等[3-4]。因此,过量的饼粕类饲料添加到动物饲粮中会对动物的生长性能和营养物质表观消化率产生负面影响[5],限制了饼粕类饲料的广泛应用。
益生菌发酵饲料能够提高断奶仔猪的生长性能,有效调节断奶仔猪的肠道微生物平衡[6],在微生物发酵后饼粕类饲料中的植物蛋白质被微生物分解为易于吸收的小分子蛋白肽,并且抗营养因子水平会显著降低[7-8],益生菌及其代谢产物还有利于抑制肠道中的致病菌并提高营养物质表观消化率[9]。研究表明,在饲粮中用发酵棉籽粕部分替代部分豆粕后,对促进肉鸡的生长性能、减缓脂肪沉积有积极的作用[10-11],同时可以降低断奶仔猪的腹泻率、改善肠道菌群[12]。王荷香等[13]研究发现,在饲粮中添加发酵菜籽粕对育肥猪生长性能没有负面影响。由此可知,饼粕类饲料在经过微生物发酵后可以作为一种良好的豆粕替代品。
仔猪在早期断奶后,肠道没有发育完善,同时其营养来源由易于被吸收的母乳变为不易被消化吸收的固态饲料,容易因肠道应激导致出现腹泻、食欲减退等现象[14]。饲喂抗生素可以有效解决这一问题,但自“禁抗令”颁布以来,寻找抗生素的替代产品显得尤为关键,而发酵饲料具有完善肠道功能、优化肠道菌群结构和降低腹泻率的作用[15-16],可以有效地减少仔猪断奶应激。例如,Gu等[12]研究表明,在饲粮中用6%的发酵棉籽粕替代豆粕可以使断奶仔猪腹泻率显著降低49.37%。单一饼粕类饲料中的氨基酸相比于豆粕,无法满足仔猪的氨基酸需要[17],不同来源的蛋白质饲料可以平衡氨基酸组成,且存在着的活性因子可以促进仔猪肠道的发育,减少应激对仔猪带来的伤害,提高生长性能,减低死亡率,帮助仔猪安全度过断奶阶段。基于此,本试验将不同饼粕源发酵饲料及其复合物添加到饲粮中,探究其对断奶仔猪生长性能、腹泻率、血清生化指标和肠道菌群的影响。
1 材料与方法
1.1 试验材料
棉籽粕、葵花籽粕、红花籽粕、麸皮和玉米粉购买自石河子市农贸市场。热带假丝酵母ZD-3和酿酒酵母CICC1001由石河子大学动物科技学院动物营养与饲料科学实验室保存。根据实验室前期优化[18-19],发酵底物为饼粕∶玉米∶麸皮=90∶5∶5,底物含水量为40%。棉籽粕和葵花籽粕接种热带假丝酵母ZD-3,红花籽粕接种酿酒酵母CICC1001,每100 g底物接种8 mL菌液(5.0×108CFU/g)和0.2 g酸性蛋白酶,混合均匀后放入30 ℃培养箱培养48 h。按照发酵后3种饼粕的氨基酸组成和仔猪的氨基酸需要制备复合发酵饼粕(发酵棉籽粕∶发酵葵花籽粕∶发酵红花籽粕=3∶1∶1)。
1.2 试验设计和饲养管理
选取120头28日龄“杜洛克×长白×大约克夏”三元杂交断奶仔猪,体重(8.42±0.76) kg。随机分成5组,每组6个重复,每个重复4头猪。参照《猪营养需要量》(GB/T 39235—2020)设计饲料配方,试验饲粮组成及营养水平见表1。对照组(C组)饲喂基础饲粮;4个试验组(M组、K组、H组和F组)分别用6%的不同饼粕源发酵饲料替代基础饲粮中6%的豆粕,其中,M组为发酵棉籽粕,K组为发酵葵花籽粕,H组为发酵红花籽粕,F组为复合发酵饼粕(发酵棉籽粕、发酵葵花籽粕和发酵红花籽粕按照3∶1∶1混合而成)。预试期5 d,正试期21 d。试验在石河子市科盛种猪场进行,试验猪自由饮水和采食,防疫消毒流程均按照试验猪场管理规定进行。
