
骨髓基质细胞及在神经系统疾病治疗研究中的应用前景
2012-11-09 05:13谢三平姚长江长江大学荆州临床医学院荆州市中心医院神经内科湖北荆州434020
长江大学学报(自科版) 2012年6期
谢三平,姚长江 (长江大学荆州临床医学院 荆州市中心医院神经内科, 湖北 荆州 434020)
骨髓基质细胞及在神经系统疾病治疗研究中的应用前景
谢三平,姚长江 (长江大学荆州临床医学院 荆州市中心医院神经内科, 湖北 荆州 434020)
中枢神经系统损伤后的再生仍然是神经科学领域的难题,因此很多研究者开始探索干细胞移植治疗神经系统疾病的可行性,而骨髓基质细胞因自身优势,倍受研究者亲睐,本文重点就骨髓基质细胞及其在神经系统疾病中的应用研究进展进行综述。
骨髓基质细胞; 干细胞移植; 神经系统疾病
由于成熟的神经元是不可再生细胞,一旦受损后周围神经组织对其修复能力将极其有限,故其相关功能将永久性缺省或缺失,所以中枢神经系统(central nervous system,CNS) 损伤性或退行性病变造成的神经功能障碍将会长期影响患者的生存及生活质量。如何治疗修复受损的神经组织,恢复患者的神经功能一直是神经科学领域广为关注和困惑的问题。自1987年Gensburger[1]从胚胎鼠中培养出神经干细胞后,揭开了神经干细胞移植治疗神经系统疾病的序幕,从而为神经系统疾病的治疗研究开辟了一条崭新的途径。目前,神经干细胞移植治疗神经系统疾病的探索已成为神经科学领域研究的热点。但由于作为细胞移植治疗的神经干细胞都是来源于胚胎或胎儿的中枢神经系统,存在可用细胞数量有限、供体材料不易获得、伦理道德等问题限制了其在临床研究中的应用。来源于骨髓的间充质干细胞不仅无此限制,而且具有间充质-神经组织转化的可塑性,因此备受神经学者的青睐。本文重点就骨髓基质干细胞及其在神经系统疾病中的应用研究进展进行综述。
1 骨髓基质细胞的生物学特性及其分离方法
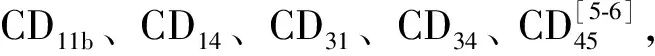
2 骨髓基质细胞的神经细胞分化潜能的体外研究
BMSCs分化为神经细胞的体外研究主要集中于化学试剂或是同神经组织细胞共同培养诱导。化学试剂诱导有抗氧化剂和神经生长因子诱导等,如全反式维甲酸(all-trans-retinoic acid,RA)、脑源性神经营养因子(Brain-Derived Neurotrophic Factor,BDNF)等。Sanchez-Ramos等[10]在研究BMSCs的神经细胞分化潜能时发现白细胞抑制因子(leucocyte inhibitory factor,LIF)可促使BMSCs向成纤维细胞分化,但不能诱导其分化为神经细胞,而BMSCs经RA和BDNF在体外共同诱导后发生了形态学的转化,由大扁平细胞转化成形态较小的纺锤形细胞,并形似神经细胞有长突起延伸于两端,经免疫荧光检测也证实有少量细胞表达神经细胞特异性表面标志nestin、NeuN和GFAP蛋白,细胞裂解物经Western blots检测也发现有nestin、NeuN和GFAP蛋白的表达。这些现象说明BMSCs有可能经体外诱导可分化为具有神经细胞特异性表面标志的神经细胞。将β-半乳糖苷酶基因标记的BMSCs同胚鼠中脑神经细胞在含RA和BDNF的神经组织细胞培养液中共同培养2周后免疫组织化学检测大约有2%~5%的β-半乳糖苷酶阳性细胞表达NeuN[10],明显高于单独培养出的BMSCs。