ADG(g/d)=(末重-初重)/试验天数;ADFI(g/d)=总采食量/(试验猪头数×试验天数);F/G=试验期总耗料量/总增重;腹泻率(%)=100×腹泻仔猪头数/(每组猪头数×试验天数)。

表1 试验饲粮组成及营养水平(风干基础)
1.3 样品采集和指标测定
1.3.1 饲粮营养水平的测定
按照《饲料分析及饲料质量检测技术》中的方法对饲粮中粗蛋白质、钙、总磷等营养物质含量进行测定。
1.3.2 生长性能和腹泻率的测定
在试验开始和结束时对空腹过夜的仔猪进行称重,每天记录各栏的仔猪采食量,计算平均日增重(ADG)、平均日采食量(ADFI)及料重比(F/G)。每天10:00和18:00观察记录肛门处被粪便污染的仔猪头数和粪便形态,计算腹泻率。计算公式如下:
1.3.3 营养物质表观消化率的测定
在试验结束前3 d,每天18:00从每个重复中取约100 g无污染粪便,加入少量浓硫酸(H2SO4)固氮,存放于-20 ℃冰箱中。检测样品时,将每个重复3 d收集的粪样均匀混合,在65 ℃干燥箱中烘48~72 h去除样品中的游离水,用粉碎机将样品粉碎过40目筛,4 ℃密封保存待测。以样品中的盐酸不溶灰分为内指示剂,计算干物质、粗蛋白质、粗脂肪、有机物、中性洗涤纤维表观消化率。干物质、粗蛋白质、粗脂肪、有机物、中性洗涤纤维按照《饲料分析及饲料质量检测技术》中的方法进行测定,盐酸不溶灰分含量按照GB/T 23742—2009进行测定。营养物质表观消化率计算公式如下:
某营养物质表观消化率(%)=100-[(饲粮中盐酸不溶灰分含量/粪中盐酸不溶灰分含量)×(粪中该营养物质含量/饲粮中该营养物质含量)×100]。
1.3.4 血清生化指标的测定
在试验结束后,每个重复随机选取1头空腹过夜的仔猪,用非抗凝管进行前腔静脉采血,静置于4 ℃冰箱1 h后,4 000 r/min离心20 min,分离血清,-20 ℃保存待测。采用试剂盒测定血清总蛋白(TP)、白蛋白(ALB)、球蛋白(GLB)、尿素氮(UN)、总胆固醇(TC)、甘油三酯(TG)、高密度脂蛋白(HDL)、低密度脂蛋白(LDL)含量,所有试剂盒均购自南京建成生物工程研究所。
1.3.5 粪便挥发性脂肪酸含量的测定
试验结束时,从每个重复中随机选择1头猪,收集其新鲜粪便,经液氮速冻后转移到-80 ℃冰箱保存,待测。粪便挥发性脂肪酸含量测定参照Nan等[20]的方法。将样品放置4 ℃冰箱解冻,准确称量样品0.5 g,将样本用超纯水稀释5倍,加入0.2 mL偏磷酸,5 000 r/min离心10 min。取上清液过0.45 μm的水系膜后采用美国安捷伦气相色谱仪(Agilent-7890B)测定样品中挥发性脂肪酸(乙酸、丙酸、丁酸)含量,色谱柱为DB-WAX(30 m×0.25 mm×0.50 μm)。
1.3.6 粪便微生物16S rRNA测序
将1.3.5中每个重复收集的新鲜粪便按照QIAamp®DNA Stool Mini Kit试剂盒说明书进行基因组DNA提取,基因组DNA完整性利用1%琼脂糖凝胶电泳检测,-20 ℃保存。16S rRNA测序工作由上海美吉生物医药科技有限公司代理完成。具体流程为:设计合成引物接头→PCR扩增与产物纯化→PCR产物定量与均一化→构建PE文库→Illumina测序。利用Illumina MiSeq测序平台对样品进行质控和过滤后,操作分类单元(OTU)聚类方式为按照97%相似性对非重复序列(不含单序列)进行,同时去除嵌合体,得到OTU的代表序列。随后按照上海美吉生物医药科技有限公司数据分析云平台(https://report.majorbio.com/)的说明进行Alpha多样性、Beta多样性和相关性分析。
1.4 统计分析
试验数据经Excel 2013初步整理、计算、统计后,使用SPSS 20.0软件进行单因素方差分析(one-way ANOVA),并用Duncan氏法进行多重比较检验分析。结果以“平均值±标准差”表示,P<0.05表示差异显著。
2 结果与分析
2.1 饲粮中添加不同饼粕源发酵饲料对断奶仔猪生长性能及腹泻率的影响
由表2可知,各组之间初重和末重均无显著差异(P>0.05)。与C组相比,F组的腹泻率显著降低了26.65%(P<0.05);M组、K组、H组的腹泻率有降低趋势,分别降低了17.44%、8.91%、15.70%(P>0.05)。与C组相比,M组、K组、H组、F组的ADG有升高趋势,分别增加了12.59%、21.00%、18.56%、20.69%(P>0.05)。与C组相比,M组、K组、H组、F组的F/G有降低趋势,分别降低了19.08%、11.84%、14.47%、17.11%(P>0.05)。

表2 饲粮中添加不同饼粕源发酵饲料对断奶仔猪生长性能及腹泻率的影响
2.2 饲粮中添加不同饼粕源发酵饲料对断奶仔猪营养物质表观消化率的影响
由表3可知,与C组相比,F组的干物质、粗蛋白质、有机物、中性洗涤纤维表观消化率分别显著升高了12.24%、17.82%、0.06%、14.11%(P<0.05),H组的粗蛋白质表观消化率显著升高了17.78%(P<0.05)。
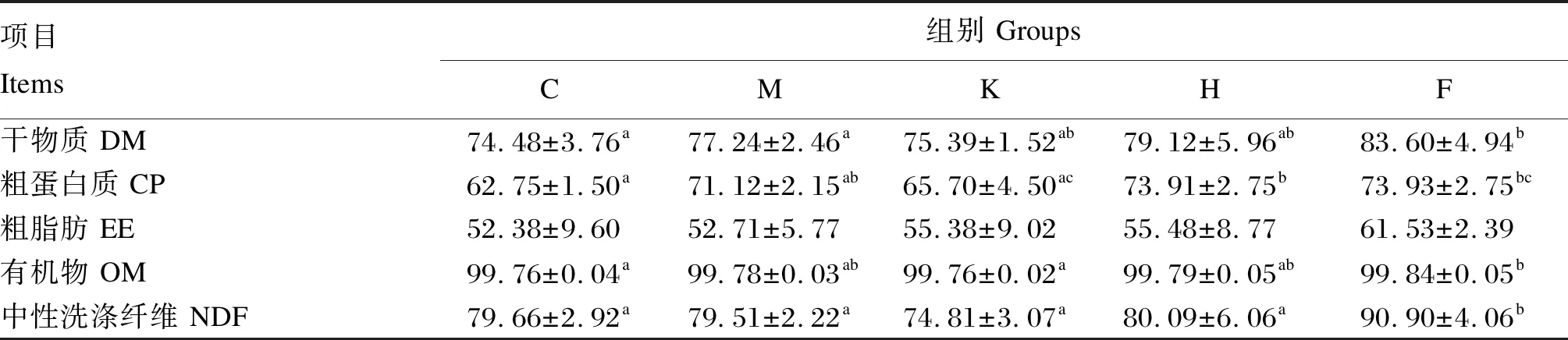
表3 饲粮中添加不同饼粕源发酵饲料对断奶仔猪营养物质表观消化率的影响
2.3 饲粮中添加不同饼粕源发酵饲料对断奶仔猪血清生化指标的影响
由表4所示,各组之间血清TP、ALB、GLB、UN、TC、TG、HDL和LD含量均无显著差异(P>0.05)。与C组相比,F组的血清TP、GLB含量分别升高了3.48%和8.81%(P>0.05),血清TC和TG含量分别降低了6.40%和10.40%(P>0.05)。

表4 饲粮中添加不同饼粕源发酵饲料对断奶仔猪血清生化指标的影响
2.4 饲粮中添加不同饼粕源发酵饲料对断奶仔猪肠道菌群的影响
2.4.1 α多样性和β多样性分析
α多样性指数包括Shannon指数、Simpson指数、Ace指数和Chao指数。Ace指数和Chao指数反映出样本中群落物种的种类,即丰富度。由表5可知,与C组相比,M组、K组、H组、F组的Ace指数和Chao指数显著升高(P<0.05),说明在饲粮中添加不同饼粕源发酵饲料后提高了肠道菌群的丰富度。Shannon指数和Simpson指数受到样本均匀度的影响,即物种的多样性,Shannon指数与样本物种多样性呈正相关,而Simpson指数与样本物种多样性呈负相关。与C组相比,M组、K组、H组和F组的Shannon指数显著升高(P<0.05),Simpson指数显著降低(P<0.05),说明各试验组的丰富度和均匀度均高于对照组。