同样绿色荧光标记的BMSCs同胚鼠中脑神经细胞在含RA和BDNF的神经组织细胞培养液中共同培养1周后免疫组织化学检测示骨髓基质来源的细胞中NeuN+细胞为2%~4%,GFAP+细胞为5%~8%,NeuN+细胞呈卵圆形或纺锤形,有双极突起,形似未成熟的神经元样细胞,位于微管缔合蛋白-2(microtubule-associated protein-2,MAP-2)阳性的胚鼠中脑神经细胞周围,但即使明确表达神经细胞标志的BMSCs也未见MAP2表达;大部分GFAP+细胞呈卵圆形或纺锤形,但其突起较NeuN+细胞的短,少部分GFAP+细胞形似神经胶质细胞呈不规则形[10]。BMSCs同胚鼠中脑神经细胞共同培养后NeuN+细胞和GFAP+细胞表达率均比单纯化学诱导增加1倍以上,说明BMSCs在向神经样细胞定向分化过程中,除细胞因子和营养因子的作用外,细胞间的相互作用也是不容忽视的。神经样细胞不表达成熟神经细胞标志蛋白MAP2,可能是因诱导时间不够,因为神经母细胞分化为功能性神经细胞需要数月时间。Abouelfetouh等[11]也同样研究发现BMSCs同海马组织碎片共同培养第3天,神经元样细胞所占比例为31.4%±2.79%,轴突样突起平均长56μ,但随着培养时间的延长,神经元样细胞所占比例逐渐降低,而轴突样突起却逐渐增长,第14天时神经元样细胞所占比例为6.8%±0.8%,轴突样突起已在细胞间交织成网很难测量,培养基中添加RA后可促进神经元样细胞数的增加。免疫组织化学检测发现一个有趣的现象,那些突起较长相对较成熟的细胞并不表达NeuN表面抗原,而那些突起较短未成熟的细胞却NeuN表面抗原呈阳性,由BMSCs分化来的形态上类似神经元的细胞中9.6%±1.7%表达NeuN抗原,所有神经元样细胞不表达胶质细胞特征抗原GFAP和Iba1,而且同时观察到这种神经元样细胞分化仅发生在海马组织粹片边缘。这也证实BMSCs具有神经细胞分化潜能,且分化动力受诱导因子及周围微环境(与神经组织的直接接触)双重影响。在Wislet-Gendebien[12]将BMSCs与小脑颗粒细胞共同培养的实验中发现成体骨髓基质细胞不仅能表达神经元特异抗原,还可与γ-氨基丁酸、甘氨酸、谷氨酸等神经递质发生反应产生单程动作电位,说明BMSCs可分化为能产生动作电位具有功能性的神经样细胞,但BMSCs分化为神经样细胞需要两个前提条件:①BMSCs表达nestin蛋白;② BMSCs与神经细胞直接接触后产生的信号整合作用。Tseng等[13]同样也证实,BMSCs在体外经生长因子和化学处理后,可向神经样细胞分化,表达神经元特异标记如神经丝蛋白NF-H(35.7±8.1%)、β-微管蛋白Ⅲ(21.8±6.3%)、tau(90.4±7.5%),还有少量神经样细胞(lt;0.1%)可分泌神经递质γ-氨基丁酸(GABA)。关于BMSCs神经细胞分化潜能的很多研究都证实BMSCs可在体外诱导为神经样细胞,并具有某些神经细胞样功能如分泌神经递质、与神经递质反应产生动作电位等。
3 骨髓基质细胞在神经系统疾病中的研究应用
因为BMSCs具有神经细胞分化潜能,所以已经作为理想的种子细胞应用于神经系统疾病的治疗研究中[14]。缺血性脑血管病是临床常见疾病,具有较高的致死、致残率,若发生不可逆性脑缺血将导致脑组织缺血、缺氧性坏死,造成受损脑组织相关神经功能永久性缺失,给患者的生存造成极大危害,因此如何修复受损脑组织恢复患者的神经功能是治疗缺血性脑血管病的关键。Shen等[15]曾在探索BMSCs移植治疗大脑中动脉闭塞 (middle cerebral artery occlusion MCAO)脑缺血的实验研究中发现细胞移植治疗2周后脑缺血动物模型的神经缺失症状开始得到改善,并且这种治疗效应至少可持续1年,经原位杂交反应也证实移植后的细胞可在宿主脑内存活并部分(22.3%±1.95%)表达星形胶质细胞特异蛋白GFAP、部分(16.8%±2.13%)表达神经元特异蛋白MAP-2,可增加缺血组织周围突触囊泡蛋白表达明显减轻突触缺失。