表5 饲粮中添加不同饼粕源发酵饲料对断奶仔猪肠道菌群α多样性指数的影响
β多样性分析通过对组间不同微生物群落多样性进行比较分析,体现出不同组间微生物群落组成的相似性或差异性。由图1可知,饲粮中添加不同饼粕源发酵饲料后,M组、K组、H组、F组显示出了与C组不同的聚类,说明不同饼粕源发酵饲料改变了肠道菌群群落的组成。

图1 主坐标分析图
2.4.2 菌群结构和组成差异分析
Venn图体现多组间所共有和独有的OTU数目,可以直观的体现组间物种组成情况。由图2可知,C组、M组、K组、H组和F组的粪便中OTU数目,分别为704、939、952、923、826个,其中共有的OTU数目为480个,C组、M组、K组、H组和F组独有的OTU数目分别为29、34、50、33、23个。

图2 韦恩图
通过图3和表6可知,在门水平上,5组均以厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidota)为主,其中C组、M组、K组、H组和F组的Firmicutes和Bacteroidota相对丰度分别为92.09%、85.71%、84.70%、91.01%、84.57%和1.97%、11.79%、11.31%、4.70%、8.36%。与C组相比,M组、K组、H组和F组的Bacteroidota相对丰度分别提高了9.82%(P<0.05)、9.34%(P<0.05)、2.73%(P>0.05)、6.39%(P<0.05)。
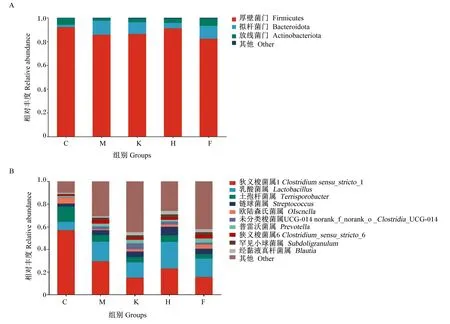
A为门水平相对丰度;B为属水平相对丰度。
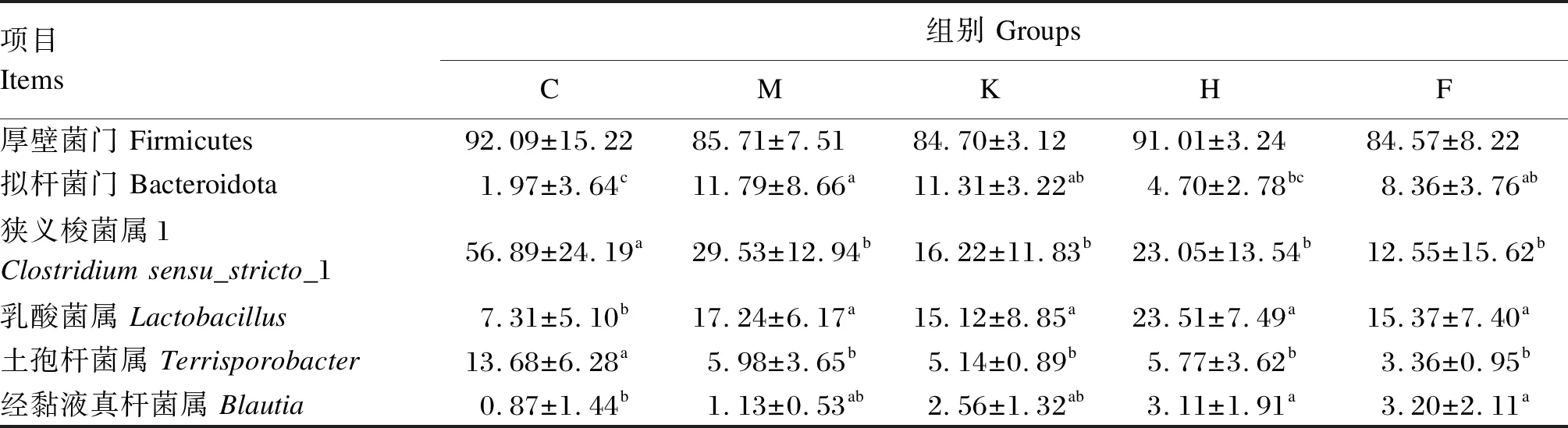
表6 门、属水平相对丰度比较
在属水平上,对相对丰度前10的菌属进行分析,C组各样本微生物主要分布在狭义梭菌属1(Clostridiumsensu_stricto_1)、乳酸菌属(Lactobacillus)和土孢杆菌属(Terrisporobacter),占总比的77.88%,其他菌属占比较低,而M组、K组、H组和F组各菌属分布均匀度提高,这与α多样性结果相互印证。与C组相比,M组、K组、H组和F组的Clostridiumsensu_stricto_1相对丰度显著降低(P<0.05),Lactobacillus相对丰度分别显著提高了9.93%、7.81%、16.20%、8.06%(P<0.05),Terrisporobacter的相对丰度分别显著降低了7.70%、8.54%、7.91%、10.02%(P<0.05);M组和K组的经黏液真杆菌属(Blautia)相对丰度提高了0.26%和1.69%(P>0.05),H组和F组显著提高了2.24%和2.33%(P<0.05)。