其结果提示BMSCs脑内移植后可分化为神经样细胞,替代修复受损的神经组织。Chen等[16]研究BMSCs脑内移植治疗短暂性大脑中动脉闭塞大鼠模型时也证实,移植后的BMSCs不仅能够存活而且向损伤脑组织周围迁移,并部分细胞表达神经胶质细胞特异标志物,减轻脑白质的损伤,促进脑皮质内β-Ⅲ-tubulin分泌,明显改善MCAO造成的皮质功能缺失症状。BMSCs在移植治疗脑缺血的实验研究中显现出的突出效果使其成为目前研究热点。
帕金森病是老年人中一种常见的慢性、进行性中枢神经系统退行性疾病,其病理特征主要是黑质内多巴胺能神经元退行性变、数目减少,导致纹状体内多巴胺递质水平下降,引起震颤、肌强直和运动迟缓等锥体外系症状。Lu[17]等曾利用腺相关病毒载体将多巴胺生成限速酶-酪氨酸羟化酶(tyrosine hydroxylase,TH)基因导入BMSCs,然后将表达TH基因的BMSCs移植到帕金森病模型鼠的纹状体内后观察发现治疗组阿扑吗啡诱导的不对称旋转症状较对照组明显减轻,6周后免疫组化检测发现TH基因主要在移植点周围表达,高效液相色谱仪及电化学检测也证实TH基因修饰的BMSCs纹状体内移植可明显增加移植点周围多巴胺递质水平。证实BMSCs可作为帕金森病基因治疗的有效载体细胞,为帕金森病的基因治疗提供了前提条件。Zhang等[18]先将BMSCs经体外诱导为神经干细胞,然后利用基因整合技术将TH基因转入其中,并移植入帕金森病大鼠模型脑室及纹状体内,10周后观察发现PD模型的不对称旋转症状明显减轻且活动能力得到显著改善,同对照组比较,纹状体内移植组多巴胺水平提高46.6%,脑室内移植组巴胺水平提高33%,而且观察到移植细胞沿着基底神经纤维发生迁移。这进一步证实了BMSCs作为种子细胞在帕金森病治疗研究中的应用价值。而且还有实验研究证实BMSCs具有多巴胺能神经元分化潜能[19]。所有这些实验研究均为BMSCs作为干细胞移植治疗帕金森病提供了理论依据。
阿尔茨海默病又名老年性痴呆,是引起老年人痴呆的最常见原因,其特征性病理改变主要有老年斑的形成、神经元纤维缠结及神经元缺失等。AD患者由于进行性智能障碍,最终导致完成丧失生活自理能力,给患者及家人造成严重影响,而目前又无有效的治疗手段,所以设法阻止或延缓神经元的缺失,缓解患者的智能衰退是治疗的关键。Wu等[20]将绿色荧光蛋白标记的BMSCs移植入β-淀粉样蛋白所致AD大鼠模型海马内,2周后水镜迷津测验证实BMSCs移植治疗组的认知能力得到明显改善,刚果红着色显示AD模型脑内神经元减少,海马CA1区尤为明显,而BMSCs移植治疗后脑内神经元虽然不能达到正常但还是有所增加,荧光视踪和免疫组织化学检测发现移植细胞在脑内存活并迁移而且可能转化为胆碱乙酰转移酶(choline acetyl transferase,ChAT)阳性神经元。Lee等[21]也研究证实BMSCs脑内移植后,可促进脑内小胶质细胞吞噬活性,减少β-淀粉样蛋白在急性诱导的AD小鼠模型脑内的存积,减轻β-淀粉样蛋白对神经元的损伤,提示了BMSCs治疗AD的可能性。
4 骨髓基质细胞治疗神经系统疾病的可能机制