线性判别分析效应大小(LEfSe)是一种用于发现高维生物标识和揭示基因组特征的软件,通过采用线性判别分析(LDA)可以估算每个组分(物种)相对丰度对差异效果。由图4可知,通过LEfSe分析(LDA阈值为3)发现,未分类消化链球菌科(unclassified_f_Peptostreptococcaceae)和棒状杆菌属(Corynebacterium)在C组中显著富集;norank_f_Muribaculaceae、普雷沃氏菌科NK3B31群(Prevotellaceae_NK3B31_group)、毛螺菌科NK4A136群(Lachnospiraceae_NK4A136_group)等7个菌属在M组中显著富集;未分类梭菌属UCG-014(norank_f_norank_o_Clostridia_UCG-014)、琼脂杆菌属(Agathobacter)等28个菌属在K组中显著富集;Blautia、普拉梭菌属(Faecalibacterium)等5个菌属在H组中显著富集;考拉杆菌属(Phascolarctobacterium)、厚壁菌门未分类菌属(unclassified_p_Firmicutes)、丁酸球菌属(Butyricicoccus)等12个菌属在F组中显著富集。

图4 微生物LEfSe分析及LDA得分图
2.5 饲粮中添加不同饼粕源发酵饲料对断奶仔猪粪便中挥发性脂肪酸含量的影响
由表7可知,各组之间粪便中乙酸、丙酸含量均无显著差异(P>0.05)。F组的粪便中丁酸含量较C组、M组、H组分别显著提升了42.98%、36.40%、37.18%(P<0.05)。

表7 饲粮中添加不同饼粕源发酵饲料对断奶仔猪粪便中挥发性脂肪酸含量的影响
2.6 肠道菌群与相关指标的相关性分析
相关性热图(heatmap)通过相关性数值可视化展示样本中不同的物种与试验数据之间的关系,评估了肠道菌群与生长性能、腹泻率、挥发性脂肪酸含量和营养物质表观消化率之间的相关性。由图5可知,乙酸含量与未分类毛螺菌科(unclassified_f_Lachnospiraceae)、普氏菌属(Prevotella)、norank_f_Muribaculaceae、文肯菌科RC9肠道群(Rikenellaceae_RC9_gut_group)相对丰度呈显著负相关(P<0.05),相关系数为-0.38~-0.47;腹泻率与Prevotella、norank_f_Muribaculaceae、Rikenellaceae_RC9_gut_group、Prevotellaceae_NK3B31_group、norank_f_Eubacterium_coprostanoligenes_group、Faecalibacterium相对丰度呈显著负相关(P<0.05),相关系数为-0.38~-0.53;粗蛋白质表观消化率与Clostridiumsensu_stricto_1、Terrisporobacter相对丰度呈显著负相关(P<0.05),相关系数为-0.39~-0.45,与Lactobacillus、罕见小球菌属(Subdoligranulum)、未分类梭菌目UCG-014(norank_f_norank_o_Clostridia_UCG-014)、未分类丁酸球菌科(norank_f_Butyricicoccaceae)、Agathobacter、Blautia等10个菌属相对丰度呈显著正相关(P<0.05),相关系数为0.38~0.68;丁酸含量与Rikenellaceae_RC9_gut_group、norank_f_norank_o_Clostridia_UCG-014、Agathobacter、Blautia和UCG-005相对丰度呈显著正相关(P<0.05),相关系数为0.38~0.44;ADG与norank_f_norank_o_Clostridia_UCG-014、未分类丁酸球菌科(norank_f_Butyricicoccaceae)、UCG-005、norank_f_norank_o_RF39相对丰度呈显著正相关(P<0.05),相关系数为0.38~0.56。
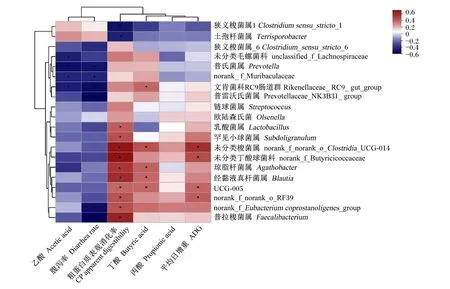
*表示显著相关(P<0.05)。* mean significant correlation (P<0.05).
3 讨 论
3.1 饲粮中添加不同饼粕源发酵饲料对断奶仔猪生长性能及腹泻率的影响
前人研究表明,饼粕类饲料在经过益生菌发酵后,其中的非淀粉多糖等损害动物生长性能的抗营养因子水平显著降低[21],酸溶蛋白、粗蛋白质含量显著提高,中性洗涤纤维、酸性洗涤纤维含量显著降低,同时会改变底物中有机酸、多肽和糖类的含量[18]。发酵饲料中的益生菌在进入机体后,能够有效地提高营养物质表观消化率[22],从而提高畜禽的生长性能。有报道指出,与未发酵棉籽粕相比,发酵棉籽粕可以提高肉鸡的生长性能和屠宰性能[23];与未发酵的菜籽粕相比,发酵菜籽粕可以显著提高肉仔鸡的生长性能[24]。此外,Zhong等[25]将断奶仔猪饲粮中5%蛋白质来源替换为大豆、酪蛋白和玉米酒精糟,结果表明不同来源的蛋白质对动物机体的影响不同。另有研究表明,益生菌发酵饲料之所以能够改善生产性能是由于在发酵过程中益生菌提高了饲料利用效率,而不是增加了动物的采食量[26]。在本研究中,添加不同饼粕源发酵饲料后,断奶仔猪的生长性能有增加的趋势,说明不同饼粕源发酵饲料可以在一定程度上改善仔猪的生长性能。
断奶仔猪的胃肠功能没有发育健全,断奶后易产生应激性腹泻,而腹泻是反映断奶仔猪肠道状态最直观的表现[27]。有研究表明,发酵棉籽粕[12]、发酵中草药渣[28]和发酵大豆[29]均可以显著降低断奶仔猪的腹泻率,由此可见发酵饲料可以有效地缓解断奶应激导致的腹泻,这与本试验的研究结果一致。益生菌及其代谢产物有利于肠道有益菌的繁殖,同时,益生菌可以影响肠道上皮细胞的更新,增加紧密连接蛋白和黏蛋白的产生,调节肠道抗菌肽的释放,与致病菌争夺营养和生存空间,并与宿主和肠道共生菌群相互作用,恢复肠道屏障[30],从而达到减缓断奶仔猪腹泻的目的。前期研究表明,热带假丝酵母和酿酒酵母可以很好地发挥益生菌的作用[18-19]。因此,在本试验中不同饼粕源发酵饲料组的腹泻率较对照组均有所改善,但只有F组的腹泻率降低达到了显著水平。这可能与不同饼粕源发酵饲料中富含膳食纤维有关。杨梦琪等[31]研究表明,发酵棉籽粕的膳食纤维可以提高黄羽肉鸡的绒毛长度和隐窝深度,同时Chen等[32]研究发现,不同来源的膳食纤维对断奶仔猪的肠道屏障功能影响不同。
3.2 饲粮中添加不同饼粕源发酵饲料对断奶仔猪营养物质表观消化率的影响