对于骨髓基质细胞的神经修复作用机制目前主要倾向于认为可能有两方面原因:①分化为神经细胞替代修复缺损的神经组织,体内外实验研究均证实BMSCs可向神经细胞分化[11,15],并可能分泌神经递质[19]、与神经递质发生反应产生动作电位[12]。Tropel等[22]也研究发现BMSCs不仅可分化为神经样细胞,而且分化后的细胞具有神经元样的功能,可与谷氨酸、藜芦定碱反应增加胞质内钙离子浓度,而且这种效应可分别被N-甲基-D-门冬氨酸受体阻断剂、河豚毒素所阻断,同样发现BMSCs源性神经样细胞可与多巴胺反应增加胞质内钙离子浓度,所有这些均证实BMSCs源性神经样细胞表达功能性神经递质受体,可作为干细胞用于神经系统的治疗。Cogle等[23]在一项BMSCs脑内移植后的临床回顾性研究中发现,BMSCs可在脑内长期存活,并可转化为神经元、星形胶质细胞和小胶质细胞,尽管发生转化的细胞数量很少,但还是说明BMSCs有可能通过神经组织转化来替代修复缺失的神经细胞,恢复其缺损的神经功能。②BMSCs可能通过自分泌或刺激周围神经组织分泌各种神经营养因子,减轻神经细胞的损伤,促进神经细胞的存活和再生。Garcia等[24]体外培养BMSCs至第6代时提取总的RNA后加入特异引物,RT-PCR反应证实BMSCs表达NGF和GDNF。 Yaghoobi等[25]也研究证实BMSCs在分化为神经样细胞前后均表达NGF和BDNF。Mahmood等[26]研究发现BMSCs通过静脉移植后可向损伤脑组织处迁移,并明显改善其神经功能,经酶联免疫吸附测定(ELISA)证实BMSCs可增加脑组织内NGF和BDNF的表达,因而证明BMSCs有可能是通过增加脑组织内相关神经生长因子如NGF、BDNF等来保护受损的神经组织、降低神经功能缺损。
5 尚存的问题与展望
由于可取材于自体,来源方便,且增殖分化能力强,可避免细胞数量不足、免疫排斥、伦理道德的限制等自身优势,同时又具有跨胚层分化潜能,可向神经细胞分化等特性,所以BMSCs已被作为理想的种子细胞应用于神经系统疾病的治疗研究中。体内外大量实验研究也证实BMSCs具有神经元、神经胶质细胞分化潜能,表现出某些神经细胞功能特征,如分泌神经递质[19]、与神经递质反应产生动作电位[12]等,且在神经系统疾病的治疗研究中取得了理想的疗效,对缺损的神经功能具有治疗修复作用。但同时BMSCs应用于神经系统疾病的研究中还存在诸多问题尚未解决。首先,目前还未发现BMSCs的特异性标志,对BMSCs的鉴别主要依靠其形态、功能特征及培养中出现的分化表型来推断,得到的细胞神经分化潜能各异,因此如何纯化鉴定得到神经细胞分化潜能最强的单一的BMSCs还是一个有待解决的问题。其次,BMSCs移植后改善神经功能的确切机制还不清楚,并且关于BMSCs向神经细胞分化的研究也仅仅只是依靠其形态特征及少数神经细胞特异标志来鉴别,而BMSCs能否真正分化为神经细胞、发挥神经细胞功能、与宿主的神经组织发生有效整合、修复缺失的神经功能等还未得到充分证明[27]。再者,BMSCs的安全性问题还有待进一步证实,BMSCs体内移植后是否会发生基因突变,引起致瘤性[28]目前还无可靠证据。最后,利用BMSCs进行基因治疗时也面临着转染效率、转基因细胞的靶向性及基因表达的持续性和可控性等问题。尽管关于BMSCs治疗神经系统疾病的研究还存在许多未知因素,但BMSCs在神经系统疾病的治疗研究中显示出来的客观效果及随着研究的不断深入,BMSCs有望成为神经疾病患者的福音。
[1]Gensburger C,Labourdette G,Sensenbrenner M. Brain basic fibroblast growth factor stimulates the proliferation of rat neuronal precursor cells in vitro[J]. FEBS Lett, 1987, 217(1): 1-5.
[2]Friedenstein A J,Gorskaja J F,Kulagina N N.Fibroblast precursors in normal and irradiated mouse hematopoietic organs [J].Exp Hematol,1976,4(5):267-274.
[3]Prockop D J.Marrow stromal cells as stem cells for nonhematopoietic tissues[J].Science,1997,276(5309):71-74.
[4]Sergeeva N S,Sviridova I K, Kirsanova V A,et al.Study of growth parameters and potentialities of differentiation of multipotent mesenchymal stromal cells from rat bone marrow in vitro[J]. Bull Exp Biol Med, 2006, 141(4):530-535.
[5]Pittenger M F,Mackay A M,Beck S C,et al.Multilineage potential adult human mesenchymal stem cells[J].Science, 1999,284(5411):143-147.
[6 ]Wexler S A,Donaldson C,Denning-Kendall P, et al . Adult bone marrow is a rich source of human mesenchymal′stem′cells but umbilical cord and mobilized adult blood are not [J ]. Br J Haematol,2003, 121 (2) :368-374.
[7]Friedenstein A J,Latzinik N V,Gorskaya Y F,et al .Bone marrow stromal colony formation requires stimulation by hematopoietic cells[J].Bone Miner,1992,18(3):199-213.
[8]Yoo J U,Barthel T S,Nishi K,et al.The chondrogenic potential of human bone-marrow-derived mesenchymal progenitor cells[J].J Bone Joint Surg,1998,80(12):1745-l 757.
[9]Quirici N,Soligo D,Bossolasco P,et al.Isolation of bone marrow mesenchymal stem cells by anti-nerve growth factor receptor antibodies[J].Exp Hematol,2002,30(7):783-791.
[10]Sanchez-Ramos J, Song S, Cardozo-Pelaez F, et al. Adult Bone Marrow Stromal Cells Differentiate into Neural Cells in Vitro[J]. Exp Neurol, 2000,164(2):247-256.
[11]Abouelfetouh A, Kondoh T, Ehara K,et al . Morphological differentiation of bone marrow stromal cells into neuron-like cells after co-culture with hippocampal slice[J]. Brain Res, 2004, 1029(1):114-119.
[12]Wislet-Gendebien S,Hans G,Leprince P, et al .Plasticity of cultured mesenchymal stem cells: switch from nestin-ostive to excitable neuron-1ike phenotype[J].Stem Cells, 2005, 23 (3) :392-402.
[13]Tseng PY, Chen C J, Sheu C C, et al. Spontaneous differentiation of adult rat marrow stromal stem cells in a long-term culture. J. Vet[J]. Med. Sci, 2007, 69(2):95-102.
[14]Dezawa M,Hoshino M,Ide C.Treatment of neurodegenerative diseases using adult bone marrow stromal cell-derived neurons[J].Expert Opin BioI Ther, 2005,5(4):427-435.
[15]Shen L H, Li J, Chen J,et al. One-year follow-up after bone marrow stromal cell treatment in middle-aged female rats with stroke[J]. Stroke,2007, 38:2150-2156.