营养物质表观消化率是反映机体吸收的重要指标。大量的研究证明,发酵饲料较未发酵饲料可以促进机体的营养物质表观消化率[29,33]。但在本试验中,M组、K组的营养物质表观消化率较C组没有显著差异变化,且中性洗涤纤维表观消化率有降低趋势。饼粕类饲料中粗纤维含量较高,即使经过益生菌发酵依然会影响营养物质表观消化率。邓铭等[34]发现在饲粮中添加3%、6%、9%的发酵构树饲料显著降低了粗蛋白质表观消化率,其原因可能为这些饼粕类饲料中残留的抗营养物质与益生菌对营养物质表观消化率的增益作用相抵消导致的。H组的粗蛋白质表观消化率显著提高,这可能是因为红花籽粕中含有抗氧化物质[35],抑制了肠道中的炎症反应,从而提升了营养物质表观消化率。不同来源的蛋白质对于动物消化酶和胆汁分泌影响不同[36],赵金标等[37]研究发现,在生长猪饲粮中由于纤维组成和理化性质的差异,导致其营养物质表观全肠道消化率和能量利用效率存在很大差异,与本试验结果相同。
3.3 饲粮中添加不同饼粕源发酵饲料对断奶仔猪血清生化指标的影响
血清生化指标可以反映出机体对营养的代谢情况。血清TP含量与蛋白质在机体内消化、吸收、代谢有直接的关系[38]。TP由ALB和GLB组成,ALB由肝脏合成,是运输合成原料和代谢废物的载体;GLB与机体免疫力相关[39]。UN是蛋白质分解的最终产物,代表着蛋白质代谢与氨基酸的平衡,其含量越低,说明氨基酸合成能力越高[40]。TP、ALB、GLB和UN共同反映着机体蛋白质的代谢。在本试验中,F组的血清TP含量有升高趋势,血清UN含量有降低趋势,这与营养物质表观消化率结果相互印证,说明F组可以提高断奶仔猪对饲粮蛋白质的吸收和代谢,具有促生长的作用。血清TC、TG、HDL和LDL是机体内脂质代谢的标志物。其中TC的主要作用为形成胆汁、构成细胞膜和合成激素参与到各种物质代谢中[41];TG反映着脂肪组织合成情况[42];HDL和LDL是胆固醇在血液中的载体,HDL可以将血清中的胆固醇运送到肝脏中合成其他物质,俗称“血液清道夫”,LDL主要是把肝脏中的胆固醇运送到全身各处[43]。Nan等[20]在肉鸡饲粮中添加6%发酵葡萄籽粕发现,处理前后血清TC和TG含量虽没有显著差异,但有下降趋势;王荷香等[13]研究发现,在断奶仔猪饲粮中添加发酵菜籽粕对于断奶仔猪的血清TC、TG含量没有显著影响,但有下降趋势,这与本试验结果相同,其中以F组下降趋势最为明显。也有研究表明在畜禽饲粮中添加益生菌发酵饲料可以显著影响机体的脂质代谢,如Niu等[44]研究发现,发酵棉籽粕可以影响黄羽肉鸡脂质代谢,进而减少腹部脂肪沉积,但在本试验结果中并没有达到显著水平,其原因为受到发酵菌种、发酵底物的影响。综上所述,饲粮中添加不同饼粕源发酵饲料对断奶仔猪血清生化指标无不良影响,并可以在一定程度上提高蛋白质和脂质的利用率。
3.4 饲粮中添加不同饼粕源发酵饲料对断奶仔猪肠道菌群的影响
在肠道中寄生着各种各样的细菌,这些细菌分为有益菌、中间菌和有害菌,当肠道环境健康时这3类细菌处于动态平衡状态,断奶会破坏仔猪肠道菌群平衡[45]。在饲粮中添加发酵饲料可以改善畜禽的肠道菌群,从而改善其健康状态和生长性能[46]。大量研究表明,在动物饲粮中添加发酵饲料可以提高肠道菌群的多样性[12,44],比如,Huang等[27]研究了发酵饲料对仔猪粪便菌群的影响,与本试验结果相同,发酵饲料组的α多样性和β多样性均得到了改善。