[16]Chen J R, Cheng G Y, Sheu C C, et al. Transplanted bone marrow stromal cells migrate, differentiate and improve motor function in rats with experimentally induced cerebral stroke[J]. J Anat, 2008, 213:249-258.
[17]Lu L, Zhao C, Liu Y, et al. Therapeutic benefit of TH- engineered mesenchymal stem cells for Parkinson’s disease [J]. Brain Res Brain Res Protoc, 2005, 15(1): 46- 51.
[18]Zhang S Z, Zou Z H, Jiang X D, et al. The therapeutic effects of tyrosine hydroxylase gene transfected hematopoetic stem cells in a rat model of Parkinso’s disease[J]. Cell Mol Neurobiol, 2008, 28:529-543.
[19]Trzaska K A, Kuzhikandathil E V, Rameshwar P. Specification of a dopaminergic phenotype from adult human mesenchymal stem cells[J]. Stem Cells, 2007, 25(11):2797-2808.
[20]Wu QY, Li J, Feng Z T, et al. Bone marrow stromal cells of transgenic mice can improve the cognitive ability of an Alzheimer's disease rat model. Neurosci Lett,2007,417(3) : 281-285.
[21]Lee J K, Jin H K, Bae J S. Bone marrow-derived mesenchymal stem cells reduce brain amyloid-β deposition and accelerate the activation of microglia in an acutely induced Alzheimer’s disease mouse model. Neurosci Lett, 2009, 450(2): 136-141.
[22]Tropel P, Platet N, Platel J C, et al.Functional neuronal differentiation of bone marrow-derived mesenchymal stem cells[J].Stem Cells, 2006, 24(12):2868-2876.
[23]Cogle C R, Yachnis A T, Laywell E D, et al. Bone marrow transdifferentiation in brain after transplantation: A retrospective study[J]. Lancet, 2004,363(9419): 1432-1437.
[24]Garcia R, Aguiar J, Alberti E, et al. Bone marrow stromal cells produce nerve growth factor and glial cell line-derived neurotrophic factors[J]. Biochem Biophys Res Commun, 2004, 316(3): 753-754.
[25]Yaghoobi M M, Mowla S J. Differential gene expression pattern of neurotrophins and their receptors during neuronal differentiation of rat bone marrow stromal cells[J]. Neurosci Lett, 2006, 397(1-2): 149-154.
[26]Mahmood A, Lu D, Chopp M. Intravenous administration of marrow stromal cells(MSCs )increases the expression of growth factors in rat brain after traumatic brain injury[J]. Neurotrauma, 2004, 21(1): 33-39.
[27]Raedt R, Pinxteren J, Van Dycke A. et al. Differentiation assays of bone marrow-derived Multipotent Adult Progenitor Cell (MAPC)-like cells towards neural cells cannot depend on morphology and a limited set of neural markers[J]. Exp Neurol, 2007, 203(2): 542-554.
[28]Burns J S, Abdallah B M, Guldberg P, et al. Tumorigenic heterogeneity in cancer stem cells evolved from long-term cultures of telomere[J]. Cancer Res, 2005,65: 3126-3135.
[编辑] 何 勇
10.3969/j.issn.1673-1409(R).2012.02.034
R741
A
1673-1409(2012)02-R068-05
2011-11-29
谢三平(1980-),男,湖北监利人,医师,硕士,主要从事神经内科临床工作。
猜你喜欢
中国临床医学影像杂志(2021年10期)2021-11-22
中成药(2021年5期)2021-07-21
昆明医科大学学报(2021年2期)2021-03-29
国际放射医学核医学杂志(2020年3期)2020-07-27
文苑(2018年18期)2018-11-08
生命科学研究(2018年1期)2018-05-29
现代检验医学杂志(2016年2期)2016-11-14
IT时代周刊(2015年9期)2015-11-11
癌变·畸变·突变(2014年1期)2014-03-01
铁路通信信号工程技术(2014年5期)2014-02-28