在仔猪肠道中Firmicutes和变形菌门(Proteobacteria)是最优势的菌门[47],Firmicutes可以通过合成β-半乳糖苷酶参与乳糖消化[48],而Proteobacteria可以编码糖苷水解酶和多糖裂解酶参与到多糖的降解中[49]。在本研究中,通过饲喂不同饼粕源发酵饲料增加了Blautia和Terrisporobacter的丰富度,而相关性分析显示Blautia和Terrisporobacter与丁酸代谢呈正相关[50],这也是粪便中丁酸含量的增加的原因。有研究表明,Lactobacillus作为一种益生菌,可以降低肠道的pH,进而抑制大肠杆菌、葡萄球菌等有害菌的增殖[51],而梭状芽胞杆菌、Terrisporobacter则常被认为是一种潜在的病原体[52]。有研究发现,蒙花苷可以逆转葡聚糖硫酸钠(DSS)损伤的肠道菌群,其中就包括norank_f_norank_o_Clostridia_UCG-014[53],而在本试验中,norank_f_norank_o_Clostridia_UCG-014相对丰度与ADG呈正相关,且各试验组其相对丰度均有增加。而Faecalibacterium和Prevotella被认为可以抵抗肠道炎症而减少腹泻[54-55]。结合本试验结果,在饲粮中添加不同饼粕源发酵饲料可以改善断奶仔猪的肠道菌群,进而调控断奶仔猪的健康状态和生长性能,其主要表现在降低了断奶仔猪的腹泻率。
3.5 饲粮中添加不同饼粕源发酵饲料对断奶仔猪粪便中挥发性脂肪酸含量的影响
挥发性脂肪酸是肠道微生物的代谢产物,同时也是蛋白质等在肠道的最终产物[56]。许多研究表明,单胃动物饲喂微生物发酵的饲料后,由于其肠道微生物的组成的改变,挥发性脂肪酸含量也会随之改变[23,44]。挥发性脂肪酸作为桥梁将肠道菌群和宿主之间紧密的联系在了一起[57]。例如,Zhao等[58]研究发现,饲粮纤维能增加产丁酸菌的丰富度从而改善断奶仔猪的生长性能。丁酸具有提高生长性能、降低腹泻率的作用;同时也是肠道细胞的代谢能量来源,具有维持宿主黏膜屏障完整性的作用[59],Liu等[60]研究表明,丁酸钠具有通过改变肠道菌群结构进而改善断奶仔猪生长性能和增强肠道黏膜屏障功能的功能;Chen等[61]研究发现,添加丁酸钠可以显著降低断奶仔猪的腹泻率。在本试验中,F组中复合饼粕源发酵饲料具有多种来源的膳食纤维,刺激了肠道蠕动,提高了肠道中的发酵效率,促进了丁酸的产生及肠道菌群的增殖,进而改善断奶仔猪的生长性能和肠道健康。
4 结 论
饲粮中添加6%不同饼粕源发酵饲料可以优化断奶仔猪肠道菌群的结构,并在一定程度上改善生长性能、降低腹泻率、提高营养物质表观消化率,其中添加复合发酵饼粕的效果最好。
猜你喜欢
湖南饲料(2022年6期)2022-12-20
当代水产(2021年2期)2021-03-29
湖南饲料(2019年5期)2019-10-15
广东饲料(2016年4期)2016-12-01
广东饲料(2016年1期)2016-12-01
新农业(2016年13期)2016-08-16
特产研究(2016年3期)2016-04-12
中国粮油学报(2016年1期)2016-02-06
动物营养学报(2015年10期)2015-12-01
食品科学(2013年17期)2013-03-11
